
Abstract
The coffee berry borer (CBB, Hypothenemus hampei) is the world's most important coffee pest. In Colombia an Integrated Pest Management Programme is being developed for this pest, whose principal components are cultural control, biocontrol and when absolutely necessary, chemical control. One of the most promising natural enemies of the CBB is the parasitoid C. stephanoderis. Apart from its parasitic action on the immature stages, it has also been observed to act as a predator. As there are no experimental studies on this aspect, the aim of this study was to quantify the predator action of this wasp under field conditions.
The experiments were carried out in the Rafael Escobar experimental substation at Supía, Caldas, 1320 m above sea level, 21,9 °C mean annual temperature, 2020 mm rainfall and 75% RH, in a plot of 3 year old Coffea arabica var. Colombia. Previously uninfested berries were infested artificially with CBB in entomological sleeve cages around the branches. Wasps were released for treatments of 5,10 and 15 days after infestation either (a) inside the sleeves, (b) free field release or (c) control. Four destructive samples were taken at 3, 10, 20 and 30 days after wasp release. For the controls, there were means of 10, 13 and 15 CBB stages per berry at 5, 10 and 15 days after infestation. For the sleeve cages there were 3, 5 and 6 stages and for the open field treatment, 4, 7 and 8 stages per berry. These latter two were significantly lower than the controls. The wasps reduced the CBB population by 65% in the cages and 49% in open field conditions, demonstrating their effectiveness as predators. The extra mortality caused by predation may thus significantly enhance their effectiveness as control agents of CBB.
Introducción
La broca del café, Hypothenemus hampei (Ferrari), es sin duda la plaga de mayor importancia económica en el cultivo del café, a nivel mundial, ya que afecta directamente al fruto (Cenicafé 1990). Para su control en Colombia, el Centro Nacional de Investigaciones del Café - Cenicafé- está implementando un programa de manejo integrado de la broca (MIB), con énfasis en control biológico, mediante el uso de parasitoides de origen africano y algunos hongos entomopatógenos (Bustillo 1990).
Los parasitoides Cephalonomia stephanoderis Betrem y Prorops nasuta Waterston, ambos (Hymenoptera: Bethylidae), que actúan como ectoparásitos de estados inmaduros de H. hampei y Phymastichus coffea (La Salle) (Hymenoptera: Eulophidae), que actúa como endoparásito de adultos de la broca, son los enemigos naturales más opcionados para el control biológico de la plaga en Colombia (Orozco y Aristizábal 1996). De los tres parasitoides introducidos, C. stephanoderis es el que mejores resultados ha presentado en cuanto al desarrollo de una metodología de cría masiva, lográndose una producción de más de 384 millones de parasitoides, lo suficiente para llevar un programa de introducción de C. stephanoderis en las diferentes zonas cafeteras del país con presencia de la plaga (Bustillo et al. 1996).
Los estudios en Colombia sobre adaptación, establecimiento, técnicas de liberación, dispersión, efecto controlador, compatibilidad con otras medidas de control y capacidad de búsqueda de frutos infestados con H. hampei, permiten confirmar la eficacia del parasitoide C. stephanoderis y su importancia como agente de control biológico de las poblaciones de la broca en los agroecosistemas cafeteros del país (Benavides et al. 1994; Aristizábal 1995; Bustillo et al. 1996, Aristizábal et al. 1997).
Benavides et al. (1994) evaluaron las primeras liberaciones masivas de C. stephanoderis en Colombia, en plantaciones de café con altos niveles de infestación de H. hampei; encontraron parasitismos entre 22 y 65% y comprobaron su establecimiento después de 2 años de haber sido liberada en el campo. Liberaciones de C. stephanoderis demostraron que el parasitoide logró establecerse en cafetales con bajos niveles de infestación de H. hampei (inferiores al 5%), registrando parasitismos entre 3.64 y 18.20%; se observó una reducción significativa de la población plaga y se demostró que el parasitoide puede ser utilizado dentro del programa de manejo integrado de la broca (Aristizábal, 1995; Aristizábal et al. 1997). Sin embargo, los actuales costos de la producción de los parasitoides pueden ser un obstáculo para que los caficultores puedan adquirirlos en grandes cantidades (Bustillo et al. 1996).
Además del efecto parasítico que presenta C. stephanoderis sobre larvas, prepupas y pupas de H. hampei, algunos autores han observado su efecto depredador sobre todos los estados biológicos de H. hampei, (Ticheler 1963; Abraham et al. 1990; Murphy y Moore 1990; Barrera et al. 1991; Benavides et al. 1994). Sin embargo estas observaciones fueron realizadas como complemento de investigaciones de laboratorio y campo que no necesariamente tenían el objetivo de evaluar el efecto depredador del parasitoide. La presente investigación tuvo como objetivo principal determinar el efecto depredador de C. stephanoderis sobre todos los estados biológicos de H. hampei en condiciones de campo, establecer sí la población de la plaga se puede reducir por la acción depredadora del parasitoide y conocer la preferencia de estados inmaduros de H. hampei sobre los cuales se pueda alimentar el betílido.
Materiales y Métodos
El experimento se realizó en la Subestación Experimental "Rafael Escobar" de Cenicafé localizada en el municipio de Supía, Caldas, ubicada a 1320 msnm, 21,9°C de temperatura media anual, 2020 mm de precipitación anual acumulada y 75% de humedad relativa, en un lote de café variedad Colombia de tercer año de cosecha, con 0,3 hectáreas de extensión aproximadamente.
Infestación artificial de H. hampei
Se realizaron infestaciones artificiales de H. hampei sobre frutos sanos de café mayores a 20 semanas de desarrollo fisiológico, mediante la utilización de mangas entomológicas. Se infestaron artificialmente 32 ramas de café por tratamiento, para un total de 288 ramas infestadas, utilizando una relación de 4:1 (adultos de H. hampei por fruto de café sano). Posteriormente, se realizó una liberación del parasitoide C. stephanoderis en relación 5:1 (avispas por fruto infestado).
Se realizaron tres tiempos de infestación (5, 10 y 15 días) logrando suministrar a los parasitoides diferentes estados biológicos de H. hampei en forma simultánea, de tal manera que el parasitoide pudiera escoger el sustrato más conveniente para alimentarse.
Descripción de tratamientos
El experimento se realizó bajo un diseño completamente aleatorio con arreglo factorial de 3 x 3, correspondiente a tres tiempos de infestación con H. hampei: 5, 10 y 15 días y la liberación del parasitoide en tres condiciones diferentes: Dentro de las mangas entomológicas, sin mangas y los controles, conformando nueve tratamientos (Tabla 1).
En los tratamientos control 1, 2 y 3 y en los tratamientos en los cuales se liberaron los parasitoides dentro de las mangas entomológica 4, 5 y 6 se dejaron las mangas puestas en las ramas durante todo el tiempo de evaluación, 30 días. En los tratamientos 7, 8 y 9 se retiraron las mangas antes de la liberación de los parasitoides para evaluar el efecto depredador del parasitoide bajo condiciones de campo abierto..
Se realizaron cuatro muestreos de frutos infestados a los 3, 10, 20 y 30 días después de la liberación de los parasitoides, para hacer un seguimiento del comportamiento del parasitoide y del insecto plaga a través del tiempo. Se colectaron 200 frutos por tratamiento, distribuidos en 8 repeticiones; en total se disecaron 7200 frutos durante el experimento.
En las disecaciones de los frutos infestados se cuantificó el número de estados biológicos de H. hampei (huevos, larvas, pupas, adultos vivos y adultos muertos), se determinó el porcentaje de frutos colonizados por C. stephanoderis, el número de estados biológicos de C. stephanoderis y el porcentaje de frutos infestados con ausencias de adultos de H. hampei. Se calculó el análisis de varianza para las variables evaluadas y se realizó la comparación de medias utilizando la prueba de contrastes ortogonales, comparando los tratamientos testigo con los tratamientos con avispa.
Resultados y Discusión
Porcentaje de frutos colonizados por C. stephanoderis
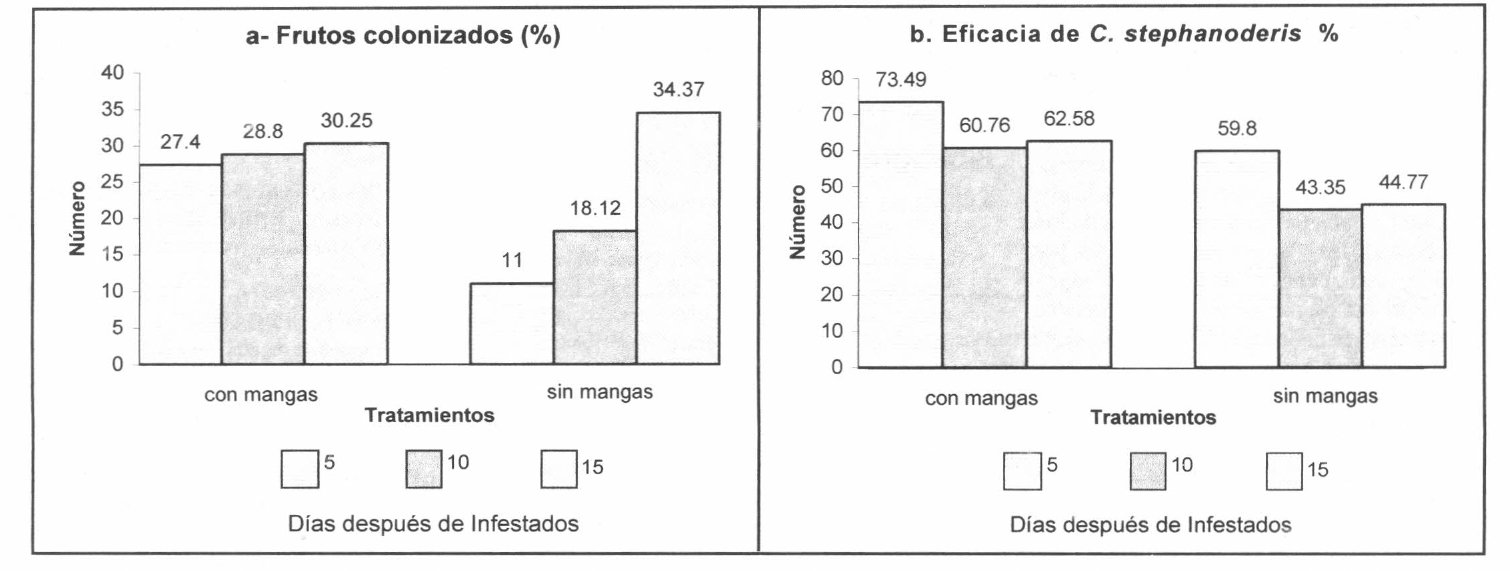
El parasitoide C. stephanoderis prefiere colonizar en mayor proporción los frutos de mayor edad de infestación: de 15, 10 y 5 días, es decir, los frutos que, en este orden de edad, presentaron mayor cantidad de estados inmaduros de H. hampei; sin embargo, para todos los tiempos de infestación se evidenció la presencia del parasitoide. El porcentaje de frutos colonizados por C. stephanoderis presentó diferencias altamente significativas (P = 0.0001) entre los tratamientos (Fig. 1 a).
Bajo condiciones de campo se ha observado que C. stephanoderis coloniza y parasita frutos de café infestados de todos los estados de desarrollo fisiológico (verdes consistentes, pintones, maduros, sobre maduros y secos), siempre y cuando existan estados inmaduros de la broca dentro de ellos; sin embargo, prefiere aquellos frutos que presentan mayor cantidad de estados inmaduros de la plaga, observándose parasitismos más altos en los frutos maduros, sobre maduros y secos con respecto a los demás (Aristizábal et al. 1996), ya que estos frutos, por tener mayor edad fisiológica y mayor peso seco, presentan condiciones más favorables para el desarrollo de las poblaciones de la broca (Ruiz 1996). Esto sugiere la posibilidad de que exista una hormona producida por las larvas de H. hampei que atrae al parasitoide.
Descripción de tratamientos del experimento sobre el efecto depredador de C. stephanoderis sobre H. hampei
Promedio de huevos de H. hampei después de la liberación de C. stephanoderis
DDL = Días después de la liberación de C. stephanoderis. T= testigos, CM = con mangas, SM = sin mangas
Estados inmaduros de H. hampei
Con respecto al número de huevos de H. hampei encontrados en los frutos durante los cuatro muestreos: 3, 10, 20 y 30 días después de la liberación de las avispas, los diferentes análisis de varianza presentaron diferencias altamente significativas entre los tratamientos (P=0.0001) (Tabla 2). Según la prueba de contrastes ortogonales, para los tratamientos en los cuales se liberaron las avispas, se presentó una reducción significativa en el número de huevos, con respecto a los testigos, especialmente en las evaluaciones de los 3 y 20 días después de la liberación de los parasitoides. Esta reducción evidencia el efecto depredador que presenta el C. stephanoderis sobre huevos de H. hampei.
Los tratamientos con liberación de avispas en mangas y en campo abierto fueron significativamente menores en el número promedio de huevos de H. hampei respecto a los tratamientos control. Se observó reducción en el número de huevos de 57% ± 8,4 y 38,3% ± 10,7 en promedio, para los tratamientos con liberación en mangas y en campo abierto respectivamente, como consecuencia de la acción depredadora del parasitoide.
Se presentó mayor cantidad de larvas de H. hampei, en la medida en que los frutos infestados tenían mayor edad de infestación; esta tendencia fue observada en los tratamientos. Sin embargo, los análisis de varianza presentaron diferencias altamente significativas (P = 0.0001). Según la prueba de contrastes ortogonales, se encontró menor cantidad de larvas en los tratamientos con avispas frente a los tratamientos testigo (Tabla 3). La reducción en el número de larvas de H. hampei fue de 68,7% ± 13,2 y 52,8% ± 10,5 en promedio, para los tratamientos con liberación en mangas y en campo abierto, respectivamente.
Con respecto a las pupas de H. hampei los análisis de varianza presentaron diferencias significativas entre los tratamientos (P = 0,0001). En los tratamientos en los cuales se liberaron las avispas se observaron valores inferiores a 0,65 pupas por fruto, resultando ser significativamente menores a los testigos. Es factible que en el consumo de huevos y larvas de primer ínstar por parte de C. stephanoderis evitó el desarrollo de pupas y por tanto la cantidad fue muy baja con respecto a los tratamientos testigo y no como consecuencia del consumo de ellas por parte del parasitoide. Por lo anterior se deduce que el parasitoide prefiere consumir principalmente larvas y huevos.
Estos resultados demuestran el efecto depredador que presenta C. stephanoderis sobre los estados inmaduros de H. hampei, lo anterior concuerda con Bustillo et al. (1996), quienes afirman no haber encontrado huevos y larvas de primer ínstar en frutos parasitados. También con Barrera et al. (1991), quienes evaluaron el impacto del parasitoide sobre las poblaciones de broca, observando no sólo el efecto parasítico, sino también el efecto depredador de la hembra del betílido, la cual consume huevos y larvas de la broca durante su permanencia en el interior del fruto.
Adultos de H. hampei vivos, muertos y ausentes
En los tratamientos testigo se encontraron 1,3 ± 0,2; 1,15 ± 0,1 y 1,1 ± 0,1 adultos de H. hampei vivos en promedio, para los frutos de 5, 10 y 15 días de infestados respectivamente; en los tratamientos con parasitoides se observaron 0,44 ± 0,2 adultos vivos por fruto. Para el número de adultos vivos se presentaron diferencias altamente significativas entre los tratamientos (P = 0,0001). Según la prueba de contrastes ortogonales fue significativamente menor el número de adultos vivos de los tratamientos con avispas respecto a los testigos, observándose una reducción de 66,6% ± 5,5 y de 58,5% ± 6,2 para los tratamientos con liberación en mangas y en campo abierto, respectivamente.
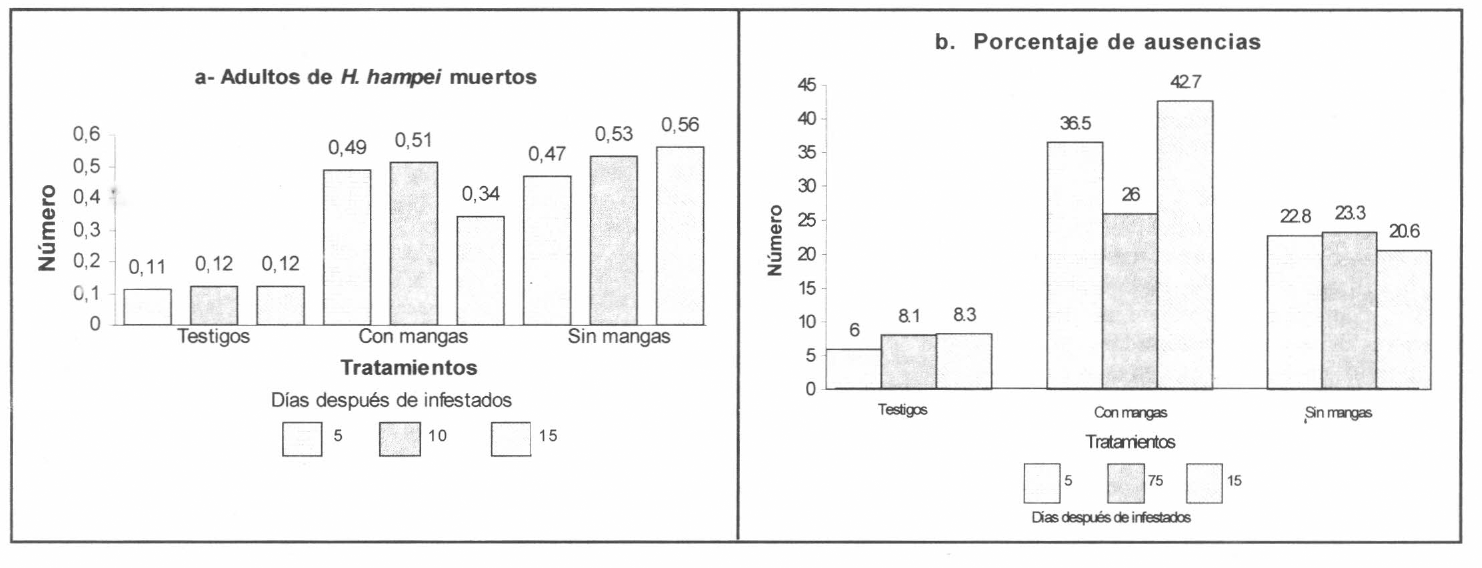
En relación con el número de adultos de H. hampei muertos, se presentaron diferencias altamente significativas (P=0.0001). Según la prueba de contrastes ortogonales fue mayor la cantidad de adultos muertos en los tratamientos con avispas respecto a los testigos. Se observaron en promedio 0,12 ±0,002 adultos de H. hampei muertos por fruto infestado en los tratamientos testigo, mientras que en los tratamientos con liberación de avispas en mangas y en campo abierto se obtuvieron promedios de 0,44 ± 0,07 y 0,52 ± 0,03 adultos muertos respectivamente (Fig. 2a).
Promedio de larvas de H. hampei después de la liberación de C. stephanoderis
DDL = Días después de la liberación de C. stephanoderis. T= testigos, CM = con mangas, SM = sin mangas
Número total de estados inmaduros de H. hampei después de la liberación de C. stephanoderis durante los cuatros muestreos
DDL = Días después de la liberación de C. stephanoderis. T= testigos, CM = con mangas, SM = sin mangas
Estos resultados indican que frutos infestados con H. hampei presentaron en promedio el 10% ± 0,19 de mortalidad natural en adultos, mientras que con la presencia del parasitoide, las mortalidades fueron significativamente mayores. Se obtuvo en promedio 37,8% ±6,4 y 44% ± 3,1 de mortalidad para la liberación en mangas y en campo abierto, respectivamente. Se observaron hembras adultas de H. hampei sin cabeza, con mutilaciones en las patas, mandíbulas y antenas, como consecuencia del ataque directo del parasitoide.
Según la prueba de contrastes ortogonales el porcentaje de adultos de H. hampei ausentes de los frutos infestados, en los tratamientos testigo, fue de 7,5% ± 1 en promedio; siendo significativamente menor a los tratamientos con parasitoides. Se presentaron promedios de 35% ± 6,9 y 22% ± 1,1 de ausencias para la liberación en mangas y en campo abierto, respectivamente (Fig. 2 b). Estos incrementos en el porcentaje de ausencias evidencia un ataque del parasitoide sobre los adultos de H. hampei, los cuales son eliminados y posiblemente sacados de los frutos.
Es posible que el parasitoide con el fin de buscar frutos infestados con un buen número de estados inmaduros susceptibles de ser parasitados, haya penetrado a varios frutos eliminando los adultos de H. hampei, alimentándose de éstos y de algunos estados inmaduros como huevos y larvas de primer ínstar, hasta encontrar un fruto en condiciones óptimas para parasitarlo, explicando así mayor cantidad de adultos muertos y mayores porcentajes de ausencias en los tratamientos con avispas frente a los testigos.
Los resultados obtenidos concuerdan con observaciones realizadas por Benavides et al. (1994), quienes observaron en frutos colonizados por la avispa, una mortalidad de la broca, en todos sus estados, del 94,84%. Ticheler (1963) encontró en frutos parasitados por la avispa un 89,2% de mortalidad de la broca y observó su capacidad depredadora. Sponagel (1994) afirma que en una cereza de café infestada con H. hampei y colonizada por C. stephanoderis, el 46,7% de las larvas estaban parasitadas (acción como parasitoide) y el 64.4% de los adultos de H. hampei estaban muertos (acción como depredador), confirmando tal efecto. Barrera et al. (1991) observaron en frutos parasitados, hembras adultas de la broca que aparecen sin cabeza o sin patas, registrando mortalidades en los adultos de la broca entre 80 y 100%. Koch (1973) resalta la condición parasítica y depredadora de C. stephanoderis, afirmando que la hembra adulta de la avispa se alimenta de la hemolinfa de brocas adultas, rompiendo con sus mandíbulas la membrana intersegmental de los tergitos proto - y mesatoráxicos.
Número total de estados inmaduros de H. hampei
Para el número total de estados inmaduros de H. hampei por fruto, se presentaron diferencias altamente significativas entre los tratamientos (P=0.0001). Según la prueba de contrastes ortogonales se observó menor número de estados inmaduros en los tratamientos con parasitoides respecto a los tratamientos testigo.
En los testigos se observaron promedios de 10,2 ± 2,5; 12,8 ± 1,1 y 14,9 ± 2,7 estados de H. hampei por fruto, para los 5, 10 y 15 días de infestados respectivamente; mientras que, los tratamientos con liberación en mangas presentaron 2,7 ± 1,6; 5,1 ± 0,9 y 5,6 ± 2,2 estados inmaduros, observándose reducciones significativas en el número de estados de 73,5; 60,7 y 62,5%, respectivamente; y en la liberación a campo abierto se encontraron 4,1 ± 1; 7,3 ± 2,1; y 8,2 ± 1,8 estados de H. hampei, observándose reducciones significativas de 59,8; 43,3 y 44,7%, respectivamente (Fig.1b). Estos resultados indican que el parasitoide C. stephanoderis presenta un efecto depredador sobre todos los estados inmaduras de H. hampei, al reducir en promedio las poblaciones de la plaga en 49,3% ± 7,4 y en 65,6% ± 5,6 para campo abierto y en mangas, respectivamente (Tabla 4).
Gauld y Bolton (1988), al estudiar el comportamiento y las formas de vida de los parasitoides hymenopteros, afirman que los hábitos de depredación son una especialización de algunos ectoparásitos cuyas hembras adultas, para suplirse de proteínas, presentan un comportamiento conocido como "host feeding" el cual esta exactamente relacionado para la producción de huevos, aunque si bien, su importancia muchas veces no es apreciada. La mayoría de los parasitoides que practican el "host - feeding", se alimentan de los estados del hospedero que no van a ser usados para la parasitación (Gauld y Bolton 1988). Con respecto a C. stephanoderis varios autores han observado este comportamiento sin referirse a él directamente, al afirmar que el parasitoide se alimenta de huevos, larvas de primer ínstar y adultos de H. hampei (Ticheler 1963; Klein et al. 1988; Abraham et al. 1990; Barrera et al. 1991; Bustillo et al. 1996), estados que el parasitoide no puede parasitar.
Stehr (1990) afirma que la depredación es común entre los insectos y los casos de mayor éxito en el control biológico han tenido que relacionarse con la depredación; también comenta que los principios básicos de la depredación (relación presa depredador) se aplican a los parasitoides, considerándolos como un tipo especializado de depredadores, ya que los parasitoides adultos atacan a la presa de diferente forma, observándose casos en donde el parasitoide mata al hospedero para alimentarse (depredación). Este comportamiento puede ser aplicado directamente al parasitoide C. stephanoderis, ya que las observaciones realizadas por diferentes autores y los resultados encontrados en esta investigación así lo demuestran.
Para la población del hospedero, el "host - feeding" puede ser un factor adicional de mortalidad, con el cual el parasitoide puede causar una mortalidad más alta en el hospedero que la misma parasitación (Gauld y Bolton 1988). Barrera et al. (1990) afirman que la eficacia inmediata de C. stephanoderis en el control de las poblaciones H. hampei en el interior de los frutos, se refleja, no sólo en el efecto del parasitismo, sino también en el efecto de depredación de la avispa sobre todos los estados biológicos de H. hampei. Sponagel (1994) afirma que C. stephanoderis es más efectivo en su acción depredadora que como parasitoide. La reducción de 49% de los estados biológicos de H. hampei encontrada en esta investigación demuestra la eficacia de C. stephanoderis como depredador, corroborando las afirmaciones anteriores, por lo tanto, para el control de H. hampei la acción depredadora parece ser más importante que la acción parasítica.
De otra parte, liberaciones de C. stephanoderis en un cafetal con bajos niveles de infestación de H. hampei (inferior al 5%) redujeron significativamente las poblaciones de la plaga. Se observó mayor cantidad de adultos de H. hampei muertos, menor cantidad de adultos vivos y menor cantidad de estados inmaduros de la plaga con respecto a un lote testigo (Aristizábal 1995; Aristizábal et al. 1997), lo cual obedece al efecto parasítico y depredador que presenta el parasitoide sobre las poblaciones de H. hampei. Se comprobó técnicamente la viabilidad de utilizar a C. stephanoderis dentro del programa de manejo integrado de la broca (MIB); sin embargo, el costo actual del parasitoide sería un limitante para que el caficultor pueda adoptarlo (Bustillo et al. 1996).
Al comprobarse el efecto depredador de C. stephanoderis sobre los estados biológicos de H. hampei se podría plantear, como estrategia, la utilización del parasitoide dentro del MIB en cafetales con niveles de infestación inferiores al 5% y con recolecciones oportunas de frutos maduros, ya que, en estas condiciones, los cafetales presentan escasos frutos infestados, con pocos estados inmaduros de H. hampei; de esta forma los parasitoides tendrían mayor eficacia en su acción depredadora que parasítica y se necesitaría menor cantidad de parasitoides a liberar para regular las poblaciones de la plaga.
Conclusiones
Se comprobó que el parasitoide C. stephanoderis presenta un efecto depredador sobre todos los estados biológicos de H. hampei (huevos, larvas, pupas y adultos) al reducir, bajo condiciones de campo, la población de la plaga. Por lo tanto, parece ser más eficaz C. stephanoderis en su acción depredadora que en su acción parasítica, de ahí su importancia en control biológico dentro del programa de manejo integrado de la broca.
El parasitoide C. stephanoderis prefiere colonizar frutos infestados que presenten mayor número de estados inmaduros de H. hampei, alimentándose especialmente de huevos y larvas de la plaga.
Se evidenció un ataque directo del parasitoide C. stephanoderis sobre los adultos de H. hampei, al observar mayor mortalidad y mayor porcentajes de ausencias de adultos de H. hampei en los tratamientos con avispas respecto a los testigos.
NOTA: Se presentan los siguientes anexos correspondientes a los datos de las figuras presentadas en el texto.
Footnotes
Anexo 1
Datos correspondientes a las figuras 1 a y 1 b.
Anexo 2
Datos correspondientes a las figuras 2 a y 2 b.