
Abstract
Understanding the immune responses of insects is necessary because they are economically important as agricultural pests and disease vectors. Although much information about insect immunity is available from research entomologists, poor knowledge about it is common among most entomologists students and workers. The aim of this review was to collect and present information obtained from the published works concerning this topic and to generate interest for understanding the immune responses and its practical implications for control of pests and parasites transmited by insects.
Introducción
Al igual que otros animales, los insectos poseen un sistema inmune que les permite diferenciar entre lo propio y lo extraño y generar numerosas respuestas ante la presencia de agentes invasores en su organismo.
Los mecanismos de respuesta inmune gracias a los cuales los insectos se defienden de patógenos y parásitos han despertado el interés de numerosos investigadores en los últimos años, ya que su comprensión permite entender, al menos en parte, aspectos como las complejas relaciones que existen entre los insectos y los parásitos que ellos transmiten o cómo estos interactúan con los microorganismos utilizados como agentes de control microbiológico.
Adicionalmente, los insectos constituyen modelos de estudio relativamente simples que podrían facilitar la comprensión de procesos complejos de respuesta inmune en otros organismos, incluyendo el hombre.
Aunque la respuesta inmune de los insectos carece de la especificidad antígeno-anticuerpo presente en otros animales y en el hombre, ellos poseen mecanismos sorprendentemente efectivos cuya diversidad guarda relación con el inmenso tamaño y variabilidad de la clase Insecta.
En la presente revisión se ha hecho énfasis en los mecanismos mejor estudiados y entendidos hasta ahora, incluyendo aspectos que probablemente constituirán objetivos de investigación en el futuro.
Los conceptos incluidos provienen de trabajos originales publicados como resultado de investigaciones y de revisiones previamente realizadas por algunos autores, en las cuales se han sumarizado y puntualizado los aspectos relevantes sobre la inmunología de insectos.
Respuesta inmune en insectos
La presencia de parásitos y/o patógenos en el hemocele puede originar profundos cambios en la fisiología y comportamiento de un insecto infectado; los insectos pueden dejar de alimentarse y su desarrollo puede ser retardado o alterado completamente; la respuesta inmune aparece como un sofisticado sistema que ha evolucionado para combatir estos microorganismos patógenos invasores y sin el cual probablemente los insectos morirían fácilmente a causa de infecciones bacteriales, virales o fúngicas. La habilidad de los insectos para defenderse de los agresores presentes en el medio y asegurar su sobrevivencia fue reconocida muchos años atrás gracias a numerosas investigaciones. En la actualidad se conoce que el sistema inmune permite a los insectos identificar y destruír los microorganismos invasores a través de dos tipos de respuesta:
Los mecanismos específicos originados en cada una de estas respuestas son determinados por aspectos como la naturaleza (biótica o abiótica) y el tamaño de la partícula o microorganismo invasor; estos mecanismos pueden incluír fagocitosis o ingestión de partículas pequeñas, encapsulación y formación de nódulos en el caso de partículas de mayor tamaño y producción de sustancias - generalmente proteínas - con funciones antimicrobiales específicas (Lackie 1988).
Las respuestas como fagocitosis, encapsulación y formación de nódulos, en las cuales participan directamente los hemocitos, o células sanguíneas de los insectos, hacen parte de la respuesta inmune de tipo celular y, aquellas que involucran la producción de sus tancias específicas con propiedades antimicrobiales, conforman la respuesta de tipo humoral (Paskewitz y Christensen 1996).
La respuesta inmune de tipo celular tiene lugar inmediatamente después del contacto con los agentes invasores, mientras que la respuesta de tipo humoral requiere tiempo, ya que se precisa la sínteis de los compuestos inmunes (Lackie 1988).
Células y tejidos que participan en la respuesta inmune
La constante necesidad de vigilar la presencia de partículas extrañas en diferentes sitios del hemocele hace necesario que las células y tejido involucrados en la respuesta inmune estén distribuidos a través del cuerpo. Aunque los hemocitos constituyen las células involucradas en la respuesta por excelencia, se ha determinado que existen otras células y tejidos cuya participación es efectiva en este proceso; la cutícula y la membrana peritrófica por ejemplo, se comportan como barreras físicas que regulan la entrada y dispersión de patógenos y parásitos; de hecho, la mayoría de microorganismos producen gran cantidad de enzimas que les permiten degradar estas barreras.
Igualmente, la capa epitelial del integumento, las glándulas salivales y hasta componentes del sistema reproductor, han sido identificados como productores de sustancias con acción antibacterial jugando un papel activo en la respuesta inmune; es así como las cecropinas, uno de los péptidos antibacteriales mejor estudiados en insectos, son producidos por las células epidermales de Bombyx mori cuando la cutícula es alterada (Dunn 1990; Boman 1991; Paskewitz y Christensen 1996; Lorena y Maung 1996).
La hemolinfa, sin embargo, es el tejido funcional responsable de la mayoría de las respuestas inmunes en insectos, está formada por los hemocitos y el plasma; los hemocitos son las células que constituyen la primera línea de defensa contra los patógenos invasores y circulan libremente en el hemocele (Lackie 1988).
La clasificación de los hemocitos se ha realizado a partir de estudios morfológicos ultraestructurales y más recientemente, gracias a técnicas que han permitido su fraccionamiento por gradientes de densidad, se han caracterizado las diferentes poblaciones celulares (Miranda y Heusden 1996; Drift y Brehelin 1993). La producción de anticuerpos monoclonales, dirigidos contra determinantes de la superficie celular, ha permitido incrementar el conocimiento sobre estas células y en la actualidad se ha logrado, incluso, identificar subpoblaciones de los diferentes tipos y precisar su participación activa en cada respuesta (Lackie 1988).
En general, se conocen cinco tipos de hemocitos: prohemocitos, plasmatocitos, células granulares, células esféricas y eunocitoides (Fig. 1) (Drift y Brehelin 1993).

Representación esquemática de los hemocitos de un lepidóptero. a.prohemocito, b.granulocito, c.plasmatocito, d.coagulocito, e.adipohemocito, f.esperulocito, g.célula eunocitoide (Modificado de Miranda y Heusden 1996).
Los
Los
Los
Las funciones de otras células como
En la literatura aparecen descritos otros tipos celulares como
A pesar de que la mayoría de los hemocitos circulan libremente en el hemocele, en los insectos al igual que en los mamíferos, se han encontrado hemocitos infiltrantes, es decir, que pueden invadir tejidos; estas células se consideran de gran importancia para la defensa contra parásitos que no están libres en la cavidad hemocélica (Paskewitz y Christensen 1996).
Aunque al interior del insecto los hemocitos atacan los agentes invasores y contribuyen a la producción de sustancias con efectos antimicrobiales, existe un tejido que constituye el principal sitio de inducción de estas proteínas y participa activamente en la respuesta inmune, este es el cuerpo graso, un tejido adiposo que en los insectos cumple funciones análogas a las del hígado en mamíferos y aunque este tejido no interactúa directamente con los microorganismos para destruirlos, al parecer, algunas señales que son transmitidas desde el sitio de infección inducen en sus células una respuesta (Faye y Hultmark 1993; Hultmark 1993; Miranda y Heusden 1996).
Mecanismos de acción
Respuesta inmune de tipo celular
Fagocitosis
Este es el principal mecanismo de defensa observado hacia partículas pequeñas como bacterias, esporas de protozoos y partículas abióticas.
En este tipo de respuesta, que ocurre con frecuencia, el microorganismo o la partícula abiótica presente en el hemocele del insecto es unido a una célula fagocítica o hemocito y posteriormente ingerido por ésta, mediante filopodias o vesículas cubiertas (Bayne 1990; Gotz y Bomman 1985).
El
En el caso de vertebrados se sabe que el reconocimiento se efectúa gracias a factores quimiotácticos, mientras que en insectos ha sido difícil probar que estos factores sean necesarios, ya que durante la fagocitosis de objetos inertes como vidrio o látex, observada in vitro, los factores quimiotácticos no desempeñarían ningún papel (Faye y Hultmark 1993; Hultmark 1993).
Algunos investigadores han sugerido que para el reconocimiento, es suficiente la gran oportunidad de que ocurra el contacto al azar entre células y partículas extrañas, por el libre movimiento de los hemocitos en una cavidad abierta (Paskewitz y Christensen 1996).
La adhesión, posterior al reconocimiento, es mediada en el caso de vertebrados por
En invertebrados se han propuesto como opsoninas las
Sin embargo, en mosquitos de los géneros Anopheles y Aedes, infectados con microfilarias, se demostró que títulos altos de hemaglutininas facilitaron el movimiento de los nemátodos desde el intestino al hemocele y la formación de cápsulas y melanización de los mismos (Nayar y Knight 1997).
Recientemente se ha postulado que la unión fagocito - agente invasor, ocurre directamente a través de receptores presentes en la membrana de los hemocitos (Paskewitz y Christensen 1996).
Formación de Nódulos
Aunque las pequeñas infecciones con bacterias u otros microorganismos son resueltas por fagocitosis, la respuesta hacia infecciones mayores está representada por la formación de nódulos.
Los nódulos son asociaciones de hemocitos que se agrupan en torno a las partículas extrañas y las atrapan limitando su acción (Lackie 1988).
Al parecer, la hemolina, una proteína inducida en gran cantidad en insectos después de ser inyectados con bacterias, funciona como un modulador de los hemocitos estimulando su migración para formar nódulos y prevenir la excesiva agregación de los mismos (Paskewitz y Christensen 1996).
Encapsulación
Cuando las partículas invasoras en el hemocele del insecto son demasiado grandes para ser fagocitadas, los hemocitos forman cápsulas multicelulares al rededor de ellas; estas cápsulas posteriormente son cubiertas por un pigmento negro conocido comúnmente como melanina.
La encapsulación ha sido descrita en saltamontes y mosquitos como estrategia para eliminar huevos de parasitoides, larvas de helmintos e infecciones con parásitos microsporidias (Lackie 1988; Dunn 1990).
En algunos insectos, especialmente dípteros vectores que poseen bajo número de hemocitos, la melanización de los agentes invasores puede ocurrir antes de la formación de la envoltura o cápsula; este proceso ha sido llamado encapsulación humoral melanótica; en este tipo de respuesta, los hemocitos sólo participan en la formación inicial de la cápsula (Dunn 1990).
Melanización y Sistema PROPO
La melanización, que acompaña los procesos de respuesta como encapsulación y encapsulación humoral melanótica, depende de la producción o absorción de unas enzimas llamadas fenol oxidasas (PO); estas enzimas existen en la mayoría de organismos incluyendo hongos, plantas y animales (Lackie 1988).
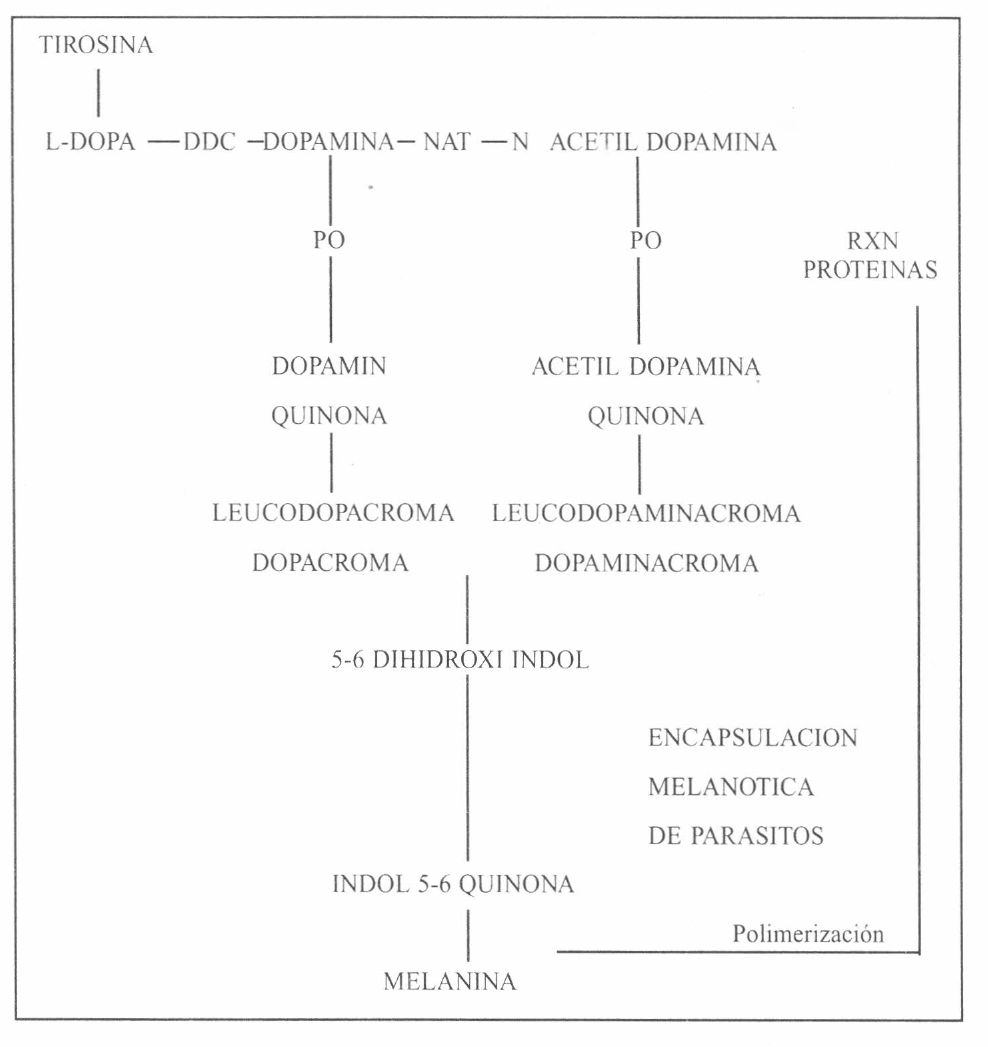
Vía Bioquímica para la formación de melanina durante la encapsulación de parásitos. Modificada de Paskewitz y Christensen (1996). La encapsulación es iniciada por la hidroxilación de la tirosina a L-dopa mediante la fenol oxidasa (PO). La decarboxilación de la L-dopa es llevada a cabo posteriormente por la dopadecarboxilasa (DDC) para formar dopamina. La dopamina es acetilada por la actividad enzimática de la N-acetil transferasa (NAT) para formar N-acetil dopamina. La fenol oxidasa actúa sobre la L-dopa, dopamina y N acetil dopamina y produce sus respectivas quinonas las cuales sufren rearreglos estructurales para formar la indol 5-6 quinona cuya polimerización resulta en la formación de melanina. Las dopa pueden reaccionar con grupos nucleofílicos de proteínas y con los hemocitos y polímeros de melanina se forman las cápsulas que secuestran los parásitos.
En insectos las fenoloxidasas, además de participar en el sistema inmune, están involucradas en la melanización y esclerotización de la cutícula y de los huevos, así como en la cicatrización de heridas.
En la hemolinfa existe un tipo particular de fenoloxidasa que se encuentra como una proenzima llamada profeniloxidasa; esta proenzima puede ser activada por componentes del microorganismo invasor y desencadenar una cascada enzimática conocida como
Insectos comúnmente utilizados para estudiar Inmunología de Insectos y Parásitos asociados con ellos
Modificada de Lackie (1988).
La cascada enzimática, PROPO, ha sido bien estudiada en insectos como Bombyx mori, donde se ha observado que los B - glucanos presentes en la pared celular de hongos, tienen la propiedad de activar el mecanismo.
Una vez que los hemocitos (granulocitos) están en contacto con el invasor, se degranulan liberando componentes que antes estaban inactivos en la hemolinfa y al cabo de una serie de reacciones se forman los polímeros de melanina que, junto con los hemocitos, forman las cápsulas que finalmente secuestran los parásitos (Paskewitz y Christensen 1996).
In vitro el sistema PROPO puede ser activado por numerosos agentes, incluyendo isopropanol, detergentes, ácidos grasos, cloroformo, productos microbiales y partículas abióticas, requiriéndose calcio para tal activación.
In vivo el sistema es probablemente activado por serin proteasas, las cuales han sido purificadas a partir de muchos insectos; estas proteasas están estructuralmente relacionadas con el sistema de complemento de mamíferos y se ha sugerido que las cascadas proteolíticas que estimulan podrían tener un origen común (Hultmark 1993).
Las serin proteasas presumiblemente existen en la hemolinfa como precursores inactivos que a su vez pueden ser activados por otras proteasas; ciertos productos microbiales al reaccionar con los primeros componentes o proteasas, causan cambios conformacionales que originan la activación de la proenzima fenol oxidasa PO y por ende del sistema PROPO (Paskewitz y Christensen 1996).
Recientes investigaciones han demostrado que la capacidad de melanización y la activacón del sistema PROPO, dependen de factores como el estado nutricional, la temperatura y el tamaño del cuerpo del insecto (Suwanchaichinda y Paskewitz 1998).
Es importante anotar que en la hemolinfa de muchas especies de insectos se han encontrado inhibidores de la PO que pueden modular su actividad, la razón de ello probablemente la constituye el hecho de que esta enzima produce quinonas de corto tiempo de duración, que in vivo podrían resultar tóxicas al insecto y por lo tanto la regulación en la activación de este sistema es esencial.
Finalmente, las fenoloxidasas son producidas en tejidos como la epidermis y la hemolinfa, aunque se ha descrito que células del músculo torácico, intestino medio y tubos de malpighi poseen capacidad de melanizar nemátodos intracelulares sugiriendo en ellos su producción (Soderhall y Aspan 1993).
Dopa Decarboxilasa
Como las fenol oxidasas, la dopadecarboxilasa (DDC) es una importante enzima que participa en la esclerotización y melanización de los huevos y la cutícula de los insectos y también se encuentra en el sistema nervioso, donde genera el neurotransmisor dopamina.
Algunos estudios recientes con moscas Drosophila mutadas para la dopadecarboxilasa con actividad temperatura dependiente, mostraron una importante disminución en la habilidad de las moscas para melanizar parasitoides en rangos de temperaturas altas a las cuales la enzima no era funcional, comprobando así la participación de esta enzima en el sistema inmune; igualmente, se ha observado que la actividad de la DDC aumenta significativamente cuando ocurren infecciones con filarias en mosquitos(Paskewitz y Christensen 1996).
Respuestas de tipo humoral
Las investigaciones iniciales sobre el sistema inmune de insectos indicaron la existencia de reacciones de gran magnitud cuando éstos eran inyectados con bacterias (Faye y Hultmark 1993).
Además de las reacciones celulares, gracias a las cuales patógenos humanos como los agentes de neumonía, disentería y cólera eran fagocitados o encapsulados por los hemocitos de los insectos retados, se detectó la presencia de un gran número de proteínas y péptidos bactericidas que se acumulaban en la hemolinfa; estas sustancias constituyen en la actualidad el aspecto más importante de la respuesta inmune de tipo humoral.
Aunque inicialmente se realizaron investigaciones con insectos pequeños, los resultados más significativos fueron obtenidos a partir de los estudios efectuados con el saltamontes gigante Hyalophora cecropia hacia 1975; estos estudios generaron importantes resultados que motivaron a un buen número de científicos a investigar fenómenos similares en otros grupos de insectos (Tabla 1) (Kunihiko et al. 1992; Wiesner 1993; Hultmar 1993; Lockey y Ourth 1996; Kang et al. 1996; Bullet et al. 1992).
La inducción de
Proteínas inmunes caracterizadas a partir de insectos
Modificada de Faye y Hultmark (1993).
Las primeras proteínas antibacteriales descritas en insectos fueron las lisozimas, cecropinas y atacinas; sin embargo, en la actualidad existen más de cincuenta factores que han sido caracterizados y clasificados en pocos grupos (Tabla 2) (Dunn 1990; Bullet et al. 1992; Hoffman y Hetro1992; Hultmark 1993). Con excepción de las lisozimas, las cuales están estrechamente relacionadas con las lisozimas tipo c de vertebrados, estas moléculas son factores nuevos descubiertos inicialmente en insectos (Hultmark 1993).
Cecropinas
Constituyen una familia de péptidos con treinta y cinco a treinta y nueve residuos de aminoácidos; fueron llamadas cecropinas por Hyalophora cecropia, el insecto a partir del cual fueron aisladas por primera vez.
A concentraciones muy bajas, de micromoles, estos péptidos pueden lisar y matar la mayoría de las bacterias con las cuales los insectos estudiados han sido retados y su actividad ha sido comprobada para bacterias Gram + y Gram -.
Aunque las cecropinas fueron inicialmente descubiertas en insectos, recientes estudios han mostrado que estos péptidos pueden también ocurrir en vertebrados y podrían estar relacionados filogenéticamente (Faye y Hultmark 1993; Hultmark 1993).
En insectos las cecropinas han sido encontradas en muchos Lepidoptera y en la mayoría de los Diptera incluyendo Drosophila, Aedes aegypti, Simulium, Anopheles, Sarcophaga y Glossina spp. (Beck y Habitch 1994; Paskewitz y Christensen 1996).
Las cecropinas son moléculas altamente anfipáticas que interactúan con las membranas lipídicas creando canales de iones o poros de diferentes tamaños; aparentemente, estos poros son formados en las membranas internas de las bacterias con la consecuente lisis celular (Faye y Hultmark 1993).
Los niveles de cecropinas encontrados en la hemolinfa de insectos como Drosophila oscilan entre 25-50 µm en moscas inmunizadas, mientras que en Hyalophora la concentración total ha sido estimada alrededor de 75µm.
Solo algunos patógenos de insectos como Bacillus thuringiensis y Serratia marcescens son resistentes a la cecropina a estos niveles; a concentraciones mayores de 100 µm algunos parásitos eucarióticos como Trypanosoma, Plasmodium y Leishmania -todos ellos patógenos del hombre y animales transmitidos por insectos han sido destruidos (Hultamrk 1993; Faye y Hultmark 1993; Paskewitz y Christensen 1996).
Algunos experimentos con cecropinas han permitido incluso, curar mosquitos transmisores de malaria infectados con el parásito, mediante inyecciones con estos péptidos a concentraciones muy altas (Paskewitz y Christensen 1996).
En la mayoría de insectos las cecropinas son expresadas en el cuerpo graso de larvas y adultos, aunque también en estado de pupa; adicionalmente, algunas han sido encontradas en hemocitos (Faye y Hultmark 1993).
Defensinas
Las defensinas son péptidos catiónicos de 4 Kda que contienen seis residuos de cisteína; estas moléculas se han aislado de insectos de los órdenes Diptera, Coleoptera y Odonata (Hultmark 1993; Faye y Hultmark 1993).
Las defensinas fueron llamadas de esta forma por su aparente similitud con las defensinas, un grupo de péptidos líticos encontrados en macrófagos y neutrófilos de mamíferos, pero trabajos recientes han señalado que las estructuras tridimensionales de ambas moléculas son diferentes y que las defensinas de insectos están más estrechamente relacionadas con las carbidotoxinas, toxinas derivadas de los escorpiones que funcionan como bloqueadores de canales de potasio (Hultmark 1993).
Aunque en algunos insectos las defensinas son producidas exclusivamente por los hemocitos, en otros como en Phormia, son sintetizadas por el cuerpo graso (Faye y Hultmark 1993).
Las defensinas poseen mayor actividad hacia bacterias Gram+ que Gram- y, de forma similar a las cecropinas, atacan la membrana celular de la célula bacterial (Hoffman y Hetro 1992; Bullet et al. 1993; Boman 1993).
Una defensina llamada royalisina ha sido aislada recientemente a partir de jalea real, un producto de las glándulas faríngeas de abejas usado para alimentar las larvas destinadas a ser reinas (Paskewitz y Christensen 1996).
Lisozimas
Las lisozimas fueron los primeros factores antibacteriales identificados en la hemolinfa de insectos; se sugiere que debieron aparecer temprano en la historia de la evolución ya que han sido encontradas en bacteriófagos, plantas, vertebrados e invertebrados (Hultmark 1993; Lockey y Ourth 1996).
Son proteínas entre 14-17 Kda que tienen efecto sobre algunas bacterias Gram+, estas enzimas rompen los péptido glicanos que forman la pared celular de bacterias y en estudios realizados en insectos como Hyalophora se ha hipotetizado que las lisozimas sirven principalmente para remover los restos de pared que quedan después de la acción de las cecropinas (Faye y Hultmark 1993).
Se han purificado lisozimas a partir de la hemolinfa de numerosos insectos como Galleria, Manduca, Locusta, Spodoptera, Calliphora y Drosophila. Como en el caso de las demás proteínas inmunes, la principal fuente de lisozima en la hemolinfa es el tejido graso, aunque se han encontrado bajos niveles en hemocitos (Paskewitz y Christensen 1996).
Las lisozimas también se han encontrado en células pericardiales y tracto intestinal y en insectos como Manduca son secretadas específicamente durante determinadas etapas de la metamorfosis (Faye y Hultmark 1993).
Diptericinas
La superfamilia de las diptericinas incluye las attacinas, sarcotoxina, diptericinas, apidacina y coleoptericina (Paskewitz y Christensen 1996).
Las
Attacinas han sido también encontradas en insectos como Glossina, Drosophila y Aedes aegypti (Paskewitz y Christensen 1996).
La
Las
Otras sustancias antibacteriales ricas en prolina encontradas en Hymenoptera, son las
La
Las
La
Hemolina
La hemolina es una proteína inmune de 48 Kda que existe normalmente en la hemolinfa de insectos, pero cuya cantidad incrementa ante la presencia de bacterias y otros microorganismos (Faye y Hultmark 1993).
Esta proteína es sintetizada en los hemocitos y el cuerpo graso y aparece como una importante mediadora en el proceso de fagocitosis, ya que puede reconocer y opsonizar partículas extrañas para después unirse a los fagocitos encargados de destruir tales partículas (Paskewitz y Christensen 1996; Bettencourt et al. 1997).
Recientemente, la hemolina ha sido descrita como miembro de la familia de las inmunoglobulinas las cuales son proteínas inmunes presentes en mamíferos que poseen gran especificidad para unirse a los antígenos, es decir, a las sustancias extrañas o moléculas que generan una respuesta inmune.
Al parecer, la hemolina está estrechamente relacionada con las extensiones celulares y moléculas de membrana involucradas en las interacciones célula - célula y de ahí sus importantes funciones en el proceso de reconocimiento (Hultmark 1993).
Péptidos Antifungales
En insectos se conocen pocos factores con actividad dirigida hacia hongos; sin embargo, recientemente se ha purificado una proteína antifungal a partir de la hemolinfa de Sarcophaga peregrina; esta proteína posee 67 aminoácidos y es expresada constitutivamente; se ha demostrado que su actividad antifúngica aumenta en presencia de péptidos antibacteriales inducibles como las sarcotoxinas (Paskewitz y Christensen 1996).
La
Citoquinas
Las citoquinas son polipéptidos liberados por las células que participan en el sistema inmune y son ampliamente reconocidas en mamíferos. Estas sustancias poseen propiedades similares a las hormonas, afectan numerosos sistemas de órganos involucrados en la defensa y son de gran importancia en la comunicación celular (Watson et al. 1987).
Recientemente, en Drosophila se ha descrito un sistema que controla la expresión de genes durante la respuesta inmune, este sistema conocido como la vía regulatoria dorso ventral "Toll", muestra gran similitud con la cascada de citoquinas durante la respuesta inmune en mamíferos (Nicolás et al. 1998; Lemaitre et al. 1997).
Igualmente, se han aislado y caracterizado, a partir de invertebrados, moléculas muy similares a la interleuquina uno (Il-1) y el factor de necrosis tumoral (TNF) dos importantes citoquinas presentes en mamíferos, proponiéndose que estas sustancias podrían estar mediando las reacciones inmunes; sin embargo, existen pocos estudios al respecto (Beck y Habitch 1994; Paskewitz y Christensen 1996; Han et al. 1998).
Mecanismos antivirales
Aunque un gran número de virus ataca insectos se conocen pocos detalles sobre los mecanismos inmunes para destruirlos.
El intestino es la principal barrera para limitar la invasión de partículas virales en el hemocele y se ha estimado que gracias a él, la infectividad puede reducirse hasta en un 50-60% (Pakewitz y Christensen 1996).
Algunos estudios realizados en cultivos celulares de insectos como Aedes aegypti y Bombyx mori infectados con virus, no indicaron la presencia de ningún tipo de mecanismo de respuesta y similares resultados fueron obtenidos con larvas de Trichoplusia ni infectadas con baculovirus; no obstante, se ha descrito actividad antiviral en la hemolinfa e intestino de Bombyx mori y se ha observado que algunas partículas virales pueden ser ocasionalmente fagocitadas (Hultmark 1993).
Mecanismos de respuesta hacia nemátodos
La encapsulación es uno de los principales mecanismos de respuesta hacia nemátodos los cuales tardan usualmente de tres a cuatro días en morir después de ser encapsulados; también se ha observado melanización humoral melanótica y respuestas de otros tejidos como el epitelio intestinal y las células del tejido graso, aunque se presume la acción de sustancias específicas que puedan actuar contra nemátodos, no se ha aislado ninguna (George y Poinar 1979).
Después de revisar los hallazgos más importantes, en lo que constituye la batería de proteínas inmunes que poseen los insectos, es necesario precisar que por su diversidad e importante función como mecanismo de respuesta inmune en insectos, estas proteínas inducibles como los genes que las regulan han sido ampliamente estudiadas (Hultmark 1993; Faye y Hultmark 1993).
Las proteínas sintetizadas y liberadas en la hemolinfa de insectos infectados con bacterias han sido visualizadas mediante inyección de aminoácidos radioactivos junto con las bacterias y posterior separación de la hemolinfa en geles de poliacrilamida SDS.
La inyección de inhibidores de RNA y síntesis de proteínas han permitido entender que la mayoría de las proteínas inmunes de insectos son reguladas a nivel trasncripcional y gran cantidad de ensayos biológicos han facilitado la purificación de las sustancias así como la construcción de librerías de cDNA inmune específicas (Hultmark 1993).
Se han clonado, por ejemplo, los genes de Drosophila que codifican para las cecropinas, diptericinas y los siete genes que codifican para las lisozimas; estudios a este nivel sugieren que las proteínas inmunes son codificadas por familias de genes estrechamente relacionadas, pero que muestran diferentes patrones de expresión; es así como algunos son preferiblemente expresados durante algunas etapas de la metamorfosis (Hultmark 1993; Faye y Hultmark 1993; Paskewitz y Christensen 1996).
Reconocimiento
Un insecto puede reconocer como extraño a un gran número de partículas de origen biótico o abiótico pero, ¿cómo ocurre este proceso? ¿Qué factores del invasor son reconocidos y cuáles componentes del sistema inmune del insecto median el reconocimiento?
El sistema inmune en insectos ha sido descrito como un sistema sensitivo que puede reconocer diferentes aspectos del invasor y diferentes componentes de este sistema se encuentran involucrados; este proceso es realmente complejo y es uno de los principales objetos de investigación en la actualidad.
Las características de la superficie de los patógenos y parásitos así como algunos productos liberados por ellos, constituyen, al parecer, las bases del reconocimiento (Hultmark 1993).
Las características de la superficie incluyen presencia de carbohidratos, lípidos, proteínas y carga de las partículas; esta última en especial para materiales abióticos ya que se ha encontrado que los insectos encapsulan y melanizan pequeños implantes de sephadex cuando éstos están cargados positivamente.
Para entender la importancia de la carga eléctrica se ha propuesto que compuestos lábiles del plasma pueden ser estabilizados al entrar en contacto con superficies de cierta carga y actuar como opsoninas (Paskewitz y Christensen 1996).
Algunos factores del sistema inmune con participación activa en reconocer partículas extrañas de origen biótico incluyen activadores del sistema PRORO hemaglutininas o lectinas y proteínas específicas que se unen a los B1-3 glucanos y/o polisacáridos.
En cuanto a los activadores del sistema PROPO se sugiere que algunos pequeños fragmentos liberados por el clivaje de las serín proteasas pueden servir como opsoninas a parásitos y patógenos facilitando el reconocimiento (Hultmark 1993).
Las lectinas son glicoproteínas que se unen específicamente a cadenas de carbohidratos y están presentes en la mayoría de los invertebrados estudiados. Entre las funciones atribuidas a las lectinas se encuentran promover la ingestión de tejidos propios por los hemocitos cuando el insecto está en etapa de pupación y servir como opsoninas de los invasores facilitando la adherencia y posterior fagocitosis llevada a cabo por los hemocitos (Soderhal y Aspan 1993; Faye y Hultmark 1993).
Las lectinas pueden también estimular los hemocitos a degranularse, originando la coagulación y liberación de los componentes de la cascada PROPO y se unen fácilmente a bacterias, protozoos y metazoos.
Finalmente, estas proteínas se encuentran libres en la hemolinfa, intestino medio y membranas celulares y pueden ser producidas por el tejido graso o los hemocitos (Faye y Hultmark 1993).
Han sido descritas dos proteínas específicas a partir de Blaberus y Bombyx, que se unen a B-13 glucanos; estas proteínas reconocen los glucanos presentes en paredes de hongos y algas aumentando en su presencia la actividad de la PO. Se han encontrado receptores para estas proteínas en los hemocitos los cuales, al entrar en contacto con la proteína, se diseminan y degranulan parcialmente.
Adicionalmente, se ha purificado a partir de Bombyx mori una proteína que reconoce los péptido-glicanos presentes en las paredes celulares de las bacterias; estos polisacáridos probablemente estimulan el sistema PROPO así como la síntesis de proteínas antibacteriales, pero aún no ha sido bien documentado (Hultmark 1993).
Como se discutió previamente, la hemolina juega también un papel importante en el proceso de reconocimiento.
Evasión del sistema inmune
Gran cantidad de microorganismos, incluyendo patógenos y parásitos, pueden evadir los mecanismos de respuesta inmune presentes en sus insectos hospederos; esto ha sido comprobado en hongos, nemátodos, protozoos, bacterias, espiroquetas y virus (Paskewitz y Christensen 1996).
La transmisión de parásitos nocivos para el hombre y animales y el interés en la utilización exitosa de agentes de control microbiológico constituyen las áreas que con mayor énfasis han generado investigaciones en este sentido.
La sobrevivenčia de los parásitos en los insectos que los transmiten requiere mecanismos de evasión bien estructurados y entender estos mecanismos precisa también comprender el mecanismo inmune que debe ser evadido; pocos estudios han sido realizados con este fin y los que existen han proporcionado resultados que apenas empiezan a ser correctamente dilucidados.
Se han propuesto algunas estrategias utilizadas por los parásitos y patógenos que incluyen la
Entre las estrategias de supresión se encuentra la producción de proteínas que bloquean elementos claves en el sistema PROPO o sustancias que tienen efectos inhibitorios sobre las proteínas inmunes producidas por el insecto.
En el caso de bacterias patógenas de insectos como Bacillus thuringiensis se ha observado que éstas producen inhibidores que actúan por clivaje proteolítico sobre las cecropinas y attacinas suprimiendo su acción antibacterial (Paskewitz y Christensen 1996).
Igualmente, se han observado asociaciones simbióticas entre varios microorganismos que aseguran la sobrevivencia de éstos al interior del insecto; por ejemplo, la bacteria Xenorhabditis nematophilus se asocia con un nemátodo que frecuentemente parasita insectos, el nemátodo ayuda a la bacteria produciendo una enzima proteolítica que degrada las cecropinas y attacinas producidas por el insecto y la bacteria a su vez ayuda al nemátodo al parasitar conjuntamente al insecto (George y Poinar 1979).
En las estrategias de
De otra forma, los hemocitos de insectos pueden servir como células hospederas de parásitos y patógenos disminuyendo la respuesta inmune; por ejemplo, Trypanosoma rangeli, el agente causal de la Tripanosomiasis o enfermedad de Chagas, vive en los hemocitos de su insecto transmisor Rhodnius prolixus.
El hecho de que estos parásitos se desarrollen al interior de las células los aisla del contacto y reconocimiento de hemocitos circulantes (Paskewitz y Christensen 1996).
La producción de metabolitos tóxicos por hongos entomopatógenos, con capacidad de suprimir la respuesta inmune de los insectos, ha sido bien documentada en el caso de hongos como Beauveria bassiana y Metarhizium anisopliae; la beauvericina y destruxinas producidas por estos hongos al interior del insecto, actúan directamente sobre los hemocitos evitando reacciones como la encapsulación y fagocitosis (Roberts 1981; Vey et al. 1985; Kopecki et al. 1992).
Resistencia
En cuanto a la resistencia de insectos a parásitos y patógenos se conoce poco, aunque se considera un aspecto crítico en contextos como el control de insectos transmisores de enfermedades.
A pesar de la selección en el laboratorio de varias cepas de mosquitos resistentes a Plasmodium, agente causante de la malaria, el mecanismo de resistencia no es aún claro; en estos insectos la resistencia ha sido expresada como la habilidad del insecto para encapsular y melanizar diferentes estados del parásito: ookinetes y oocistos (Paskewitz y Christensen 1996).
Algunos investigadores sugieren que los esporozoitos de Plasmodium y otros parásitos transmitidos por insectos dípteros pueden sobrevivir en el hemocele de estos insectos en virtud de sus superioridad numérica, pero se conoce que existen parásitos que pueden evadir la encapsulación, por lo tanto parecen poseer su propio mecanismo de defensa (Lackcie 1990).
Estudiar las diferencias bioquímicas y genéticas entre una cepa susceptible y una resistente seleccionada en el laboratorio es una alternativa que podría proveer gran cantidad de información sobre las bases moleculares de la resistencia como herramienta para estudiar el control genético de la capacidad vectorial y la utilización exitosa de agentes de control microbiológico a los cuales algunos insectos pueden ser resistentes.
Así mismo, el desarrollo de mapas genéticos, de gran auge en la actualidad, constituye una importante oportunidad para identificar aquellas regiones del genoma que determinan las respuestas en insectos resistentes.
Estudios futuros
Dada la complejidad del sistema inmune de insectos, aunque mucho se ha avanzado en este campo, existe un gran trabajo por hacer y cada uno de los aspectos anteriormente revisados ofrece innumerables posibilidades.
En cuanto al reconocimiento, las moléculas que lo favorecen están mejor identificadas en el caso de hongos y bacterias y menos en virus, nemátodos y protozoos y queda aún por establecer el papel definitivo de moléculas como las lectinas.
Los componentes bioquímicos, vías de producción y mecanismos de acción de muchas proteínas inmunes son desconocidas y quedan muchas otras de estas proteínas por identificar así como muchos genes que las regulan por aislar.
Las señales de transducción y la comunicación célula-célula que permiten la realización de todos los procesos de respuesta inmune, los procesos fisiológicos involucrados y las interacciones con otros sistemas sólo empiezan a estudiarse hasta ahora y constituyen un vasto campo para la investigación.
La relación con la inmunidad de vertebrados comprende un tópico de gran interés y se han generado hipótesis como el hecho de que ambos sistemas inmunes podrían haber evolucionado a partir de un ancestro común.
La Entomología puede obtener en el estudio de la inmunología de insectos numerosos conocimientos prácticos en áreas como el control microbiológico de insectos; aspectos como el mecanismo de defensa desarrollado por el insecto plaga ante el microorganismo a utilizar, el tiempo que tarda esta respuesta en ser efectiva y cuales relaciones se establecen entre el insecto y el parásito/patógeno, podrían indicar el potencial de un determinado agente de control microbiológico.
Una respuesta inmune fuerte puede reducir o restringir la capacidad vectorial de algunos hospederos de patógenos para el hombre y los animales así como respuestas débiles pueden aumentar la habilidad de agentes infecciosos en insectos; entender los mecanismos por los cuales los parásitos evaden estas respuestas podrían sugerir mecanismos para ser explotados en el control de estos parásitos.
Por último, con la disponibilidad de herramientas como cultivos celulares de insectos, tanto de células epiteliales como hemocitos, se vislumbra la oportunidad de estudiar in vitro detalles de la respuesta inmune y entender este proceso a nivel molecular.