
Abstract
La acelerada destrucción de los ecosistemas colombianos y el gran desconocimiento de la biota hacen necesario el uso de métodos que permitan la identificación rápida de zonas con gran riqueza, endemismos o fragilidad ecológica para incluirlas en proyectos de conservación o para la identificación del grado de alteración antrópica que presenten. Las mariposas, al igual que algunos taxa de la entomofauna asociada con el suelo, han sido utilizadas como indicadores de tales zonas; el uso de los Rhopalocera puede ser más ventajoso por involucrar un gran número de elementos por hábitat y ser fáciles de identificar y determinar en el campo, pero, ¿Compensa la facilidad de uso de las mariposas la mayor resolución que como indicadores de diversidad biótica brinda la comunidad de insectos asociados con el suelo? Con el fin de comparar la correspondencia entre la diversidad estructural de la vegetación y la diversidad faunística de los dos grupos de insectos, se estudiaron las comunidades asociadas con tres diferentes tipos de vegetación -Bosque Amazónico pluvial de suelos arenosos, Caatinga y Sabana- presentes de la Serranía de Taraira (Vaupés, Colombia), durante seis semanas. Para la captura de las mariposas se usó la jama en tres transectos que incluían los tres tipos de vegetación, y trampas Van Someren-Rydon con banano, melaza y jabón como cebo; la entomofauna se coleccionó por medio de trampas "Pitfall", colocadas dentro de una cuadrícula de 50x50 m (dos cuadrículas por cada tipo de vegetación y tres subparcelas de 5x5 m con cinco trampas por cuadrícula). Para mariposas se utilizó un análisis de tipo descriptivo y para la artropofauna un análisis cuantitativo. Los resultados obtenidos en los dos grupos son coincidentes, siendo la comunidad de Bosque la más diversa, lo cual coincide con la mayor complejidad estructural y diversidad taxonómica de su vegetación. Los tres tipos de vegetación se comportan como sistemas independientes, aunque Caatinga se comporta más como una prolongación de la biota del Bosque Amazónico (menos diversificada y con menor dominancia relativa de algunos taxa, aunque con la presencia de elementos propios). Se destaca la alta correspondencia entre la biodiversidad de las comunidades de mariposas y la de artrópodos asociados con el suelo, ratificando la utilidad de las primeras como bioindicadores de fácil uso para prospecciones rápidas.
Introducción
Son muchos los factores que afectan la diversidad y composición de una comunidad animal; el conjunto de poblaciones que la integran debe responder a las características ambientales, y luego seleccionar o adaptarse a los microhábitats disponibles dentro de la estructura espacial de un hábitat dado; en tal sentido, es presumible que mientras más compleja sea la estructura vegetal de una comunidad, mayor será la disponibilidad de nichos y por tanto la riqueza y diversidad de especies (Anderson y Shugart 1974; Dinitz y Kitayama 1994). De manera más específica, la riqueza de una comunidad de mariposas depende principalmente de la diversidad florística, debido a la fuerte especificidad de huéspedes de estos insectos (Ehrlich y Raven 1964; Singer 1984). A esto se suma una fuerte estratificación debido a la influencia de gradientes de luz, viento, humedad, temperatura y altitud, y la influencia de otros factores como la disponibilidad de recursos alimentarios para larvas y adultos, la estacionalidad del recurso y los depredadores (Gilbert 1984; Rausher 1981; Adams 1985, 1986; Callaghan 1985; Brown 1991; Sparrow 1991; Kremen 1992; Fagua y Ruiz 1995). Como resultado, se tiene que la riqueza y composición de una comunidad de mariposas puede ser tomada como reflejo de la diversidad biótica y estructural de cualquier ecosistema terrestre.
Por su parte, la entomofauna asociada con el suelo también es afectada por buena parte de estos factores; sus poblaciones tienen componentes de varios grupos taxonómicos que incluyen, además de herbívoros, a xilófagos, saprófagos y depredadores. Igualmente puede tomarse como una referencia de la diversidad biótica y de microhábitats de una unidad paisajística, pero su poder de resolución es mayor, en comparación con las mariposas, debido a que incluye mayor número de taxa y niveles tróficos.
Dentro de este marco de referencia se incluye ahora el término indicadores biólogicos, que son grupos de especies o grupos taxonómicos que con su presencia pueden indicar el estado de la biota en cuanto a parámetros como biodiversidad y biogeografía o grado de intervención humana (Kremen et al. 1993). Se espera que los bioindicadores sean grupos usualmente abundantes, estables y preferiblemente sedentarios dentro de un ecosistema, que estén ecológicamente muy diversificados, sean fácilmente determinables y tengan una biología y taxonomía bien conocidas; su manipulación en campo y laboratorio debe ser sencilla y deben presentar ciclos de vida cortos y tener sensibilidad y fidelidad ecológica. Estas características se complementan si presentan áreas definidas de endemismos y centros de diversidad (Brown 1991; Coddington et al. 1991; Kremen et al. 1993; Colwell 1994; Pearson 1994).
El uso de especies indicadoras ahorra tiempo, dinero y esfuerzo en trabajos de prospección y vigilancia ecológica, constituyéndose en una herramienta de primera mano para el análisis ambiental. No obstante, el grado de confiabilidad de los mismos, y en especial de las mariposas, es algo que generalmente no se cuestiona. Este trabajo pretende comparar la efectividad de las mariposas como bioindicadores cualitativos del tipo de hábitat; labor realizada como un complemento al reconocimiento preliminar de la biota del norte de la Serranía de Taraira (Departamento del Vaupés, Colombia). Así, los resultados en Rhopalocera se contrastan con un análisis cuantitativo detallado de la entomofauna asociada con el suelo. Para esto se comparan las respectivas comunidades de insectos de tres unidades de vegetación (Bosque, Caatinga y Sabana).
Materiales y Métodos
La Serranía del Taraira está en el extremo sur del departamento del Vaupés, Colombia (Fig. 1); es de origen precámbrico y posee colinas graníticas bajas con alturas hasta de 375 msnm. La precipitación media multianual es de aproximadamente 4.000 mm (Estación Biológica de Caparú; mínimo mensual superior a los 100 mm) y el régimen es bimodal estacional con máximos en julio-agosto. La temperatura promedia es de 25,1° C.
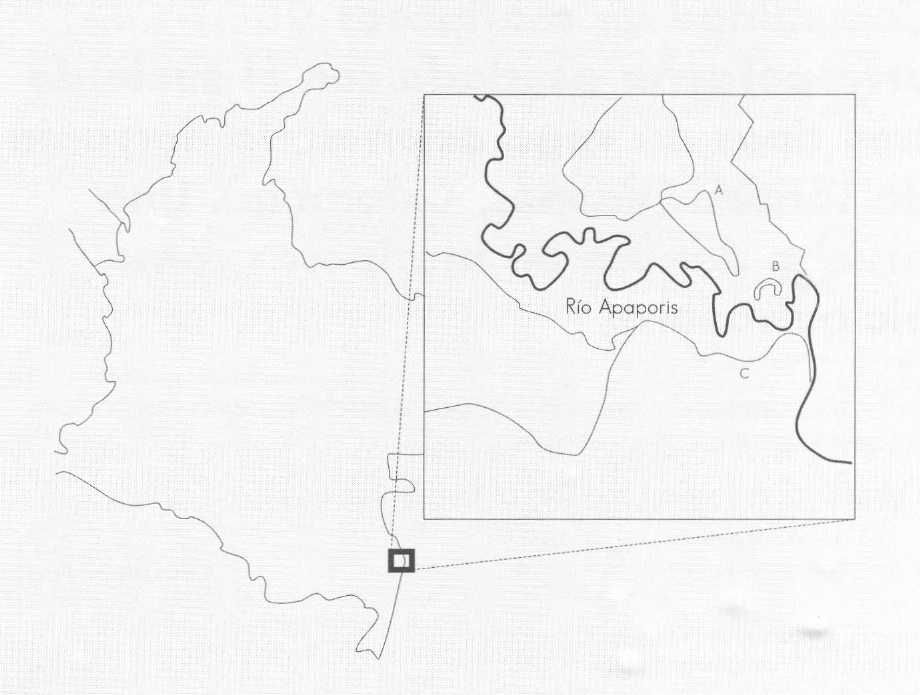
Mapa de la zona de estudio. a: Serranía de Taraira. b: Laguna de Taraira, Estación Biológica de Caparú. c: La Pedrera.
En el área de estudio se encontraron tres unidades diferentes de vegetación: «Bosque» (selva pluvial sobre suelos arenosos) (Domínguez 1985), que son bosques dominados por elementos arbóreos con DAP superiores a los 80 cm, dosel a los 25 m, con algunos elementos emergentes, interior oscuro y sotobosque bien diferenciado. «Caatinga», compuesta por árboles delgados con DAP entre los 5 y 25 cm, cuyo dosel alcanza los 20 m, interior medianamente claro, sotobosque muy tupido y proliferación de bejucos y musgos. «Sabana», bosques achaparrados con elementos que no superan generalmente los 12 m, con dosel no continuo o sin dosel y DAP entre 3 y 10 cm; es una unidad rica en orquídeas, líquenes y musgos. También se encontró una zona de explotación aurífera abandonada hace aproximadamente cinco años.
El trabajo se realizó entre julio y agosto de 1993, con una permanecia de seis semanas en la zona de estudio. La riqueza de mariposas se obtuvo del total de la captura o por la observación mediante dos procedimientos: Trampas Van Someren-Rydon y el uso de jama a lo largo de transectos. Para el primero se seleccionaron aleatoriamente tres sitios en cada una de las unidades paisajísticas y en cada uno se colocaron ocho trampas Van Someren-Rydon (con frutas mezcladas con melaza como cebo); las trampas permanecieron durante cuatro días y se revisaron diariamente. Para el segundo se marcaron tres transectos de 2 a 4 km que cruzaban áreas con los tres tipos de vegetación; en éstos se procedió a colectar las mariposas observadas. Cada transecto se recorrió entre las 9:00 y las 16:00 h, para un total de 210 horas de trabajo de captura de un colector (cabe destacar que se presentaron lluvias frecuentes y fuertes durante 16 de los 34 días de muestreo). La afinidad entre las comunidades de las tres unidades se comparó mediante los índices de Jaccard y Soremsem (confróntese Crisci y López 1983). La estimación de la abundancia fue descriptiva y se obtuvo según la frecuencia de las observaciones al seguir los transectos. Para este estudio se definió: Abundante: más de 10 observaciones por día; Común: 10 a 5 observaciones por día; Escasa: 5-1 observaciones por día; y Rara menos de 1 observación por día.
Para la artropofauna asociada con el suelo se escogió una parcela de 50 x 50 m en cada unidad paisajística, seleccionando aleatoriamente dentro de ésta tres subparcelas de 10 x 10 m, en las cuales se colocó en el centro y en cada uno de los extremos, un recipiente plástico con una solución acuosa de 8 partes de etanol 70 %, 2 de formol 5% y detergente (5 trampas Pitfall en total por subparcela). Las trampas se dejaron durante 8 días. Para cada parcela y sus respectivas subparcelas se obtuvo el índice de diversidad de Shannon-Weaver y el índice de Simpson, para tener una aproximación de la «concentración dominante» de especies (Hair 1987). Para conocer las afinidades entre las diferentes subparcelas se hizo una matriz de presencia-ausencia de los taxa, de la cual se obtuvo el coeficiente de similitud de Jaccard y los dendrogramas correspondientes mediante el uso del programa NTSYS- pc, versión 1.70 (1992 Applied Biostatics, Inc).
Resultados
Mariposas:
Heliconiinae y Morphiinae fueron las únicas subfamilias con un número de especies similar en las tres unidades comparadas; en tanto que Hesperiidae, Papilionidae, Nymphalinae, Satyrinae y Riodinidae fueron más diversas en «Bosque». Lycaenidae presentó mayor número de especies en «Caatinga» y «Sabana», en tanto que ejemplares de Ithomiinae y Brassolinae prácticamente sólo se registraron en «Bosque» (Fig. 2; Tabla 1). El mayor número de especies en «Bosque» está acorde con la mayor complejidad taxonómica y estructural de la vegetación de esta unidad paisajística.
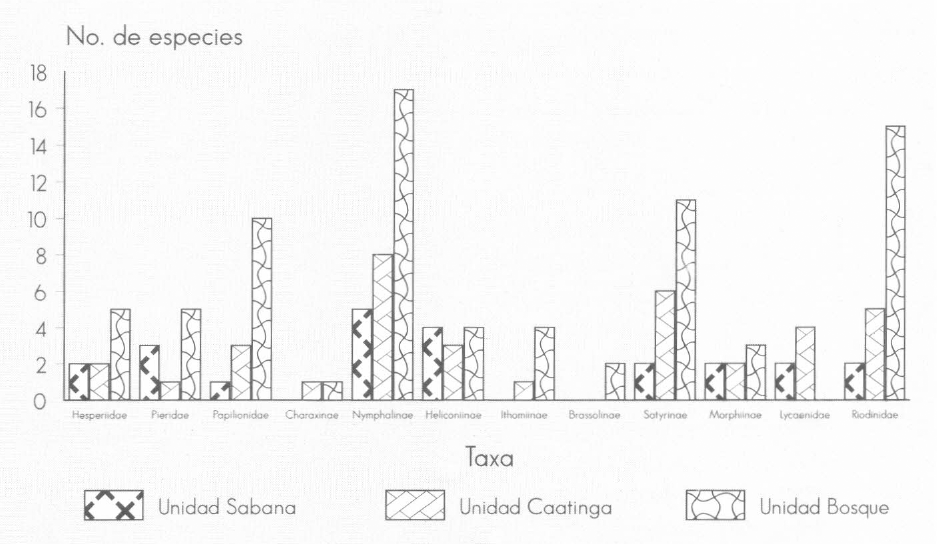
Número de especies colectadas u observadas por familia o subfamilia en cada unidad de vegetación.
Especies colectadas y observadas en la Serranía de Taraira, Vaupés. 1993.

En cuanto a la distribución de especies, todas o la mayoría de las pertenecientes a Hesperiidae, Brassolinae, Satyrinae, Ithomiinae y Riodinidae se observaron o capturaron en una sola unidad de vegetación, mientras que varias de las especies de Papilionidae, Pieridae, Heliconiinae, Nymphalinae y Morphiinae se encontraron en dos; sólo Baeotus amazonicus Riley y Eunica viola Bates (Nymphalinae), Heliconius sara Fabricius (Heliconiinae) y Morpho peleides Kollard (Morphiinae) fueron comunes para las tres unidades paisajísticas (Tabla 1). Cabe destacar que la mayoría de las especies del primer grupo tienen en común una capacidad de vuelo limitada y pertenecen generalmente al estrato herbáceo-arbustivo; por el contrario, las especies del segundo grupo son capaces de desplazarse grandes distancias y permanecen generalmente en el estrato medio o en el dosel arbóreo. No se observan mayores diferencias según los hábitos alimenticios de las especies, ya que en ambos grupos se encuentran tanto especies generalistas como especialistas.
En la Tabla 2 se observa como el número de especies de «Bosque» duplica el de las presentes en «Caatinga» y triplica al de «Sabana». La mayor proporción de especies observadas en una sola unidad paisajística es también mayor en «Bosque» (67% del total de las especies de la unidad), seguido por «Caatinga» (48%), mientras que un 26% de las observadas en «Sabana» fueron vistas sólo en esta unidad. Según los índices de Soremsem y Jaccard, existe una similaridad ligeramente mayor entre «Bosque» y «Caatinga», pero no hay una afinidad significativa (valores superiores a 0,7) entre cualquiera de las tres unidades, siendo cada una lo suficientemente diferente para ser considerada una comunidad independiente (Tabla 3).
Número total de especies colectadas en cada unidad de vegetación, número de especies encontradas en una sola unidad (únicas) y de especies comunes entre unidades.
Indices de Similaridad entre las unidades de vegetación
Valores superiores a 0,7 indican que existe una afinidad significativa entre las muestras comparadas.
Cabe destacar que en cuanto a especies abundantes y comunes sí existe mayor afinidad entre las mariposas de «Bosque» y «Caatinga», que comparten varios elementos como especies de Pierella, Haetera y Euptychia, cuya abundancia varía; algunos elementos comunes en «Bosque» pasan a ser abundantes en «Caatinga», mejor adaptados quizás a la comparativamente menor diversidad taxonómica de sus grupos vegetales, que cuenta con algunos elementos especializados que se ven favorecidos en este ambiente. Por el contrario, la mayoría de las especies abundantes y comunes de «Sabana» son prácticamente propias de esa unidad, como Eumaeus sara Constantino (cuyos registros de colección en «Caatinga» corresponden exclusivamente al ecotono con «Sabana»), Calystrina sp.1 (Lycaenidae), Euptychia hermes Fabricius (Satyrinae) y Anastrus sp.2 (Pyrginae). Eunica viola fue también la única especie abundante en las tres unidades (Tabla 4).
Especies abundantes y comunes de mariposas según la unidad de vegetación.
Abundante: más de 10 observaciones por día. Común: 10 a 5 observaciones por día.
Aunque se registró la presencia de algunos elementos indicadores de fuerte intervención humana como Junonia evarete (Cramer) (Nymphalinae) o Urbanus proteus (L.) (Pyrrhopiginae), éstos se restringieron a una pequeña mancha de «Sabana» de origen antrópico utilizada como pista de aterrizaje. Una especie propia de los márgenes de bosque y del dosel arbóreo, Heliconius sara Fabricius, fue también abundante en todas las áreas de «Caatinga» o «Bosque» donde se evidenciaba alguna intervención humana; no obstante, la zona de estudio puede considerarse como poco alterada.
En resumen, se propone que en la zona de estudio existen tres comunidades de mariposas bien definidas correspondientes a las tres unidades de vegetación estructuralmente diferenciables; aunque existe una mayor afinidad entre especies abundantes y comunes pertenecientes a «Bosque» y «Caatinga».
Artropofauna asociada con el suelo:
La distribución del número de individuos por taxa es comparativamente similar entre «Bosque» y «Caatinga», mostrando diferencias únicamente en cuanto a una mayor representación de Diptera en la segunda unidad (Fig. 3a,); en contraste, «Sabana» tiene a Hymenoptera como segundo orden en número de individuos, debido principalmente al incremento en especímenes de Formicidae (Fig. 4b); existe también una representación comparativamente mayor de Aracneida, Mycrocoryphia, Hemiptera y Diptera con respecto a las otras dos unidades. Collembola registró el mayor número de individuos en las tres unidades paisajísticas (Fig. 3a y 4f).
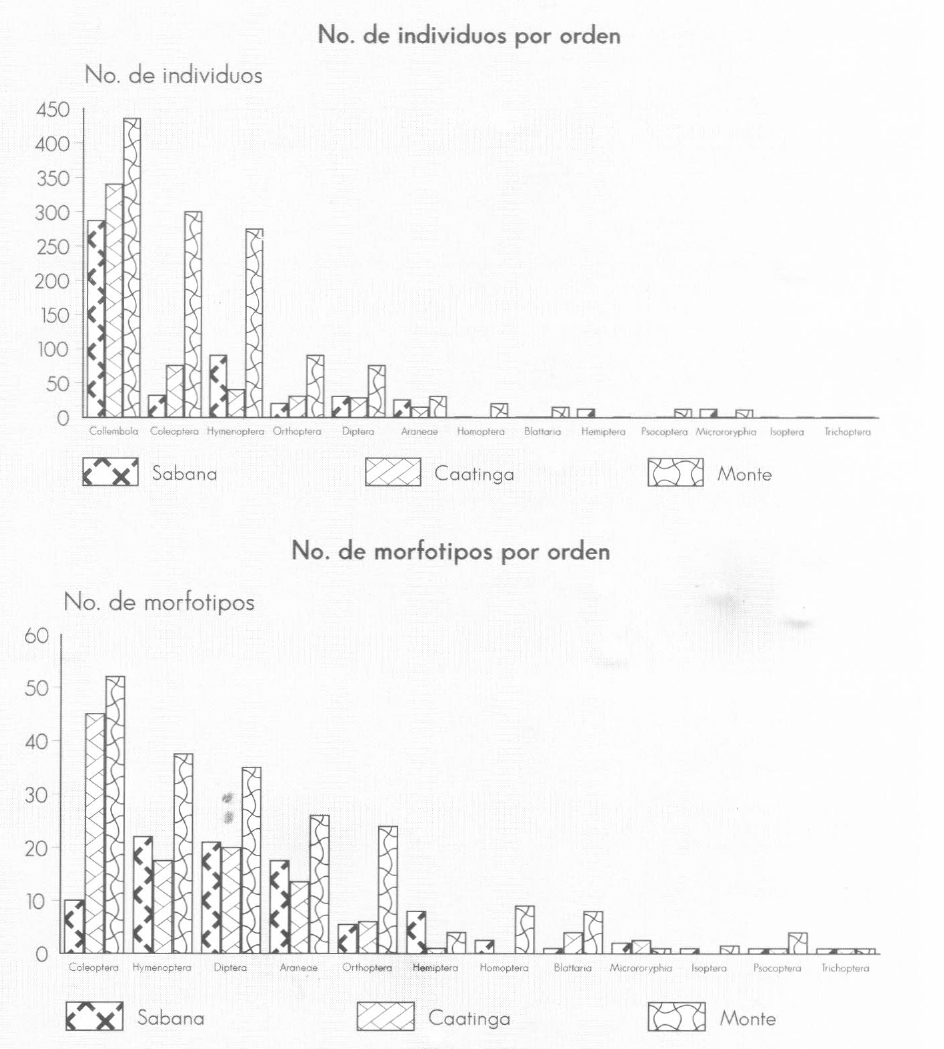
Número de individuos (
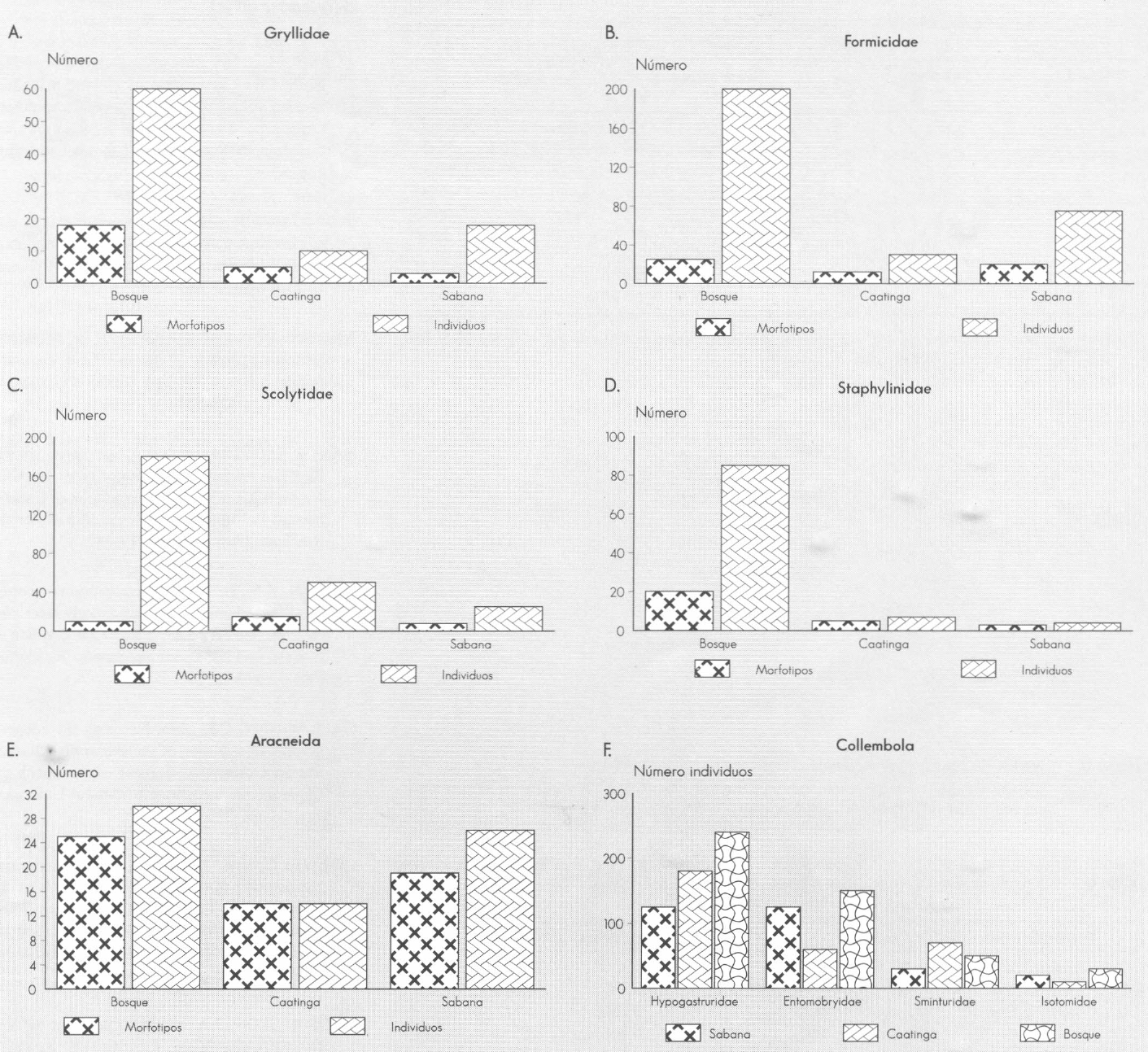
Número de individuos y morfotipos capturados para taxa específicos en la únidades estudiadas.
Las diferencias son más claras al comparar el número de morfoespecies por orden (Fig. 4b); en «Bosque» existen un grupo con alto número morfoespecies (Araneae, Coleoptera, Diptera e Hymenoptera, Orthoptera) y otro con bajo (órdenes restantes) sin que se presente un orden con un número de morfoespeciess comparativamente alto respecto de los restantes; en «Caatinga» Coleoptera es el de mayor número de morfoespecies, aunque la distribución en los otros órdenes es parecida a la de «Bosque» pero con ausencia de Homoptera e Isoptera y mayor presencia de Microcoryphia. «Sabana» presenta un comportamiento totalmente diferente, Hymenoptera es el orden más diverso y Coleoptera es superado por Diptera y Aracneida; Hemiptera, al igual que Microcoryphia, presenta el mayor número de morfoespecies en esta unidad paisajística.
A nivel de taxa se encuentra que Aracneida, Formicidae y Gryllidae presentan el menor número de individuos y morfoespecies en «Caatinga»; Scolytidae y Staphylinidae lo presentan en «Sabana» y es destacable el alto número de morfoespecies de la primera familia en «Caatinga» (Fig. 4c). Un caso especial es el de Collembola, donde la distribución del número de individuos por familia varía según la unidad paisajística; nuevamente se refleja la afinidad entre "Bosque" y «Caatinga» y la diferencia entre cualquiera de estas dos y «Sabana» (Fig. 4f).
En el análisis de similaridad se encontró mayor afinidad entre las subparcelas de «Caatinga» y las de "Bosque" (Tabla 5; Fig. 5); este dendrograma se acopla con los resultados de los párrafos anteriores. Al comparar los valores del índice de diversidad se encuentra que cada una de las subparcelas y la parcela de "Bosque" en conjunto son más diversas que cualquiera de las subparcelas de «Caatinga» o de «Sabana» (Tabla 6); el valor del índice de diversidad fue más homogéneo en las subparcelas de «Caatinga», lo cual puede ser reflejo de una mayor monotonía en la composición de las comunidades. Los valores más bajos del índice de Simpson encontrados en «Caatinga» pueden ser indicativos de una menor dominancia relativa de unos pocos taxa en esta unidad paisajística (Tabla 6).
Matriz de coeficientes de Similitud (según Jaccard) entre las diferentes subparcelas.
Los números corresponden a las tres subparcelas respectivas de la parcela de cada unidad (Sabana: 1, 2, 3; Caatinga: 4, 5, 6; Bosque: 7, 8, 9)
Indices de diversidad, según el número de individuos por taxa.
H: Valores según función de Shannon-Weiber. D: Valor según función de Simpson.
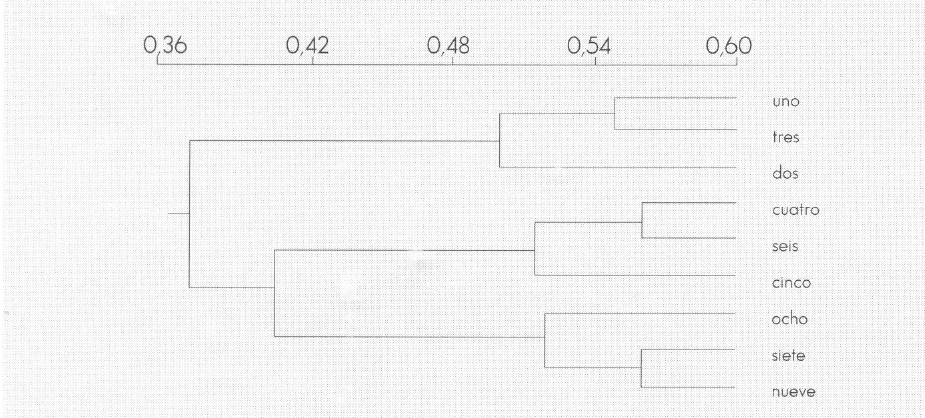
Matriz de coeficientes de similitud (según índice de Jaccard) entre las diferentes subparcelas. Los números corresponden a las tres subparcelas respectivas de la parcela de cada unidad (Sabana: 1, 2, 3; Caatinga: 4, 5, 6; Bosque: 7, 8, 9).
En resumen se tiene nuevamente una comunidad de "Bosque" más diversa y un poco más relacionada con "Caatinga" que con "Sabana", lo cual confirma en buena medida el análisis realizado con mariposas.
Discusión
La correspondencia entre la complejidad estructural de la vegetación y la diversidad de la comunidad de mariposas y de la artropofauna asociada con el suelo, ratifica las propuestas que de esta correlación presentan Anderson y Shugart (1974), Gilbert (1984), Hair (1987), Brown (1991), Kremen (1992) y Dinizt y Kitayama (1994). Las comunidades de «Sabana» y de "Bosque" son completamente diferenciables, mientras que «Caatinga», pese a ser un sistema independiente, podría ser una prolongación de la biota de "Bosque" adaptada quizás a unas condiciones de pobreza de nutrientes más severas, siendo menos diversificada, con mayor dominancia relativa de sus taxa y con pocos elementos propios; la menor variación en los índices de diversidad obtenidos de sus subparcelas apoyan esta hipótesis.
Vale la pena destacar que según los parámetros dados por Brown (1991), es preferible dar mayor peso al análisis de las especies abundantes debido a su muy probable mayor permanencia e importancia ecológica en el hábitat. Si así se hace, las mariposas reflejan también una aproximación incluso mejor que lo obtenido en artropofauna, ya que se hace evidente la relación entre «Caatinga» y «Bosque».
Una causa de la baja diversidad registrada en mariposas y la artropofauna de «Sabana», adicional a la relación con una menor complejidad estructural, podría ser el área pequeña y fraccionada que presenta esta unidad, distribuida a manera de islas en medio de la vegetación de "Caatinga Alta" y "Bosque"; lo que también explicaría el alto número de elementos propios de dosel de bosque amazónico aquí observado. Un aspecto interesante de la entomofauna de «Sabana» es la alta dominancia y diversidad de Formicidae, lo que puede estar relacionado con el alto número de plantas mirmecófilas observadas en la unidad.
Un aspecto interesante es el bajo número de registros de observación, lo cual puede ser consecuencia de la época del muestreo, coincidente con la fase de lluvias; no obstante, también puede estar asociado con las características de la vegetación: bosques que crecen sobre suelos arenosos extremadamente pobres y sometidos a lavados permanentes, o que surgen prácticamente de la roca madre (Huguett et al. 1979). Esto puede generar un sistema con muy bajo número de especies, pero también con alta diversidad; el bosque se convierte en un mosaico fisionómicamente homogéneo pero muy variable a nivel de especies, donde la composición de una hectárea puede ser muy diferente a la de las circundantes. En favor de esto se puede decir que inventarios previos cercanos a la zona de estudio (Estación Biológica de Caparú, Laguna de Taraira, que tiene vegetación de "Igapo" y Bosque pluvial amazónico de suelos arenosos principalmente; (Deffler 1993; datos no publicados); muestran una fauna de mariposas con varios elementos en común pero también con numerosas especies que nunca se observaron, y viceversa.
Se considera valioso mencionar que la mayoría de las especies vegetales de «Sabana» permanecen florecidas gran parte del año (según pobladores de la región); de las especies observadas se encontró que en su mayoría abren una o unas pocas flores por individuo y por día; este comportamiento, que generalmente obliga al polinizador a visitar diferentes plantas para completar sus necesidades energéticas, permite un mayor cruzamiento entre las plantas polinizadas (Baker 1983); ésta podría ser una estrategia importante para asegurar un flujo constante de polinizadores en una comunidad de vegetación que está tan fraccionada y aislada. Es necesario realizar posteriores estudios acerca de estos aspectos para tener algún conocimiento de este tipo de vegetación de la Amazonia.
Conclusiones
Se tiene que la coincidencia entre lo obtenido de manera cualitativa y con muy pocos ejemplares en mariposas y el trabajo más detallado y laborioso con la artropofauna del suelo, confirma, al menos de manera parcial, la efectividad de las mariposas como bioindicadores del tipo de hábitat, respaldando los trabajos de Brown (1991), Kremen (1993) y New (1993); y hacen evidente ventajas tales como su facilidad de observación en campo, siendo muchas especies identificables en pleno vuelo, y su manejo, quizás de los más sencillos. Se ratifica entonces su condición de excelentes bioindicadores y, al menos para el presente estudio, su alto grado de confiabilidad.
Footnotes
Agradecimientos
Quiero agradecer a todas las personas que integraron la expedición «Amazonas-93» por su constante, desinteresada y excelente colaboración; especialmente Tigga Kefersteine, al profesor Alberto Cadena, y a nuestro guía Elías Macuje. A Rocío Cortez, Javier Rodríguez y Sara y Thomas Deffler, por la información acerca de la vegetación. Al profesor Gonzalo Andrade y los biólogos Carlos Sarmiento, Yaneth Muñoz y Angela Amarillo por sus valiosas críticas y correcciones. A Adriana Hurtado y Jean F. Le Crom por la bibliografía utilizada en la determinación. A Sparta por todo.