
Abstract
In this work the community structure of trichoptera and its spatial distribution along an altitudinal gradient at the Carrizal stream located on the Iguaque Santuary (Boyacá, Colombia) was studied. Monthly determina- tions of physical and chemical parameters were registred from september 1994 to october 1995 in order to establish the biotic and abiotic relationships. The community con- sisted of 15 genera belonging to nine fami- lies. The most important genera of caddisflies found were Ochrotrichia (28%) and Contulma (17.8%), Helycopsiche (13.4%), and Morto-niella (9.49%). The physical-chemical and biological characteristics of the Carrizal stream is dictated by the rainfall pattern. This lotic ecosystem can be considered as oligo- trophic, due to limited levels of nitrogen and phosphorus, low conductivity and quantity of suspended solids.
Introducción
Trichoptera es un orden de insectos am- pliamente distribuido en el mundo a ex- cepción de la región antártica y ocupa una gran variedad de hábitats de agua dulce, lóticos y lénticos. La mayoría de los gru- pos prefieren corrientes limpias y oxige- nadas, alcanzando la mayor diversidad en los sistemas lóticos fríos, aunque algunas especies viven en sistemas lénticos y re- mansos de ríos y quebradas (Wiggins 1977).
Los tricópteros son muy importantes por su utilización como bioindicadores, te- niendo en cuenta el estrecho rango de con- diciones ecológicas toleradas por muchas de sus especies, así como por su papel en la transferencia de energía a través de los niveles tróficos de la mayoría de los sis- temas acuáticos. Estas características han hecho que este grupo sea considerado un elemento fundamental en las investiga- ciones tanto básicas como aplicadas so- bre la ecología de los cuerpos de agua.
Los tricópteros son insectos holometábo- los y, a excepción de algunas especies, todos sus estados inmaduros son acuáti- cos. Generalmente pasan por cinco insta- res larvales, luego de los cuales en todas las especies, incluyendo las de vida libre, construyen un estuche que se fija a un sustrato para pasar al estado de pupa. Es- tos estuches presentan formas variadas y son construidos con diversos materiales: granos de arena, restos vegetales, peda- zos de hojas y secreciones de seda.
Las familias y géneros de Trichoptera han sido bien estudiados en las zonas templa- das y en América Central, mediante cla- ves e ilustraciones, entre las que se pue- den citar las de Flint (1967), Hickin (1967), Wiggins (1977, 1984), Pennak (1978), Botosaneanu y Flint (1982), Mor- se y Holzenthal (1984), Merrit y Cum- mins (1984) y Hilsenhoff (1991).
Hulbert et al. (1981) mencionan que des- de la época de Burmeister, en 1839, se han descrito tricópteros de la región tro- pical. Sin embargo, la única lista regio- nal de Trichoptera fue publicada por Ulmer en 1913, citado por Roldán (1988), y contiene 162 especies. Los tricópteros neotropicales han sido estudiados por Flint (1963, 1966, 1967, 1972, 1974a y b, 1975, 1978, 1980) y este autor reporta, en 1981, 1.100 especies al sur de los Esta- dos Unidos de América, de los cuales 425 han sido señalados de Sur América con exclusión de Argentina, Chile y Uruguay. A pesar de este número aparentemente grande, Flint (1981) estima que sólo la tercera parte de los tricópteros de la sub- región brasilera ha sido descrita. La defi- ciencia en el conocimiento de este grupo se debe en primer lugar, al gran número de especies aún no descritas y por otro lado, a la falta de investigación básica que reconozca, documente y dé a conocer esta biodiversidad.
La tricopterofauna colombiana ha sido poco estudiada, lo que no permite esta- blecer un cálculo real de sus especies. Estudios básicos sobre taxonomía y bio- ecología de los Trichoptera han sido rea- lizados por Correa et al. (1981), Roldán (1988), Flint (1991) y Quintero y Rojas (1987). Otros trabajos importantes para Colombia son los de La Rotta (1989), Rincón (1995) y Holzenthal y Flint (1995). Con base en estos trabajos se pue- de establecer hasta el momento un total de 136 especies, distribuidas en 42 géne- ros y 14 familias.
La presente investigación tuvo como ob- jetivo fundamental conocer las caracte- rísticas estructurales y las variaciones es- paciales y temporales que presenta la co- munidad de tricópteros de la quebrada Carrizal y determinar de qué manera las características químicas y físicas del me- dio influyen en ésta.
Igualmente se pretende contribuir al es- tablecimiento de un sistema de bioindi- cación regional, basado en los macroin- vertebrados acuáticos.
Este trabajo está enmarcado dentro del proyecto de investigación «Biodiversidad y Conservación de los Sistemas Acuáti- cos del Santuario de Fauna y Flora de Iguaque (Boyacá)», financiado por la Universidad Pedagógica Nacional y el Fondo para la Protección del Medio Am- biente "José Celestino Mutis" FEN CO- LOMBIA.
Materiales y Métodos
El presente trabajo se realizó en la que- brada Carrizal, ubicada en el Santuario de Fauna y Flora de Iguaque. Esta reser- va se encuentra localizada en jurisdicción de los municipios de Villa de Leyva, Arcabuco y Chíquiza, en el departamen- to de Boyacá, sobre la cordillera Orien- tal, entre 5° 36' 02" y 5° 44' 38" de latitud norte y 73° 22' 57" y 73° 31' 20" de lon- gitud oeste (Garcés y De La Zerda 1994). Tiene una superficie de 6.750 hectáreas y alturas que oscilan entre 2.400 y 3.600 msnm (Fig. 1).
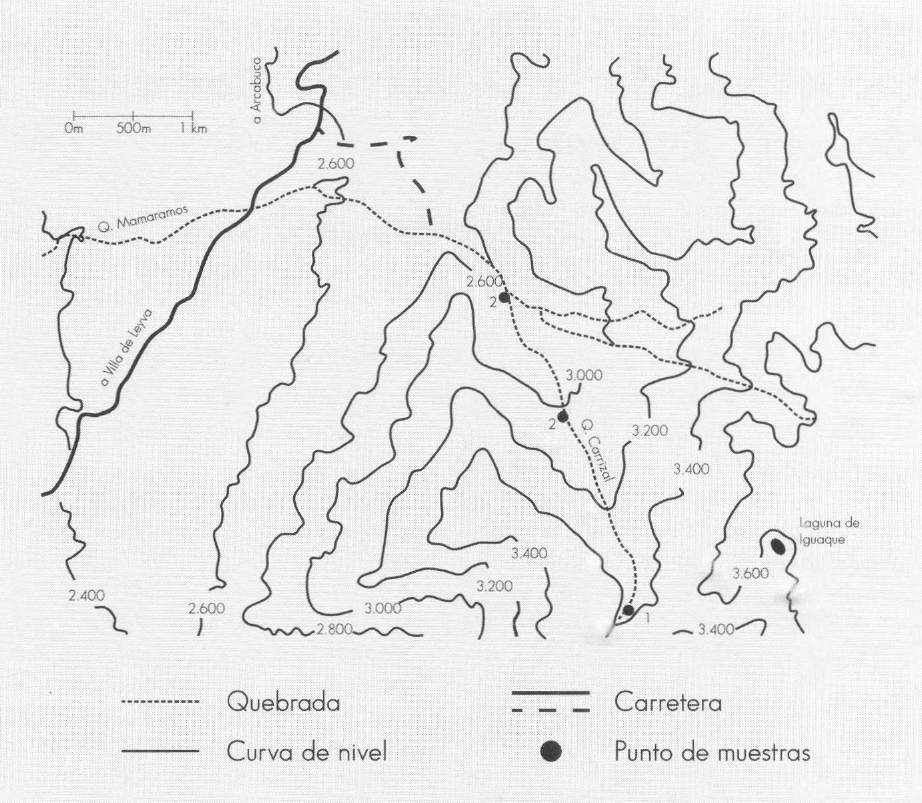
Localización de la zona de estudio. Quebrada Carrizal (Santuario de Fauna y Flora de Iguaque, Boyacá)
Entre septiembre de 1994 y octubre de 1995 se realizaron diez muestreos. La selección de las estaciones de muestreo se hizo con base en mediciones del oxí- geno disuelto, la conductividad y la com- posición de la comunidad de tricópteros: en diferentes coriotopos. Acorde con las mediciones mencionadas se establecieron tres estaciones de muestreo en un gradien-. te altitudinal. La primera Estación se ubi- có hacia el nacimiento de la quebrada a 3.350 msnm, zona de vida bosque muy húmedo montano (bmh- M), que presen- ta vegetación con formas de crecimiento arbustivo, herbáceo y mucinal, y formas de vida caulirrósula (frailejones) y gra- minoide (pajonales). La segunda y terce- ra Estaciones se ubicaron a 3.050 y a 2.780 msnm, correspondientes al límite superior e inferior de la zona de vida bos- que muy húmedo montano bajo (bmh- MB). Esta zona presenta flora heterogé nea con pino colombiano, amarillo y arra- yán (Barrera y Acosta 1995).
En cada uno de los sitios se tomaron muestras de las formas inmaduras y de adultos. Los estados inmaduros se colec- taron de ocho coriotopos así: Hojarasca (H), Gravilla en corriente lenta (GCL), Gravilla en corriente rápida (GCR), Mus- go sobre piedras en corriente lenta (MCL), Musgo sobre piedras en corrien- te rápida (MCR), Piedras en corriente len- ta (PCL), Piedras en corriente rápida: (PCR) y Ribera o vegetación anfibia (R).
En cada una de las estaciones, las mues- tras se tomaron en una área aproximada de 10 mβ, empleando un tiempo aproxi- mado de cinco minutos en cada coriotopo. Los estados inmaduros se colectaron uti- lizando redes de Thienneman. Paralela- mente, en los dos primeros muestreos se utilizaron métodos convencionales (Sur-ber, colecta directa, red triangular y de mano). Los especímenes se colocaron ini- cialmente en frascos con alcohol etílico al 50% y posteriormente se limpiaron y separaron en alcohol al 70%.
La metodología empleada en esta inves- tigación (muestreo en coriotopos) fue pro- puesta inicialmente por Braukmamm (1987) e implementada por primera vez para Colombia en este trabajo por suge- rencia de Riss (1994). Esta metodología, a diferencia de las métodos convencio- nales que se han venido desarrollando para el estudio de los invertebrados acuá- ticos, tiene en cuenta la heterogeneidad espacial presente en un sistema lótico y permite caracterizar la distribución hori- zontal y vertical de los macroinvertebra- dos acuáticos en cualquier tipo de sustrato (grava, piedras, rocas, macrofitas, etc). Estos aspectos son esenciales para el co- nocimiento de la autoecología de los di- ferentes grupos y permitirán, en el futu- ro, establecer sistemas de bioindicación regionales apropiados para la caracteri- zación de los recursos acuáticos.
Para el muestreo de adultos se usaron trampas de luz y se revisó la vegetación aledaña a la cuenca, teniendo en cuenta que los adultos son principalmente noc- turnos y que durante el día reposan en hábitats oscuros y fríos cerca a los cuer- pos de agua. La captura de los adultos se realizó en las orillas de la quebrada, utili- zando como trampas de luz, lámparas Coleman de luz blanca y tubos fluores- centes conectados a una batería de 12 watios y se colocó una tela de algodón blanca de aproximadamente un metro cuadrado. Los muestreos se efectuaron entre las 18 y las 22 horas, cuando los adultos son más activos y son atraidos por las fuentes de luz.
Igualmente se analizó el comportamien- to temporal de 30 parámetros físico-quí- micos. En el campo se tomaron datos de temperatura del agua, conductividad y oxígeno disuelto (ppm) con un oxímetro YSI modelo S54, previamente calibrado a cada altitud. La determinación de los de- más variables fisicoquímicas: color, al- calidad (total, hidróxidos, carbonatos y bicarbonatos), acidez (total, mineral y sales hidrolizadas), pH, CO₂, dureza (to- tal, carbonácea y no carbonácea), calcio, magnesio, hierro, manganeso, sodio, clo- ruros, nitratos, nitritos, amonio, sulfatos, ortofosfatos, fósforo total, turbidez y só-: lidos disueltos totales se realizó dentro de las siguientes 24 horas a su recolección, en el Laboratorio de Aguas del Departa- mento de Ingeniería Ambiental de la Uni- versidad Nacional, siguiendo los méto- dos recomendados por APHA-AWWA- WPCF (1989).
La identificación de las familias y géne- ros de Trichoptera presentes en la quebra- da se realizó con base en las claves de Wiggins (1977), Flint (1967, 1977, 1983, 1991), Botosaneanu y Aint (1982), Rol- dán (1988) y Morse y Holzenthal (1984). Las determinaciones fueron corroboradas por el doctor Oliver Flint, del Smithsonian Institution, en Washington.
El análisis e interpretación de los resulta- dos físicos, químicos y biológicos se hizo con base en técnicas de clasificación y or- denamiento. Para el análisis de los pará- metros físico-químicos de la quebrada Ca- rrizal, previa transformación de los da- tos, se aplicó un modelo estadístico de ordenamiento con base en una matriz de correlación, que analiza la matriz secun- daria por filas y por columnas; esto es a nivel de sitios y de variables (Biplot o Análisis de componentes principales).
Para la clasificación de los datos biológi- cos, la matriz de datos se transformó en una matriz de similitud, mediante la apli- cación del Indice de Bray & Curtis, pre- via transformación logarítmica de las abundancias de los tricópteros. Este índi- ce se calculó tanto en modo Q (entre co- riotopos) como en modo R (entre géne- ros). A partir de la matriz de disimilitud se elaboró un dendrograma, utilizando para ello el subprograma Cluster, del pa- quete estadístico Statistical Ecology.
Con respecto a la ordenación de las va- riables biológicas se utilizó un Análisis Multidimensional no Paramétrico (NM- DS), para lo cual se emplearon los resul- tados de la matriz de similitud (modo Q), aplicando el coeficiente de asociación de Bray Curtis y los resultados del análisis de componentes principales (PCA) entre coriotopos. Además se calcularon los ín- dices de diversidad de Shannon-Weinner, Riqueza de Margalef y Equitatividad de Pielou.
Resultados y Discusión
Aspectos Físico-Químicos
Desde el punto de vista climático, en el Santuario de Iguaque se presenta un ré- gimen de precipitación bimodal: el pri- mer pico en los meses de abril-mayo y el segundo de octubre a noviembre (Garcés. y De La Zerda 1994). De manera similar se presenta la distribución de los cauda- les, con dos períodos de aguas altas entre octubre- noviembre y abril-mayo y dos períodos de aguas bajas entre diciembre- marzo y junio-agosto.
En las tres estaciones de muestreo, el comportamiento de la mayoría de las va- riables físico-químicas a lo largo del año es uniforme; sin embargo, en la época de mayor precipitación (septiembre-noviem- bre) y en menor intensidad en abril y mayo, aumentan los valores de turbidez, color y amonio (Fig. 2). Como es de es- perar, los valores de turbidez muestran una notable relación con la precipitación e indican el aporte de material alóctono en suspensión, procedente de los sistemas aledaños a la cuenca.
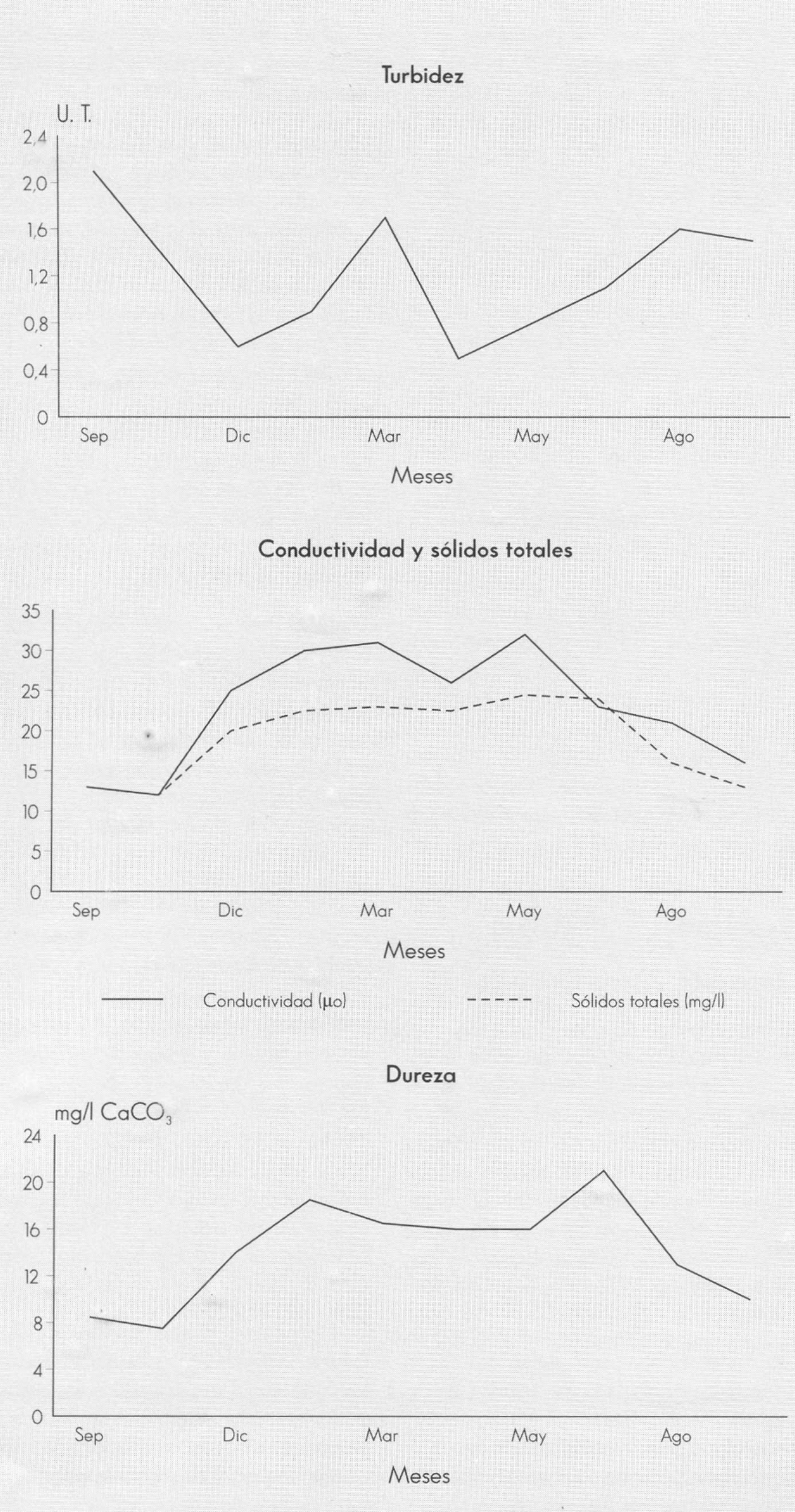
Caracterización físico-química de la qebrada Carrizal. Santuario de Iguaque, Boyacá. 1994-1995.
La conductividad, los sólidos totales, el calcio, el magnesio, la dureza total y la alcalinidad muestran un comportamien- to opuesto, pues disminuyen a sus nive- les más bajos como resultado del efecto de dilución, debido al aumerito de los cau- dales de la quebrada en esta época (Fig. 3); en estos meses, los valores de casi to- dos los parámetros son similares en todo el sistema. La conductividad no sobrepa- sa en ningún caso los 32 S/cm y la alcali- nidad presenta valores muy bajos. Esto es debido a que las cuencas de drenaje están constituidas, casi en su totalidad, por areniscas de color blanco amarillento del cretáceo (Molano 1990).
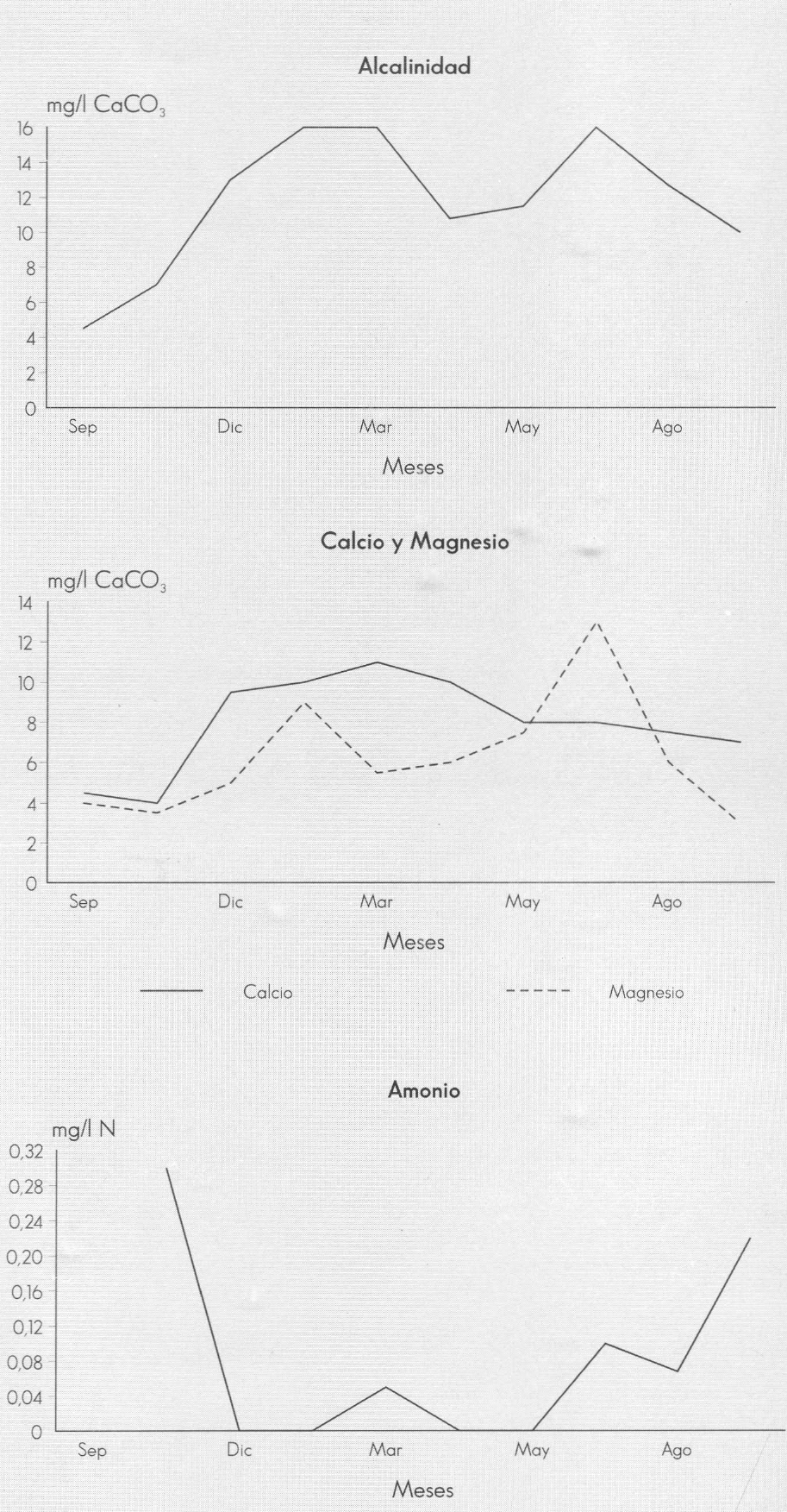
Caracterización físico-química de la quebrada Carrizal. Santuario de Iguaque, Boyacá. 1994-1995.
Tanto la dureza como la conductividad están definidas especialmente por las va- riaciones en las concentraciones de cal- cio y magnesio. En la quebrada Carrizal, los valores de conductividad siguen las variaciones en las concentraciones de cal- cio (Wetzel 1981).
El régimen pluviométrico parece ser el factor más importante para determinar el comportamiento de las variables físico- químicas. Lo anterior se puede corrobo- rar en el biplot de componentes principa- les (Fig. 4), en el cual se agrupan los ele- mentos, como: color, nitritos, turbidez, amonio, acidez y CO₂ con los meses o épocas más lluviosas (septiembre, octu- bre y noviembre) (Grupo I); alcalinidad, calcio, conductividad y sólidos totales con los meses de diciembre, marzo y mayo, y los elementos dureza y magnesio con las épocas de febrero, junio y diciembre, ra- tificando que el comportamiento físico y químico depende de las épocas de alta y baja precipitación.
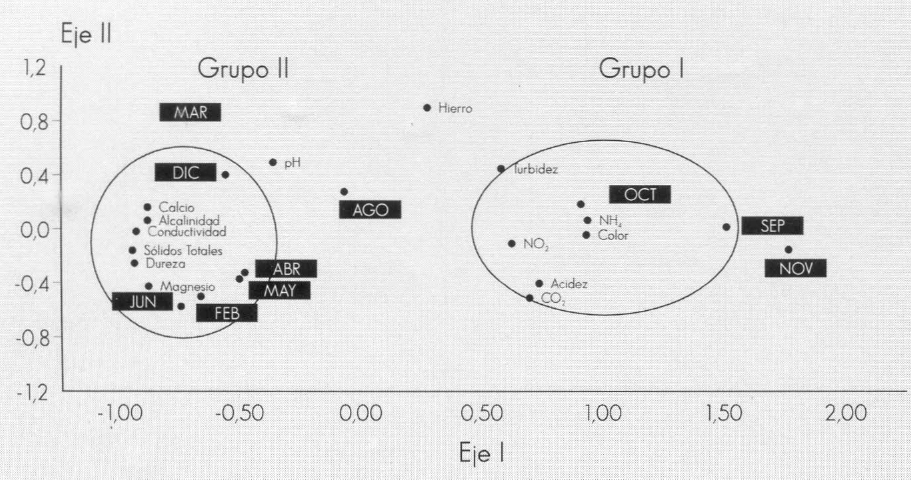
Análisis de componentes principales. Parámetros físico-químicos.
La mayoría de los ríos tropicales tienen un ciclo anual modificado por el patrón de lluvias, esta influencia es más marca- da cuando el tamaño de la cuenca es pe- queño (Payne 1986). Esto se pudo cons- tatar en la Estación 1 a 3.350 msnm (na- cimiento de la quebrada), donde las va- riaciones en los parámetros físico-quími- cos fueron mayores.
La quebrada Carrizal puede catalogarse como un sistema acuático de aguas cla- ras, blandas, ligeramente ácidas y sobre- saturadas de oxígeno (7,7 mg/l), hecho que se ve favorecido por la topografía quebrada de la región y las bajas tempe- raturas del agua (9-11°C). Aunque no se presentan fluctuaciones marcadas para este factor, en los meses de mayor preci- pitación se presenta una ligera variación, posiblemente como consecuencia de los procesos de oxidación del material orgá- nico proveniente de los sistemas aleda- ños. Las características mencionadas an- teriormente son típicas de los sistemas acuáticos de alta montaña tropical (Do- nato 1991).
Aspectos biológicos
En los seis muestreos en los coriotopos se encontró un total de 5.210 individuos pertenecientes a 15 géneros y a nueve fa- milias de Trichoptera. Por su abundancia relativa se destacan los géneros Ochro-trichia (28,3%), Contulma (17,8%), Helicopsyche (13,4%), Mortoniella (9,49) Neotrichia (8,1%), Grumichella (5,2%) y Alisotrichia (4,7%).
La composición de la comunidad de tri- cópteros fue bastante homogénea en las tres estaciones de muestreo; sin embar- go, se presentó una alta diferencia en cuanto a la abundancia de individuos de cada género. En la Estación 1, los géne- ros más importantes fueron: Mortoniella, Helycopsiche y Smicridea; en las Esta- ciones 2 y 3 abundaron los géneros Ochrotrichia, Contulma, Helycopsiche, Neotrichia y Alisotrichia (Fig. 5).
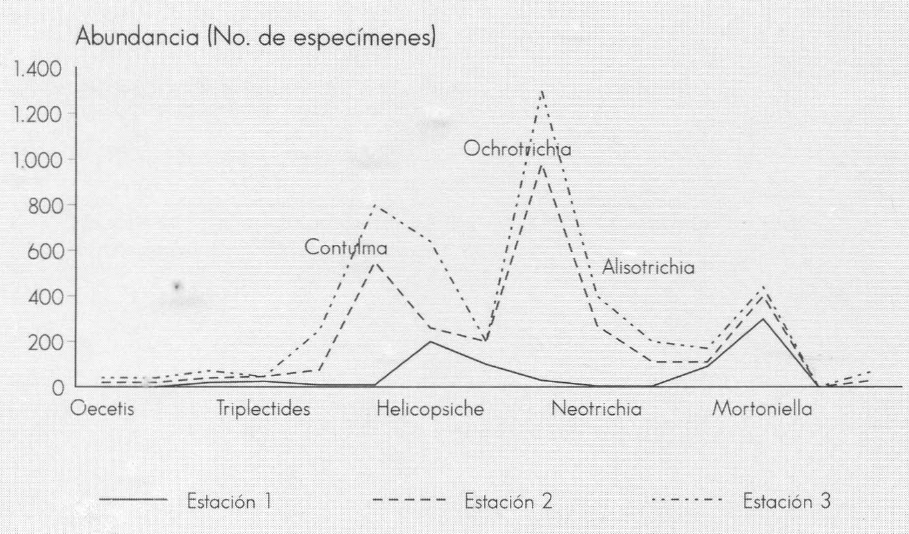
Abundancia de tricópteros en tres estaciones de la quebrada Carrizal. Santuario de Iguaque (Boyacá).
A diferencia del muestreo por coriotopos, con los métodos tradicionales (Surber, colecta directa, red triangular y de mano) cambia la estructura de la comunidad de tricópteros, pues se sobreestima la impor- tancia de géneros como Helycopsiche y Grumichella, y se subestima la importan- cia de otros como Ochrotrichia, Neotrichia y Alisotrichia, los cuales están muy bien representados en el Neotrópico (Fig. 6).
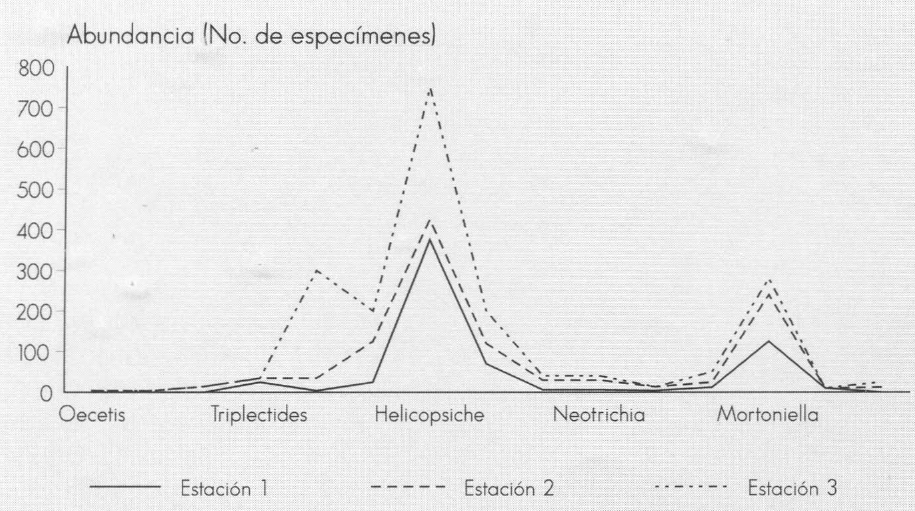
Abundancia de los tricópteros de la quebrada Carrizal. Métodos: Suber, coleta directa, red triangular).
Con respecto a la distribución espacial, el coriotopo Musgo corriente rápida (MCR) alberga el 33,8% de la tricóptero- fauna total; el Piedra corriente rápida (PCR) el 17,9%, el Ribera (R) y Musgo corriente lenta (MCL) el 12,4 y 8,12%, respectivamente. (Fig. 7). El menor por- centaje se encontró en los coriotopos GCR y GCL, sustratos que presentan alta inestabilidad, especialmente para aquellos grupos que construyen redes, como es el caso de Smicridea (Hydropsichidae), cu- yas formas inmaduras necesitan una base firme para una vida sedentaria (Townsend 1980), este género, por el contrario, fue muy abundante en los cariotopos MCR y PCR.
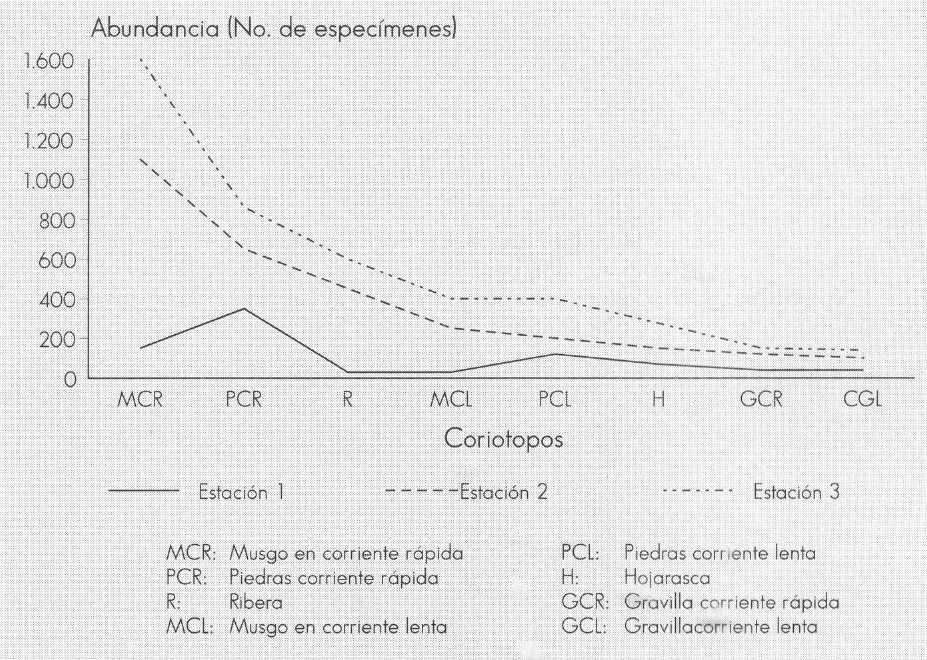
Abundancia de los tricópteros en diferentes coriotopos de la quebrada Carrizal. Santuario de Iguaque (Boyacá).
Los resultados obtenidos a partir del Aná- lisis Multidimensional no Paramétrico (NMDS) entre muestras de tricópteros, permiten evidenciar la heterogeneidad espacial en un sistema lótico, como la quebrada Carrizal. Como se puede apre- ciar, en la Figura 8 se evidencian cuatro grupos. El Grupo I conformado por los coriotopos MCR, MCL y R y constituido por los géneros dominantes Contulma y Ochrotrichia, los cuales son principal- mente neotropicales (Wiggins 1977) y presentaron una similaridad del 64%. En este grupo también se incluyen géneros codominantes como: Neotrichia, Grumichella, Alisotrichia y Smicridea, con por- centajes de similaridad conjunta del 66% (Fig. 9). Los coriotopos MCL y R presen- taron las diversidades más altas (1,95 y 1,92 bits), presentando igualmente una alta uniformidad (0,74 y 0,71).
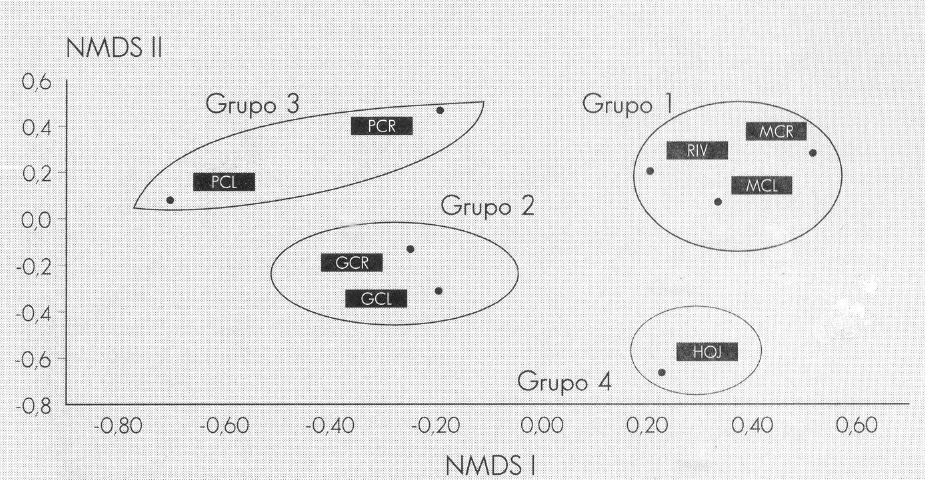
Ordenamiento de coriotopos en la quebrada Carrizal, Santuario de Iguaque, Boyacá. Análisis Multidimensional no Paramétrico (NMDS).
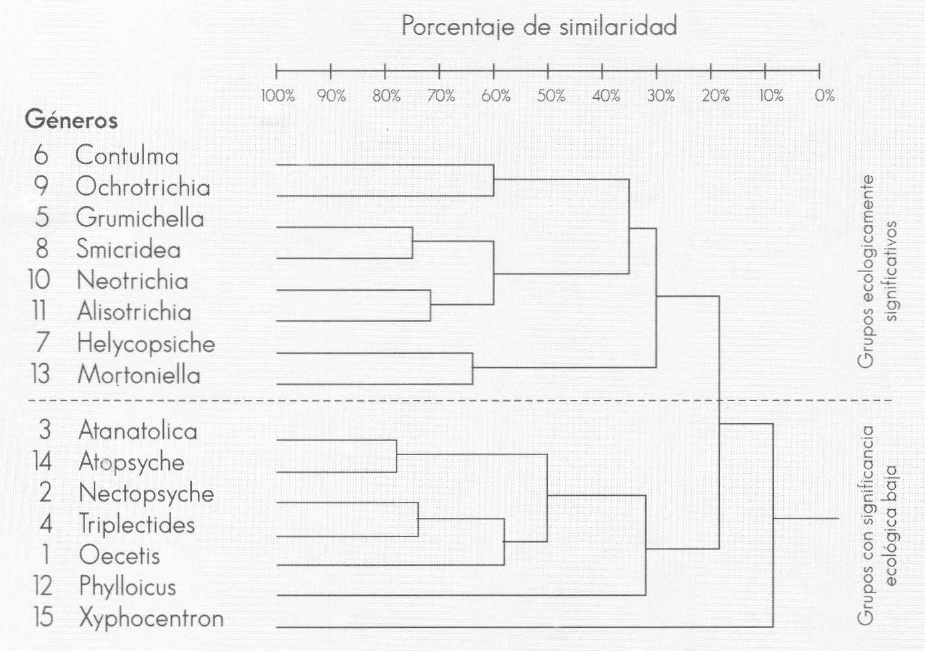
Dendograma de asociación entre géneros de tricópteros de la quebrada Carrizal, Santuario de Iguaque (Boyacá) (Coeficiente de Bray-Curtis).
Según Payne (1986) y Townsend (1980), en las áreas litorales y en la vegetación marginal la diversidad y la abundancia de los diferentes grupos se incrementa dra- máticamente. En este coriotopo se loca- lizó un gran número de larvas, ninfas y adultos de diferentes grupos de insectos.
Los coriotopos GCL y GCR, constituyen el Grupo II, y en este grupo sobresalen por su abundancia alta en todos los mues- treos los géneros Cotulma y Helicopsyche, con bajos valores de diversidad (1,62 y 1,99 bits) y valores de uniformi- dad de 0,63 y 0,77, respectivamente.
El Grupo III incluye los coriotopos PCL Y PCR, los cuales presentaron las diver- sidades más bajas (1,25 y 1,77 bits) y valores de uniformidad de 0,5 y 0,69, res- pectivamente, evidenciados por la domi- nancia de los géneros Mortoniella y Helicopsyche. El Grupo IV corresponde al coriotopo H, integrado por los géneros Nectopsyche, Triplectides, Phylloicus, Atanatolica y Atopsyche que constituyen los grupos más abundantes. Estos géne- ros se consideran de significancia ecoló- gica baja, debido a que se encontraron en pocos coriotopos y sólo en algunos mues- treos. En relación con otros coriotopos, aquí no hay una dominancia tan marca- da; todos los géneros son igualmente im- portantes, y esto se refleja en los altos valores de diversidad (1,93) y de unifor- midad (0,74).
El género Xyphocentron presentó una dis- tribución espacial muy restringida y se en- contró ocasionalmente en el coriotopo R y en una abundancia relativa muy baja. Como se puede apreciar en el dendrogra- ma (Fig. 9), la similaridad conjunta fue muy baja (9%), lo que indica que cada uno de los grupos tienen requerimientos de hábitat específicos.
Los macroinvertebrados acuáticos tienen gran importancia en los estudios de im- pacto ambiental, donde constituyen una variable fundamental para la evaluación de la calidad del agua de un sistema acuá- tico. Con los métodos tradicionales se enfatiza la importancia de los muestreos en rocas y gravilla; sin embargo, este tra- bajo permite enfatizar la importancia de tomar muestras en coriotopos donde se encuentra la diversidad más alta de los sistemas acuáticos.
Con respecto a la distribución altitudinal de los tricópteros, la mayor diversidad (2,04 bits) fue para la Estación 3 ubicada a 2.780 msnm, donde se encontraron 14 géneros. De la Rotta (1989) reporta valo- res altos en esta misma altitud. El menor valor de diversidad correspondió a la Es- tación 1. Así mismo, las abundancias más bajas fueron para esta estación. Allí, la corriente ejerce su mayor influencia so- bre la comunidad béntica, el agua es tur- bulenta y superficial, y ejerce un efecto abrasivo sobre el lecho de la quebrada, por lo tanto sólo los organismos que po- seen las adaptaciones más obvias resis- ten a ser arrastrados y se encuentran en estos sitios (Townsend 1980).Los géne- ros más abundantes fueron Helycopsiche y Mortoniella, los cuales se encontraron adheridos a sustratos pedregosos. Estos géneros son euritolerantes y presentan una alta distribución en el neotrópico (Wig- gins 1977).
Conclusiones
La quebrada Carrizal puede catalogar- se como un sistema acuático de aguas claras, blandas, sobresaturadas de oxí- geno, ligeramente ácidas y con una ba- ja mineralización El régimen pluviométrico parece ser el factor más importante para deter- minar el comportamiento de las va- riables físico-químicas y biológicas de la quebrada Carrizal. En la época de alta precipitación se nota una consi- derable reducción en las abundancias de los grupos de Trichoptera; sin em- bargo, el número de géneros perma- nece constante. La metodología propuesta en este tra- bajo, utilizando coriotopos a diferen- cia de los métodos convencionales empleados para el muestreo de los macroinvertebrados acuáticos, tiene en cuenta la heterogeneidad espacial presente en un sistema lótico y per- mite caracterizar su distribución ho- rizontal y vertical.
Footnotes
Agradecimientos
La autora desea expresar sus agradeci- mientos al Doctor Hans Wolfgang Riss por sus valiosas sugerencias metodológi- cas, al doctor Oliver Flint por la determi- nación de los ejemplares y el suministro de bibliografía. Al Fondo para la Protec- ción del Medio Ambiente FEN COLOM- BIA por la financiación otorgada y al Departamento de Biología de La Univer- sidad Pedagógica Nacional, en especial a los estudiantes María Teresa Páez, Nelson Pineda y Martha Liliana Uribe y a los profesores Carlos Arturo Sierra y Yolanda Ladino por su constante apoyo.