
Abstract
Tagosodes orizicolus (Muir) is an important pest of rice in Latin America because it transmits the Hoja Blanca virus and causes mechanical damage to the plant through its feeding and oviposition. In Colombia, all rice varieties developed through the ICA-CIAT-FEDEARROZ Agreement are reported as resistant to «sogata», common name of the insect. However no efforts to diversify the resistance sources have been made, and commercial varieties and advanced lines are being used as donors of resistance. Given the narrow genetic basis of the resistant sources, this study was undertaken to identify the resistant parents or donors more frequently used in Latin America, and to establish the heritability mode of Makalioka and Mudgo. The materials were characterized as resistant or susceptible based on free-choice and no-choice tests and, on the survival and oviposition of the insect. Two groups resulted: Mudgo, Amistad 82, IRAT 120, IRAT 124 and Makalioka as resistant materials; and Chianan 8, Colombia 1, Bluebonnet 50, IR 8 (ICA), IR 8 (IRRI), Tetep and Cica 8 as susceptible materials. It was found that the damage caused by the insect to the materials was associated with insect survival, oviposition and egg hatching. To determine heritability, two parents were selected: Makalioka and Mudgo, which were crossed with IR 8. F₁ and F₃ populations were also evaluated based on the free-choice test with alternative. Finally, based on the crosses' reactions to the insect damage and comparing them with the parents, a genetic model of resistance is proposed for Mudgo and Makalioka to be a single dominant gene of the homozygous form (AA) accompanied by a modifier gene which interferes in a greater or lesser intensity with the resistance expression (bb>Bb>BB) depending on the material and on the exposure time of the plant to the insect. For Mudgo the recessive homozygous form (bb) which accelerates the expression of the susceptible phenotype and for Makalioka the homozygous dominant form (BB) which delays the expression of the susceptible phenotype.
Introducción
El saltahojas Tagosodes orizicolus (Muir) (Homoptera: Delphacidae), clasificado anteriormente como Sogatodes orizicola, es conocido por los arroceros como sogata (Asche y Wilson 1990). Este insecto, en sus estados de adulto y ninfa, se alimenta de la planta de arroz, causando un daño mecánico; además es transmisor único del virus de la Hoja Blanca (VHB) (Gálvez y Jennings 1959). Las medidas recomendadas para el control de este insecto incluyen el uso de prácticas adecuadas de manejo, uso de agentes biológicos y químicos y siembra de variedades resistentes. Esta última medida es considerada como una de las soluciones más prácticas y eficientes (Jennings y Pineda 1971; Martínez 1971).
Observaciones de campo realizadas en los años 1967 y 1968 en Palmira (Valle del Cauca), con respecto a la variedad IR 8, indicaron que ésta se encontraba relativamente libre de sogata, mientras que la variedad Bluebonnet 50, sembrada en una parcela adjunta, presentó daño. Esta observación indicó que existen diferencias entre los genotipos en cuanto al daño producido por el insecto sogata, sugiriendo que deben comenzarse estudios detallados sobre la resistencia de la planta al insecto (Jennings y Pineda 1970a).
Con base en los estudios conocidos del insecto y los realizados sobre el mejoramiento genético de la planta, se propuso estudiar la base genética de algunos materiales seleccionados como resistentes al daño mecánico del insecto. Los objetivos del presente estudio fueron:
Caracterizar doce cultivares de arroz al daño mecánico de T. orizicolus.
Estudiar el modo de herencia de la resistencia en dos cultivares resistentes.
Materiales y Métodos
Caracterización de cultivares
El trabajo se realizó bajo condiciones de invernadero en el Centro Internacional de Agricultura Tropical-CIAT, en Palmira, durante los meses de marzo a diciembre de 1992, utilizando insectos de una colonia libre de virus, multiplicada en los invernaderos del CIAT.
La genealogía de algunas variedades registradas como resistentes al daño mecánico se analizó, y se seleccionaron ocho posibles fuentes de resistencia (Tabla 1), las cuales se compararon con cuatro testigos susceptibles: IR 8 (ICA), IR 8 (IRRI), Colombia 1 y Bluebonnet 50. La caracterización de los materiales se hizo mediante las pruebas de libre escogencia, no escogencia, supervivencia y oviposición.
Derivación de las posibles fuentes de resistencia al daño mecánico de T. orizicolus.
Mudgo es un cultivar tradicional del Asia y se desconocen sus progenitores.
Prueba de libre escogencia
La siembra de los 12 cultivares se realizó en bandejas plásticas de 27 cm de ancho por 53 cm de largo y 6 cm de alto, en surcos individuales, distribuidos al azar dentro de las bandejas. Diez días después de la siembra se realizo un raleo y se dejaron 10 plantas/surco. A los 5 días después del raleo, las bandejas se introdujeron en una jaula de malla de 1,95 m de largo, 1,10 m de ancho y 1 m de altura y se infestaron con un promedio de 10 insectos/planta. La prueba se realizó tres veces con seis repeticiones; cada bandeja constituyó una repetición. El diseño experimental fue el de bloques completos al azar. Cada surco se calificó en forma individual a partir del primer día de infestación, utilizando la escala de evaluación 0-9 (Tabla 2).
Escala de evaluación modificada en el CIAT para evaluar el daño mecánico de T. orizicolus.
Fuente: Pantoja y Hernández (1993)
Prueba de no escogencia
Los 12 cultivares se sembraron en materos de 10 cm de diámetro por 10 cm de alto, utilizando cinco semillas/cultivar, pero al final se dejaron sólo dos plantas/matero. Quince días después de la siembra, cada matero se cubrió con una jaula de muselina y se infestó con 10 insectos. La prueba se realizó una vez, utilizando siete materos por cultivar y se dejó un matero como testigo sin infestar. El diseño experimental fue el de bloques completos al azar. Las evaluaciones se realizaron diariamente a partir del día siguiente a la infestación, utilizando la escala de evaluación a daño mecánico a T. orizicolus (Tabla 2).
Prueba de supervivencia y oviposición
Los 12 cultivares se sembraron en materos de 10 cm de diámetro por 10 cm de alto, distribuidos al azar, empleando cinco semillas/cultivar y luego se dejó una planta/matero. A los 15 días de edad, las plantas se cubrieron con un tubo de acetato transparente y se infestaron con una pareja de insectos vírgenes. La prueba se realizó tres veces con cinco materos/variedad, bajo un diseño experimental de bloques completos al azar. Las variables de respuesta fueron la supervivencia de la hembra y el número de huevos/semana (oviposición); ambas evaluaciones se realizaron simultáneamente. Cada semana, la pareja de insectos se pasó a otra planta de la misma variedad y edad (15 días) y las plantas reemplazadas se observaron bajo el microscopio para contabilizar los huevos depositados por las hembras. El traslado de los insectos se hizo semanalmente hasta cuando la hembra murió.
Evaluación de las poblaciones F₁ y F₃
El trabajo experimental se realizó en los meses de marzo a junio de 1993, en los invernaderos del CIAT. Los cultivares resistentes, seleccionados a partir del primer grupo de experimentos fueron: Mudgo y Makalioka; éstos se cruzaron con la variedad susceptible IR 8 (Tabla 3). Las generaciones F₁ y F₃ de cada uno de los cruzamientos, y los progenitores se evaluaron mediante la prueba de libre escogencia. De las poblaciones F₁ se evaluaron 10 plantas/cruce y progenitor, y de la población F₃ se evaluaron 100 familias y 75 plantas/familia. Las evaluaciones se realizaron individualmente (planta por planta) cuando el testigo susceptible IR 8 presentó un daño igual a 9.
Relación de los cruzamientos estudiados.
El análisis de los datos de los cruces de la F₁ se realizó en forma conjunta, a diferencia de la población F₃ que se realizó en forma individual. Los datos se expresaron como porcentaje de plantas susceptibles. En la F₃, las familias se clasificaron en grupos de porcentaje de susceptibilidad en rango de 10 unidades (eje X) y frecuencia de familias (eje Y); posteriormente se agruparon en categorías para estimar el modelo genético más apropiado. La hipótesis sobre la validez de los modelos genéticos se probó aplicando la prueba de Chi-cuadrado, usando el paquete estadístico SAS (SAS Institute 1985).
Resultados
Prueba de libre escogencia
Tres grupos de cultivares presentaron daño mecánico superior a 7 durante la evaluación (Tabla 4); sin embargo, los grupos presentaron diferencias en cuanto al tiempo requerido para alcanzar este grado. Al día siguiente de la infestación, el primer grupo de cultivares que mostró este grado de daño, estuvo conformado por Chianan 8, Bluebonnet 50 y Colombia 1. A los 16 días de la infestación, el daño lo presentaron IR 8(ICA) e IR 8 (IRRI), y, finalmente, a los 19 días, Cica 8 y Tetep.
Reacción de doce cultivares de arroz al daño mecánico de T. orizicolus según la escala de evaluación del CIAT (0-9) y los días de exposición al insecto bajo libre escogencia.
En cada columna, promedios seguidos por la misma letra no son estadísticamente diferentes según la Prueba de Rangos Múltiples de Duncan.
Los cultivares que sufrieron menor daño al final de la evaluación, grado menor de 5, fueron: Mudgo, Amistad 82, IRAT 120, IRAT 124 y Makalioka. Los cultivares IRAT 124 y IRAT 120 provienen del mismo cruzamiento; sin embargo, su comportamiento fue distinto y al final hubo diferencias significativas entre ellos; IRAT 124 fue muy similar en su comportamiento a su progenitor Makalioka.
Prueba de no escogencia
De igual forma que en la prueba de libre escogencia, esta prueba mostró tres grupos de cultivares que presentaron daño superior a 7 durante la evaluación (Tabla 5). Chianan 8, Bluebonnet 50 y Colombia 1 mostraron el daño a los 15 días de infestadas las plantas, en tanto que Cica 8 e IR 8 alcanzaron el mismo daño a los 22 días después de la infestación, y Tetep a los 31 días después. Los cultivares Mudgo, Amistad 82, IRAT 120, IRAT 124 y Makalioka fueron los que menor daño mostraron. Los cultivares que tuvieron un comportamiento similar a Mudgo (testigo resistente) fueron IRAT 124 y Makalioka. Amistad 82 e IRAT 120 sufrieron daño, pero no presentaron plantas muertas por el insecto.
Reacción de doce cultivares de arroz al daño mecánico de T. orizicolus según la escala de evaluación del CIAT (0-9) de acuerdo con los días de exposición al insecto bajo no escogencia.
En cada columna, valores seguidos por la misma letra no son estadísticamente diferentes según la Prueba de Rangos Múltiples de Duncan.
Prueba de supervivencia y oviposición
La supervivencia de la hembra fluctuó entre 1,5 y 4,0 semanas en Makalioka y Chianan 8, respectivamente (Tabla 6). Los cultivares en los cuales la supervivencia fue similar a Makalioka fueron: Amistad 82, IRAT 120, Mudgo e IRAT 124, que también presentaron poco daño mecánico (Tablas 4 y 5). La supervivencia fue similar a Chianan 8 en los cultivares Bluebonnet 50, Colombia 1, Tetep, IR 8 (ICA), IR 8 (IRRI) y Cica 8, los cuales fueron susceptibles al daño del insecto. Estos resultados indican que T. orizicolus vive más tiempo sobre variedades susceptibles, lo cual es consistente con observaciones previas (Jennings y Pineda 1970b).
Supervivencia y oviposición de las hembras de T. orizicolus sobre doce cultivares de arroz 1
En cada columna promedios seguidos de la misma letra no son estadísticamente diferentes según la Prueba de Rangos Múltiples de Duncan.
El número de huevos/semana varió entre 114,7 en Chianan 8 y 9,93 en Makalioka. Los cultivares que presentaron un número de huevos similar a Chianan 8 fueron los mismos en los cuales la supervivencia de la hembra fue alta; los cultivares que presentaron baja oviposición fueron los mismos que mostraron baja supervivencia de la hembra, a excepción de Amistad 82. La oviposición sobre Amistad 82 fue similar a la observada sobre Bluebonnet 50 (testigo susceptible), pero la supervivencia de la hembra fue similar a Makalioka (resistente).
Estos resultados sugieren que el posible mecanismo que opera en Amistad 82 es el de tolerancia, pues la planta soporta niveles de población del insecto tan altos como los que se alcanzan en variedades susceptibles, pero la planta no sufre daño (Isjakov et al. 1986), o que la supervivencia del insecto en Amistad 82 puede estar controlada por factores distintos a aquellos que afectan la oviposición.
Evaluación de las poblaciones F₁y F₃
En la evaluación de la generación F₁, en la cual se sometieron a prueba los cruces IR 8/Mudgo, IR 8/Makalioka y Makalioka/Mudgo, y los progenitores IR 8, Makalioka y Mudgo, los parentales Mudgo y Makalioka se comportaron como resistentes (0% de plantas susceptibles) e IR 8 como susceptible (80% de plantas susceptibles) (Tabla 7). Al comparar las progenies F₁, las cuales en teoría debían comportarse de manera relativamente homogénea por ser todos los individuos iguales pero de constitución genética heterocigota, se observó una proporción variable de plantas con reacción susceptible. En principio se planteó la existencia de un gen simple dominante (A), presente en estado homocigoto en los parentales Mudgo y Makalioka (AA), como responsable de la resistencia y su respectivo alelo recesivo (a) presente en estado homocigoto en IR 8 (aa), como determinante de la susceptibilidad. Según los resultados observados, el grado de expresividad de este gen estaría supeditado a la interacción con otros factores genéticos y ambientales. El efecto se hizo más evidente en la evaluación de la generación F₃, donde se sembraron simultáneamente 100 familias (con 75 plantas/familia) y los respectivos progenitores (IR 8, Mudgo y Makalioka).
Porcentaje de plantas susceptibles al daño mecánico de T. orizicolus en las poblaciones F₁ y F₃ de los cruzamientos IR 8/Mudgo, IR 8/ Makalioka y Makalioka/Mudgo.
Ver Figuras 1, 2 y 3 que visualizan la distribución de las familias en categorías según el porcentaje de plantas susceptibles.
Los progenitores resistentes Mudgo y Makalioka presentaron un porcentaje de plantas susceptibles de 25 y 41%, respectivamente, en contraste con el 0% observado en la evaluación de la F₁; el progenitor IR 8 confirmó su condición de susceptibilidad (Tabla 7). Este hecho, sumado a la manera como segregaron las familias F₃ (con una proporción variable de individuos susceptibles desde 0 hasta 100%) (Fig. 1, 2 y 3), condujo a postular la hipótesis de que la herencia para la reacción al daño mecánico de sogata es debida a la acción de un gen principal dominante (A), el cual confiere resistencia y cuya expresividad está condicionada por la interacción de un gen modificador (B).
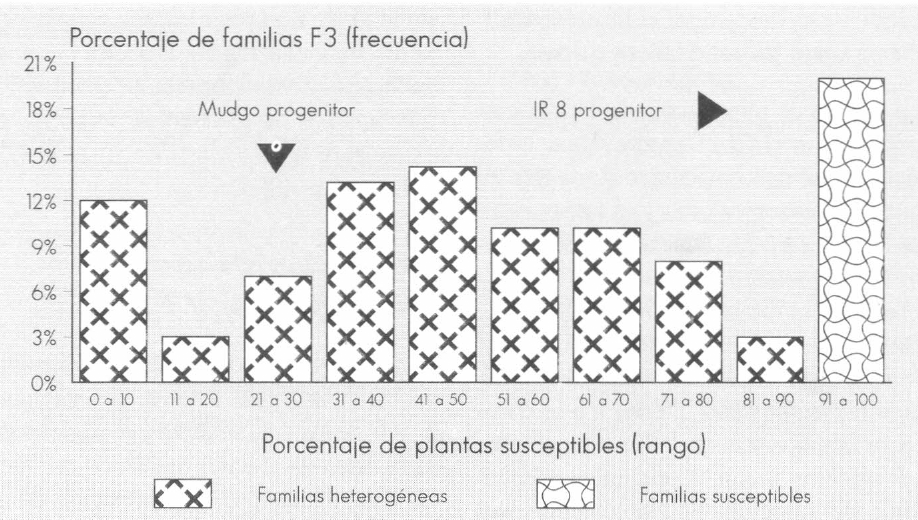
Distribución de 100 familias F3 del cruce IR 8/Mudgo según el rango de porcentaje de plantas susceptibles al daño mecánico de T. orizicolus.
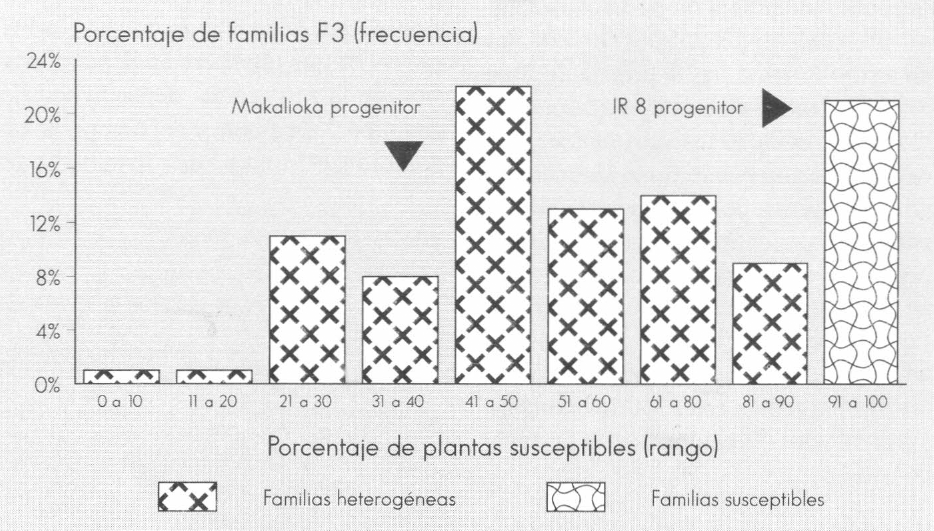
Distribución de 100 familias F3 del cruce IR 8/Makalioca según el rango de porcentaje de plantas susceptibles al daño mecánico de T. orizicolus.
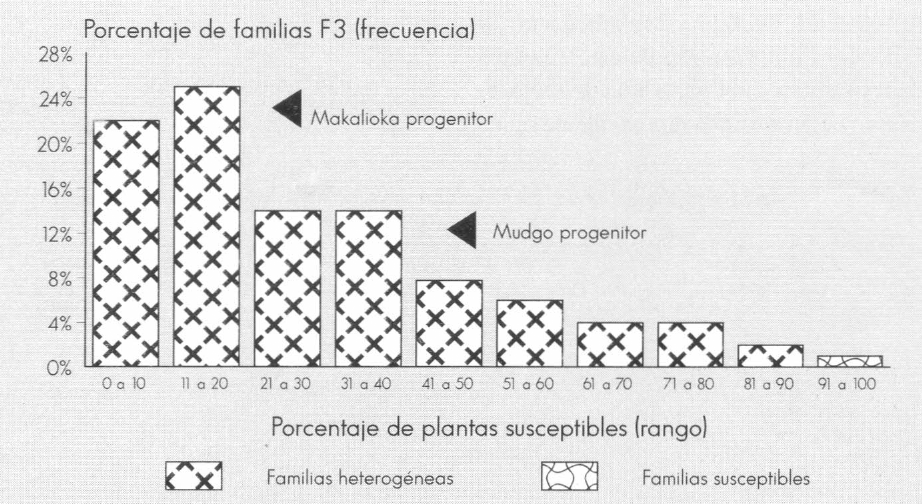
Distribución de 100 familias F3 del cruce Makalioca/Mudgo según el rango de porcentaje de plantas susceptibles al daño mecánico de T. orizicolus.
En la Tabla 8 se hace una descripción detallada del modelo genético, y aparecen los parentales IR 8, Mudgo y Makalioka con el genotipo propuesto; la respectiva Prueba del Chi cuadrado se consigna en la Tabla 9 para la aceptación de dicha hipótesis.
Segregación genotípica y fenotípica esperada para la resistencia al daño mecánico de T. orizicolus con base al modelo propuesto.
Clasificación de las familias F3 de los cruzamientos por su reacción al daño mecánico de T. orizicolus.
O = Observado P.T. = Proporción Teórica E = Esperado Xβ tabulado = se acepta la hipótesis nula
= Familias que presentan una mezcla de plantas resistentes y susceptibles en porcentajes definidos
Discusión
Caracterización de cultivares
El estudio se realizó bajo condiciones de invernadero con el fin de caracterizar nuevas fuentes de resistencia al daño mecánico de sogata que pudieran ser utilizadas en un programa de mejoramiento, con el fin de ampliar la base genética de la resistencia varietal y reducir las posibilidades de aparición de biotipos del insecto.
Existen diferencias entre las condiciones de invernadero y las encontradas en el campo, especialmente en lo relacionado con las interacciones genotipo-medio ambiente e insecto-planta. Además, en las evaluaciones hechas en el invernadero generalmente se utiliza la prueba de libre escogencia, en donde el insecto puede escoger entre un grupo de cultivares puestos a su disposición, en tanto que bajo condiciones de campo, el insecto se alimenta sobre las variedades presentes
La prueba de libre escogencia es la más utilizada en el CIAT para evaluar la resistencia al daño mecánico de sogata en poblaciones segregantes y en líneas avanzadas. La variedad Bluebonnet 50 se utiliza como testigo susceptible y las evaluaciones se hacen cuando las plantas de Bluebonnet 50 han muerto; los cultivares o líneas que según la escala de evaluación presentan daño inferior a 5 se clasifican como resistentes. En esta prueba, los insectos tienen la opción de alimentarse y multiplicarse sobre los cultivares que prefieren, a diferencia de la prueba de no escogencia en donde el insecto es obligado a alimentarse en cada cultivar, situación que refleja mejor las condiciones presentes en los campos comerciales de arroz. Además, en la prueba de libre escogencia el nivel de infestación promedio inicial es de 10 insectos/planta, pero esta infestación puede verse alterada por un incremento de los insectos en los cultivares susceptibles, aquellos que son preferidos por el insecto; por consiguiente, la aparición de daño mecánico es más rápida en esta prueba que en la de no escogencia. A pesar de las pequeñas diferencias existentes entre las dos pruebas, los resultados obtenidos fueron bastante consistentes.
Los cultivares IRAT 120 e IRAT 124 se han utilizado recientemente con cierta frecuencia como fuente de resistencia al daño mecánico de sogata, en tanto que Amistad 82, Mudgo y Makalioka no lo han sido. La utilización de estas fuentes de resistencia en el programa de mejoramiento varietal brinda una excelente oportunidad para ampliar la base genética de la resistencia a sogata. Por otra parte, es necesario y muy importante estudiar los mecanismos de resistencia presentes en estos cultivares (Martínez et al. 1977; Triana et al. 1994).
Evaluación de las poblaciones F₁ y F₃
Con base en los resultados encontrados en las evaluaciones de las generaciones F₁ y F₃ de los cruzamientos, se sugiere que la resistencia al daño mecánico producido por sogata se debe a la acción de un gen principal dominante (A) presente en el estado homocigoto en los parentales Mudgo y Makalioka, el cual confiere resistencia y cuya expresividad está condicionada por la acción de un gen modificador (B) que interfiere en la manifestación de la resistencia, dependiendo del genotipo de la planta y del tiempo de exposición al ataque del insecto; para Mudgo se propone que el gen modificador se encuentra en la forma homocigota recesiva (bb), lo cual acelera la expresión del fenotipo susceptible, y para Makalioka se propone en la forma homocigota dominante (BB), la cual retarda la expresión del fenotipo susceptible.
Con respecto al modelo se aclara que el gen modificador (B) actúa en forma diferencial sobre la expresión del gen principal (A), dependiendo de las formas alélicas presentes en el locus, así: BB+ reduce en forma leve la resistencia de un genotipo, dando lugar a la presencia de una proporción baja de plantas susceptibles en la familia; Bb++ reduce la resistencia de un genotipo a nivel moderado, dando lugar a una proporción intermedia de plantas susceptibles en la familia; y bb+++ afecta en forma significativa la resistencia de un genotipo, motivando la presencia de una proporción alta de plantas susceptibles.
Conclusiones
Los cultivares Chianan 8, Bluebonnet 50, Colombia 1, IR 8 (ICA), IR 8 (IRRI), Cica 8 y Tetep se clasificaron como susceptibles al daño mecánico de T. orizicolus y los cultivares Amistad 82, IRAT 120, IRAT 124, Mudgo y Makalioka como resistentes.
La herencia de la resistencia al daño mecánico para sogata en los cultivares Mudgo y Makalioka es debida a la acción de un gen principal dominante (A) que confiere resistencia y cuya expresividad está controlada por la acción de un gen modificador (B); para Mudgo se propone el gen de modificador en forma homocigota recesiva (bb) y para Makalioka en forma homocigota dominante (BB).