
Abstract
In order to quantify the decrease in quality and production of G. paniculata, the difference between the mechanical leaf prunning and the leafminer Liriomyza huidobrensis (Blanchard) damage was evaluated. Two trials were conducted to compare the defoliation against the damage produced by two leafminer populations. The first trial was conducted in Madrid (Cundinamarca) at 2.600 masl. Four levels of defoliation (0, 25, 50 and 75%) were evaluated at four crop stages: 1, 5, 9 and 13 weeks. In the second trial, in Santafé de Bogotá, three leafminer adult densities were evaluated: 0, 0.7 and 1.3 adults per leaf of Gypsophila at the same four crop stages. The defoliation did not affect the production severely, when it ocurred only once at any crop stage, decreasing only 22% with a 75% defoliation on the 9th week. This is the most susceptible crop stage to leaf loosing. The defoliation of 25% in week 13th increased production in 13,8%. Any defoliation at weeks 1 and 5 retarded the harvest between 6 and 12 days, without affecting the flower quality. In the second trial, the densities of 0.7 adults per leaf on week 5th and 1.3 on week 9th decreased production in 38 and 46.7% respectively. The leafminer did not decreased the production on week 13th, but when the attack ocurred during the first week it retarded, from 15 to 20 days, the beginning of the harvest. The flower and stem biomass was affected by the leafminer on weeks 5 and 9.
Introducción
En Colombia se han duplicado las ventas de flores en los últimos años, ya que de 100 millones de dólares en 1980 pasó a 205 millones en 1988 (Garavito 1991). El cultivo de Gypsophila (Gypsophila paniculata L.) ha tenido gran auge en los últimos años, y una de las razones de este hecho es el incremento en su demanda, ya que es muy apetecida por su forma de crecimiento que llena adecuadamente espacios.
En 1980, Colombia participó en el mercado estadounidense con el 83,6% de la Gypsophila, pero esta cifra se ha reducido considerablemente y llegó al 33,8% en 1989 (Asocolflores 1992); la causa de esta disminución en la participación puede ser la pérdida de capacidad competitiva dentro del mercado exterior o por los problemas propios del mercado.
Dentro de los insectos indeseables de este cultivo se encuentra el minador de follaje Liriomyza huidobrensis (Blanchard) (Diptera: Agromyzidae), y por este motivo el mercado europeo exige la eliminación completa de las hojas, pues temen la entrada de la plaga a su territorio. La importancia de conocer la influencia que tiene Liriomyza sobre la calidad y producción de la Gypsophila es esencial para realizar un manejo adecuado del problema, tratando de reducir los costos de producción que actualmente son muy altos, pues aproximadamente se invierten $3'000.000/ciclo/hectárea en aplicaciones de insecticidas (Arias 1993) 1 , con la consiguiente contaminación del medio ambiente.
ARIAS, C. 1993. Comunicación personal. Antes Ing. Agrónomo de Floramérica S. A.
Con el interés científico de conocer si el corte manual diario de un 25 hasta un 75% del área foliar simula adecuadamente el daño producido por el minador, se planeó la ejecución del presente trabajo, que tuvo como principal objetivo la cuantificación de la disminución en calidad y producción del cultivo de G. paniculata al realizar un daño manual en el follaje, en cuatro edades del cultivo, relacionándolo con el efecto producido por dos densidades de población de adultos del minador L. huidobrensis.
Revisión de Literatura
El minador
El insecto pertenece a la Clase Insecta, Orden Diptera, Familia Agromyzidae, Subfamilia Phytomyzinae, Genero Liriomyza, Especie huidobrensis (Blanchard). El huevo dura en promedio de 3 a 6 días, la larva entre 10 y 17 días, la prepupa 9,25 horas, la pupa entre 8 y 17 días y el adulto entre 4 y 6 días, siendo mayor la longevidad de las hembras (Ledieu y Bartlett 1983; Sarmiento et al. 1986; Salas et al. 1988). La relación de sexos es de 1:1 (Parrella 1987; Sarmiento et al. 1986) y la longevidad de las hembras se incrementa linealmente a medida que se incrementa la temperatura (Zoebisch et al. 1992). Salas et al. (1988) encontraron una relación de 1,44:1 entre hembras y machos de la especie L. trifolii (Burgess).
La relación de punturas totales a punturas de oviposición varía de 1:1 a 40:1, cambiando con la temperatura, la calidad de la hoja y la planta hospedante (Parrella 1987; Zoebisch et al. 1992); Sarmiento et al. (1986) confirmaron la relación 40:1. La cantidad de huevos que la hembra puede colocar varía entre 100 y 600 (Parrella 1987).
En Gypsophila, la relación puntura/mina es 25,5% mientras que en fríjol es de 4,1% (Van Elferen y Yathom 1989). Al parecer, la base de la preferencia alimenticia del minador es genética (Parrella 1987), ya que mientras la larva de L. trifolii se encuentra en el mesófilo de empalizada, la de L. huidobrensis mina el mesófilo esponjoso (Spencer 1973, citado por Parrella et al. 1985). En daños leves, L. huidobrensis reduce la fotosíntesis del crisantemo en un 80,5%, encontrándose mayor reducción al incrementarse el daño. L. trifolii no reduce mucho la fotosíntesis porque no afecta las funciones estomatales.
Defoliación
Las plantas responden diferencialmente al daño por defoliación, dependiendo: de la edad en la que se presente, de si la defoliación es puntual o continua, de la cantidad de área foliar que se pierda, del sitio de la planta donde se produzca el daño, de la nutrición y movimiento del agua dentro de la planta, de factores climáticos y de la competencia con otras plantas, principalmente por luz.
En haba (Vicia faba L.), la pérdida del 66 y 100% de las hojas en los períodos de prefloración tardía, fructificación temprana y maduración de frutos reducen estadísticamente la producción (Coggin y Dively 1980).
La yuca (Manihot esculenta Crantz) es muy susceptible a la defoliación, sobre todo en suelos poco fértiles, donde la producción puede caer hasta un 46% (Arias y Bellotti 1984).
Dentro de otras respuestas de las plantas, se incrementa la producción bajo condiciones especiales de cultivo y de la pérdida de área foliar en sí. El incremento de ramas del cultivo de papa (Solanum tuberosum L.), luego de una defoliación severa, se debe al rompimiento de la dominancia apical, pudiendo explicar la mayor tuberización. Una defoliación leve podría no romper esta dominancia (Harris 1974, citado por Cranshaw y Radcliffe 1980). Jaramillo et al. (1989), al estudiar el nivel económico de daño de la polilla de la papa, Phthorimaea operculella (Zeller), en Rionegro (Ant.), encontraron que con una defoliación del 75% se incrementaron más los rendimientos que con una defoliación del 50%; el incremento fue más notorio al realizar el tratamiento de defoliación a los 35 días después de la siembra.
La remoción de hojas viejas y poco eficientes produce una mayor desviación de los nutrientes a las partes más fotosintéticamente activas (Humphries 1967, citado por Cranshaw y Radcliffe 1980). Mientras que Peterson et al. (1992) observaron que la transpiración y la conductancia estomatal de la alfalfa fueron mayores en hojas afectadas por gorgojo que en hojas sanas, aunque las ratas fotosintéticas fueron similares; este comportamiento los llevó a descubrir que el principal efecto del daño por el gorgojo es reducir el incremento del tejido foliar que fotosintetiza sin afectar las tasas fotosintéticas.
Los desbalances fuente-vertedero creados por la defoliación son corregidos por respuestas integrales que demuestran un gran grado de plasticidad fisiológica en las plantas (Chapman et al. 1991); estos mecanismos se definen como compensación y tolerancia. Hearn y Room (1979), citados por Brook et al. (1992), definen la compensación como el crecimiento y desarrollo que no ocurriría si no existiera una condición de estrés, mientras que la tolerancia es la sustitución pasiva de estructuras y frutos deteriorados por otros, ya que ese deterioro sustituye mecanismos de desprendimiento fisiológico. Dentro de las formas de compensación de las plantas se encuentran: el crecimiento de las ratas fotosintéticas de las hojas que permanecen (Detling et al. 1979; Harris 1974 y Neales e Incoll 1968, citados por Capinera y Roltsch 1980), el rompimiento de la dominancia apical (Hillmann 1984 y Cline 1991, citados por Ovaska et al. 1993) y el incremento del potencial para absorber nutrientes (Chapin y Slack 1979 y Coughenour et al. 1990, citados por Ovaska et al. 1993).
Gypsophila
Esta planta experimental pertenece a la familia Caryophillaceae y es de origen eurasiático (Shillo 1985). Es una planta herbácea y perenne que posee flores grandes y blancas de gran volumen, lo cual la hace preferida para el comercio. Las ramificaciones aparecen desde la base de la planta; bajo condiciones no inductivas, la planta permanece en una forma similar a una roseta, que es un racimo de muchas ramas laterales no elongadas. Bajo condiciones inductivas con luz artificial, tiene lugar la elongación del tallo central y de los laterales, comenzando el crecimiento rápido (Shillo 1985). Para este evento, la Gypsophila requiere baja cantidad de nutrientes. Antes de la floración se debe mantener una relación de N:P:K similar a 1:1:1, para luego reemplazarla por una relación 1:2:1 hasta el final del cultivo (Danziger s.f.).
En el comportamiento del crecimiento de G. paniculata se definen tres etapas: la primera corresponde a la acumulación de energía, que dura entre tres y cuatro semanas; la segunda al desarrollo de ramas reproductivas que dura seis o siete semanas y la tercera a la producción de flores, que va hasta el cese de la floración.
Dentro de los factores que influyen en la floración se encuentran: el fotoperíodo, que está en el rango de 12 a 18 horas, ya que a mayor longitud del día se produce mayor número de flores (Shillo 1985; Shillo y Halevy 1982; Kusey et al. 1981), la radiación solar y la temperatura, porque al aumentar, se incrementa el número de tallos florales por planta (Shillo 1985).
Materiales y Métodos
Ensayo de defoliación
Este ensayo se realizó entre los meses de abril a septiembre de 1993 en Floramérica S.A., en Madrid (Cund.), con temperatura promedia de 23°C (bajo invernadero) y 2.600 msnm. Se utilizó un diseño de bloques completos al azar con un arreglo factorial 4x4, con tres replicaciones. El primer factor, la defoliación, se definió con cuatro niveles: 0, 25, 50 y 75% de área foliar perdida. El segundo factor, la edad del cultivo, tuvo cuatro niveles: 1, 5, 9 y 13 semanas de cultivo, luego del trasplante. Las replicaciones se conformaron en tres camas del invernadero. En cada una de ellas se instaló riego por goteo y bombillos incandescentes de 220 v y 200 w (0,25 bombillos/mβ) para darle luz artificial al cultivo. Los tratamientos se distribuyeron en las tres replicaciones (tres camas), siendo cada tratamiento la combinación de los niveles de los dos factores. Se realizaron las labores de cultivo esenciales, como son: tutorado, encanaste, deshierbe y fertilización continua, proporcionando 278 ppm de N, 89 ppm de K, 4 ppm de P, 12 ppm de Mg, 1 ppm de Fe y 0,135 ppm de Mo.
Para realizar los tratamientos se tomaron dos parámetros fundamentales: el primero, defoliar en hojas completamente formadas para evitar respuestas fisiológicas al cortar las hojas de los retoños de cada rama; y el segundo, homogeneizar el tipo de daño al realizar el corte a todas las hojas, evitando la dificultad de tomar hojas al azar dentro de los estratos de la planta, labor que aumentaría el sesgo experimental.
Para el modelo de defoliación se tomó la nervadura central como la mitad de la hoja, se marcaron puntos cada cinco milímetros sobre ella (tomándola como eje. x), y sobre esos puntos marcados, se midió la distancia perpendicular hacia el borde de la hoja. Al hacer esta medición se obtuvieron datos del área foliar para cada uno de los intervalos, según la fórmula:
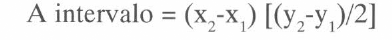
A intervalo = Área del intervalo sobre la nervadura central.
(x₂-x₁) = intervalo (5 mm).
у₂ = Distancia perpendicular hasta el borde de la hoja en el punto x₂.
у₁ = Distancia perpendicular hasta el borde de la hoja en el punto x₁.
Al acumular todos los datos de área de intervalo desde la base hacia el ápice de la hoja, se obtuvieron los intervalos donde se encuentran el 25, 50 y 75% de área foliar, y además se obtuvo el dato del área total de la hoja (Tabla 1) mediante la fórmula:
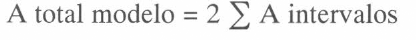
El corte se realizó con tijera, quitando diariamente un pedazo de 25% de área de cada hoja (según el modelo), ya que se trató de simular el daño de insectos, y éstos no atacan áreas tan grandes en un solo día. Al llegar la fase de cosecha, los tallos florales se cortaron cuando el 33% de las flores se encontraban abiertas; con una longitud mayor de 60 cm, desde el nivel de las flores hacia el tallo; estos valores son considerados como ideales para el mercado de exportación.
Tipos de hojas de Gypsophila paniculata L. según su tamaño e intervalos de corte (25, 50 y 75% de área foliar), obtenidos para la realización de los tratamientos de defoliación.
Ensayo de daño del minador L. huidobrensis
Este ensayo se realizó en los invernaderos de la Universidad Nacional de Colombia en Santafé de Bogotá, a 2.600 msnm y con una temperatura media dentro del invernadero de 21°C, durante los meses de septiembre de 1993 a marzo de 1994.
Se utilizó un diseño de bloques completos al azar con arreglo factorial 3x4 con tres replicaciones, donde los factores fueron: la densidad de población de adultos de minador L. huidobrensis con tres niveles: 0, 0,7 y 1,3 minadores por hoja de Gypsophila; y la edad del cultivo con cuatro niveles: 1, 5, 9 y 13 semanas después del transplante a sitio definitivo.
El riego con fertilizante se aplicó diariamente a las plantas con una regadera, a razón de 950 cm³ de solución por planta. El fertilizante utilizado fue igual al del ensayo de defoliación, con las mismas concentraciones. Se instalaron bombillos de 110 v y 150 w, a razón de 0,66 bombillos por mβ. Se dieron condiciones de luz artificial continua al cultivo durante ocho horas nocturnas. Como el ensayo se realizó con poblaciones de minador, fue necesario hacer un programa de aplicaciones de insecticidas para las plantas que no tuvieran tratamiento de infestación.
La densidad del minador se estableció a partir de la experiencia de Sarmiento et al. (1986), quienes manejaron la densidad de un minador por hoja de Gypsophila para la realización de su trabajo. Según este parámetro, se optó por tomar un número mayor (1,3) y menor (0,7) al dato propuesto, con el fin de obtener un daño diferente en las plantas con cada uno de los tratamientos mencionados.
Para realizar los tratamientos se contó el número de hojas de las seis plantas de las unidades experimentales a infestar y se multiplicó por la densidad de insectos que correspondió a dichas unidades.
Con una jaula de malla de 2,35 m de largo, 1,45 m de ancho y 1,80 m de altura se cubrieron las seis plantas de las unidades experimentales, y dentro de ella se liberaron las correspondientes poblaciones de minador. Los minadores se obtuvieron mediante aspiración con una máquina D-Vac, en un cultivo comercial de Gypsophila. Preliminarmente se llevó una muestra al Instituto de Ciencias Naturales donde se determinó que la mayoría de los insectos pertenece a la especie L. huidobrensis. Dentro de las jaulas, los minadores permanecieron durante 72 horas, y después se aniquilaron con insecticidas.
En los dos ensayos se midieron las siguientes variables:
Peso total de tallos florales por planta. Esta variable es la producción final de cada planta expresada en gramos, obtenida mediante la suma de los pesos de los tallos cosechados en cada fecha de corte. Peso total de material no comercial por planta. Esta variable proviene del corte de los tallos florales. Es el peso de estructuras que por ser vegetativas o poco desarrolladas deben ser eliminadas para <<tmaquillar>> los tallos que se lleven al mercado internacional. Comportamiento de la cosecha en el transcurso del tiempo. Son los días en que la suma del peso de los tallos florales cosechados por planta alcanza a ser el 0% (comienzo), 50% у 100% (final de la cosecha) de la totalidad de tallos cortados. Número de tallos basales por planta. Es la cantidad de ramas formadas a partir del suelo hasta los primeros 4 - 5 cm. Esta variable se midió con el fin de relacionar la producción de macollas (tallos basales) con los tratamientos de defoliación e infestación en cada semana. Biomasa. Se tomó por estructuras: biomasa de tallos, de hojas y de flores (secadas a 105°C durante 48 horas). Esta variable se midió con el fin de conocer las posibles respuestas fisiológicas en la planta, cuando se realizaron los tratamientos de defoliación y de infestación. Área foliar total al momento de los tratamientos. Variable que se tomó sólo en el ensayo con minador, para conocer la respuesta instantánea de las plantas al agente defoliador.
Resultados y Discusión
Ensayo de defoliación
Peso total de tallos florales por planta
La producción final en plantas defoliadas en las semanas de medición no difirió estadísticamente (Tabla 2). El único efecto notorio se encontró en la semana 9, en la cual hubo una reducción no significativa de la producción en 22% con respecto al testigo, cuando la defoliación fue del 75%.
La producción, al defoliar en la semana 13, tendió a incrementarse hasta un 13,8%, con un 25% de pérdida de área foliar con respecto al testigo de esa semana (sin ser significativo). Este comportamiento muestra el posible efecto de recuperación cuando hubo pérdida de área foliar en las semanas evaluadas, haciendo notar que en la etapa de establecimiento (semana 1), la planta respondió positivamente a la pérdida de hojas por una sola vez, hasta un 75%, situación que pudo deberse a que las hojas no cumplen su función de fuente en un 100%, y al ser removidas, la energía de la planta se encaminó a otros procesos como el crecimiento de yemas o elongación de brotes ya formados, sin repercutir en la producción final. No sucedió esto en las semanas 5 y 9, cuando se perdió gran parte del material fotosintéticamente activo, tal vez porque la planta podría experimentar una detención en su crecimiento, en especial cuando ha ocurrido la diferenciación de yemas reproductivas (semana 9).
En la semana 13, las plantas pasaron por el momento de alta producción y una defoliación leve pudo inducir a un incremento de la producción, siendo un posible mecanismo de tolerancia instantánea, como lo registraron Kletter y Wallach (1982), citados por Brook et al. (1992), donde, al parecer, la pérdida leve de estructuras reemplaza procesos de derrame fisiológico o abscisión de hojas viejas, haciendo incrementar la fotosíntesis de las hojas que permanecen en la planta, con lo cual se puede incrementar la producción.
Peso total de material no comercial por planta
Al realizar corte de follaje en la semana 1 no se encontró efecto de cualquier grado de defoliación sobre el peso de material no comercial por planta. En la semana 5, las defoliaciones del 25, 50 y 75% incrementaron el peso de material no comercial en 36, 29 y 31%, respectivamente, con respecto al testigo de esa semana (200,63 g). Para la semana 9, el comportamiento fue similar al de la 5a. semana, aunque el incremento fue de 66,4% con el tratamiento de 50% de defoliación y de 45,5% con el de 75% de defoliación con respecto al testigo (169,3 g). En la 13a. semana, la defoliación redujo no significativamente el peso de material no comercial, sobre todo con 50% de pérdida del área foliar (31,1%), y con 75% de defoliación (10,4%) con respecto al 0% de defoliación (220,8 g) (Tabla 2).
Es posible establecer la relación que existe entre las etapas fisiológicas por las que está pasando la planta cuando es defoliada y la respuesta que ejerce ante esta defoliación: En establecimiento, el desorden metabólico causado por la defoliación puede ser "arreglado" fácilmente por la planta, lo que no sucede en las etapas de crecimiento activo, cuando, al parecer, se incrementa la producción de estructuras vegetativas como mecanismo de recuperación de la planta, quizás por una respuesta de incremento de la brotación axilar. En la semana 13, la planta no consume energía en recuperar las hojas perdidas, porque ha pasado la mayor parte de la floración y es posible que la planta esté entrando en un período de crecimiento vegetativo lento, donde puede disminuir su actividad metabólica.
Comportamiento de la cosecha en el transcurso del tiempo
Esta variable corresponde a los días en que se logró el 0, 50 y 100% de la producción total para cada uno de los tratamientos de defoliación (Fig. 1). Se observó una tendencia a demorar el comienzo de la fase de cosecha en las plantas de los tratamientos del 50 y 75% de defoliación en la semana 1 (tratamientos 3 y 4), en 4 y 9 días de cultivo, respectivamente, con relación al testigo (75 días de cultivo). El 75% de defoliación en la semana 5 también demoró el comienzo de la fase de cosecha en 6 días con respecto al testigo de esa semana (72 días de cultivo).
El 50% de la cosecha posee un comportamiento similar. Las plantas de los tratamientos que llegaron más tarde a este porcentaje de cosecha fueron las de 50 y 75% de defoliación en la semana 1 (12 y 15 días de atraso) y 50 y 75% de defoliación en la 5ª semana (9 días de atraso) con respecto a los respectivos testigos, los cuales llegaron al 50% de la cosecha en 84 días de cultivo. El 100% de la cosecha lo cumplieron todos los tratamientos entre 131 y 140 días, sin tener diferencias significativas.
El comportamiento de las plantas cuando son defoliadas severamente (50 y 75%) en la semana 1 es posible explicarlo como la consecuencia de una reducción del metabolismo luego de la defoliación, como mecanismo de defensa para lograr recuperarse del factor adverso y llegar a la fase reproductiva, lo cual se realiza un tiempo más tarde.
Cuando hubo defoliación en la semana 5, las plantas pudieron tener la misma respuesta, pero esa posible reducción de la tasa de crecimiento les pudo causar una disminución de la producción, porque se encontraban en un período de crecimiento rápido y de acumulación de asimilados para la floración. Las plantas de los tratamientos de la semana 9 no pudieron disminuir su tasa de crecimiento, quizás debido a la inducción de floración, pero por la cantidad reducida de asimilados, la producción se afectó, por lo menos con el 75% de defoliación. En la semana 13, la defoliación no afectó la fase de cosecha, ya que cuando se defolió, se había cosechado más del 50% de la producción.
Comportamiento promedio de las variables peso total de tallos florales por planta, peso total de material no comercial por planta y número de tallos basales de Gypsophila paniculata var. perfecta, en ensayo de defoliación. Madrid, 1993.
Promedio de tres replicaciones
En cada columna, los promedios seguidos por la misma letra no difieren entre sí, según la Prueba de Duncan, con significancia al 5%
Comportamiento de la producción de Gypsophila paniculata L. al cumplir 0% (comienzo de cosecha), 50% y 100% (fin de la cosecha), acorde con los tratamientos de defoliación.
Número de tallos basales por planta al finalizar la cosecha
Aunque se registró una reducción en dos tallos basales con 75% de defoliación en la semana 1, las diferencias entre tratamientos de las semanas observadas no son significativas (Tabla 2).
El análisis de esta variable demuestra que la planta no induce brotación basal como respuesta a la defoliación en las semanas de crecimiento vegetativo, en contraste con lo enunciado por Hillmann (1984), citado por Ovaska et al. (1993), quien aseguró que uno de los procesos fisiológicos compensatorios más importantes en las plantas, es el rompimiento de la dominancia apical. Es posible que los mecanismos de recuperación se centren en la relocalización de asimilados en los retoños apicales, como lo observaron Chapman et al. (1991).
Biomasa
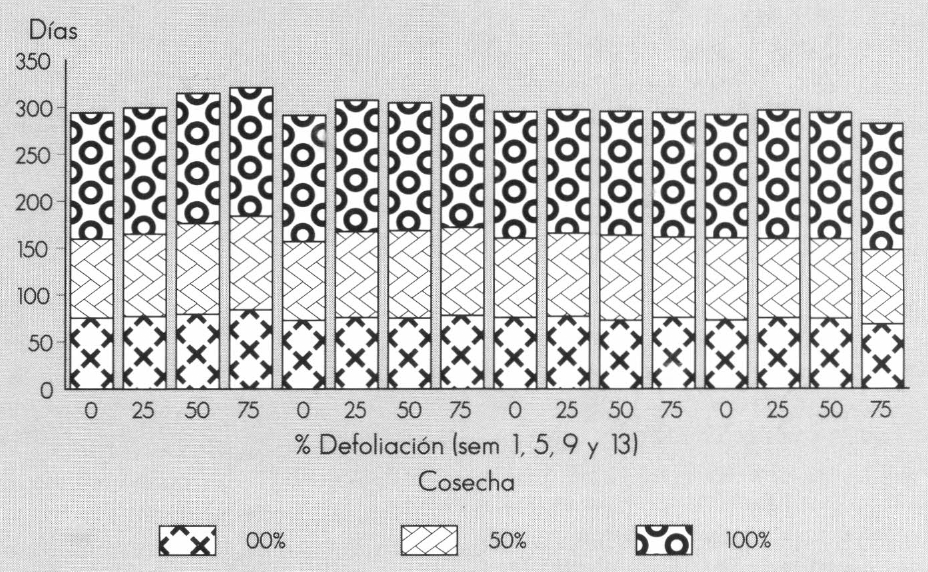
La biomasa de tallos (Fig. 2) no se afectó con ninguno de los grados de defoliación en las semanas 1, 5 y 9, aunque hubo un incremento no significativo con 75% de defoliación en la semana 1 (149,19 g) con respecto al testigo de esa semana (127,13 g). La variable se incrementó con el 25% de defoliación en la semana 13 (156,11 g) con respecto al valor de la biomasa de tallos con 75% de defoliación en la misma semana (108,38 g). Este parámetro es importante porque los tallos representan la mayor cantidad del peso de los tallos florales; además, tallos gruesos y fuertes resisten más los procesos de postcosecha y transporte.
La biomasa de hojas es un parámetro claro de la compensación de la planta a los tratamientos de defoliación. Para esta variable, todos los tratamientos fueron similares estadísticamente, según la prueba de Duncan, al nivel de 5%, lo que indica que a pesar de que las plantas fueron defoliadas hasta un 75% por una sola vez en cuatro edades del cultivo, al final de la fase de cosecha terminaron con un peso seco de hojas similar, evidenciando la tolerancia o compensación a la pérdida de área foliar.
La biomasa de flores no se afectó significativamente por la defoliación en las semanas de crecimiento vegetativo. Para la semana 13, esta variable se incrementó con el 25 y 50% de defoliación (32,7 y 32,9 g), los cuales difirieron estadísticamente al valor de la biomasa de flores cuando hubo 75% de defoliación (22,1 g).
La biomasa total de tallos y de flores mostró la respuesta de las plantas a la defoliación severa (75%) en la semana 1, y a la defoliación leve (25%) en la semana 13. En la primera semana, la posible disminución de la tasa de crecimiento y el realojamiento de asimilados contribuyeron posiblemente al incremento de la eficiencia de la planta, tanto a nivel fotosintético (sin haber respuesta de rompimiento de la dominancia apical) como en la absorción de nutrimentos (Ovaska et al. 1993), produciendo posiblemente tallos más gruesos, aunque con varios días de retraso de la fase de cosecha. En la semana 13 es posible que una defoliación leve (25%) incrementara las tasas fotosintéticas de las hojas jóvenes y de los pedazos de hojas que permanecieron en la planta, produciendo un incremento de la cantidad de asimilados que se alojaban en las estructuras florales y en los tallos.
Comportamiento promedio de la biomasa de estructuras (tallos, hojas y flores) por planta de Gypsophila paniculata L. a los tratamientos de defoliación.
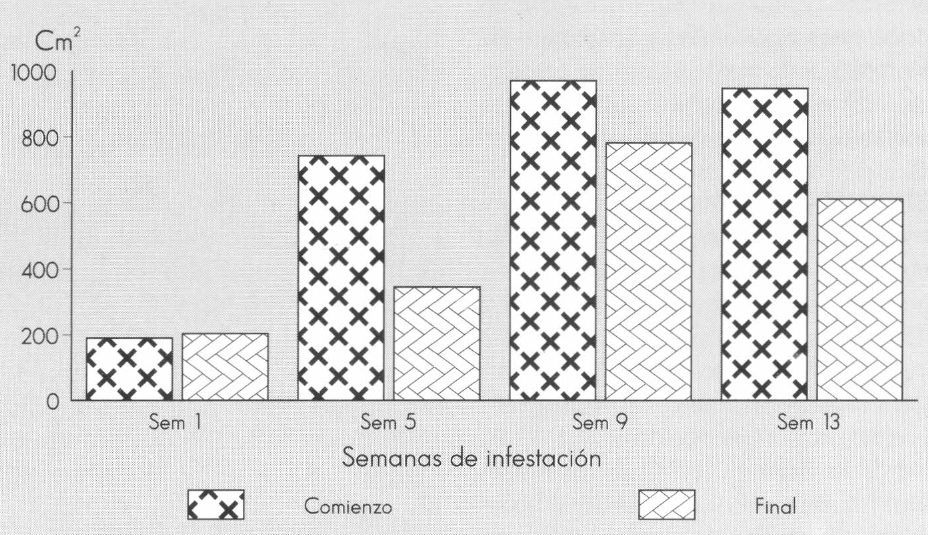
Comportamiento del área foliar de Gypsophila paniculata L. a través del tiempo, al ser infestadas por Liriomyza huidobrensis (Blanchard).
Ensayo de daño del minador
Área foliar total y pérdida al momento de finalizar el daño por punturas y por minas
La presencia del minador disminuyó drásticamente el área foliar (Fig. 3) en la semana 1, cuando la planta posee buena capacidad de recuperación, y durante las tres semanas de permanencia del minador, manteniendo constante su área foliar, sin incrementarla. A partir de las semanas 5, 9 y 13, momentos en los cuales se realizó la infestación, las plantas no pudieron mantener la tasa de producción de hojas a un ritmo por lo menos similar al de la tasa de pérdida de área causada por punturas y larvas de minador durante las tres semanas de permanencia de los estados inmaduros del minador L. huidobrensis. La pérdida de área foliar se encontró entre el 14 y el 49,3%, evidenciando que los minadores no alcanzan a defoliar mayor porcentaje de área, aun siendo poblaciones elevadas como la de 1,3 adultos/hoja.
Peso total de tallos florales por planta
La producción de Gypsophila cuando hubo presencia de minador en la semana 1 se disminuyó en 37,3 y 33% con cualquier cantidad de adultos por hoja (0,7 y 1,3 minadores, respectivamente). Un comportamiento similar tuvieron los tratamientos de la 5a. semana, cuando las densidades media y alta de minador redujeron la producción en 38 y 27,8%, respectivamente. Para la semana 9, la densidad de 1,3 adultos/hoja redujo la producción en 46,7%, con respecto al testigo. En la semana 13, la presencia del minador no afectó el peso total de tallos florales por planta (Tabla 3).
Diferencias estadísticas se encontraron entre el valor de la producción cuando no hubo minador (testigo) en las semanas 1 y 9 (229,63 y 236,92 g, respectivamente) con respecto a la presencia de 0,7 adultos por hoja en la semana 5 y 1,3 en la semana 9 (127,13 y 126,12 g, respectivamente), según la prueba de Duncan al 5% de significancia.
La variable medida indica que, al parecer, en la fase vegetativa el minador es un problema, ya que disminuyó la producción por la permanencia que tuvo durante dos a tres semanas realizando varios tipos de daño como son: la reducción del área foliar y la posible secreción de toxinas en la saliva o excrementos, que posiblemente deterioraron la fisiología de la planta sin importar la edad en que se encontraba.
En la semana 13 sucedió lo mismo que con la defoliación: la planta ya ha pasado por el mayor período de cosecha y resistió el ataque del minador, aunque sin incrementar la producción, quizás por el efecto de la pérdida continua de área foliar que le impidió optimizar mecanismos de tolerancia o compensación.
Comportamiento promedio de las variables peso total de tallos florales por planta, peso total de material no comercial por planta y número de tallos basales de Gypsophila paniculata var. perfecta, en el ensayo con daño del minador Liriomyza huidobrensis (Blanchard). Santafé de Bogotá, 1994.
Promedios de tres replicaciones
En cada columna, los promedios seguidos por la misma letra no difieren entre sí, según la Prueba de Duncan, con significancia al 5%
Peso total de material no comercial por planta
Para la semana 1, la variable se disminuyó en 41,6 y 45,8% a medida que aumentó la densidad de minador. En la semana 5, cualquier presencia de minador no afectó el peso. Para los tratamientos de la semana 9, la producción de material no comercial se redujo con 0,7 adultos/hoja en 45,1% (113,43 g), con respecto al testigo (0 minadores en la semana 9, con 206,62 g), pero con 1,3 adultos/hoja, la reducción fue del 30,2% (144,05 g). En la semana 13, el peso total de material no comercial por planta se incrementó a medida que aumentó la densidad de minador (Tabla 3). Se encontraron diferencias estadísticas entre el testigo de la semana 9 (206,62 g) y las densidades 0,7 y 1,3 adultos/hoja en la semana 1 (96,62 y 89,68 g, respectivamente), y 0,7 adultos/hoja en la semana 9 (113,43 g), según la prueba de Duncan al 5% de significancia.
La disminución del material no comercial de los tallos florales de las plantas infestadas la 1a. semana de cultivo, implica que la planta posiblemente redujo la actividad fotosintética por la presencia del minador, lo que no sucedió en las plantas infestadas la semana 5, que posiblemente intentaron incrementar la tasa de producción de hojas y ramas para alcanzar una acumulación de asimilados que les permitiera llegar a floración, y por esto mantuvieron una mayor cantidad de partes vegetativas a nivel de flor.
En la semana 9, el comportamiento fue diferente, quizás porque los asimilados que se produjeron se enviaron a las flores; al tener la necesidad de incrementar las partes vegetativas, se creó una competencia entre los vertederos. Cuando hubo floración (semana 13), los ataques del minador no redujeron el peso de material no comercial, sino que estimularon la producción de partes vegetativas a nivel de flor, como respuesta a la presencia del minador causando daño.
Comportamiento de la cosecha en el transcurso del tiempo
El comienzo de cosecha (0%) se retardó con cualquier presencia del minador en la primera semana (100 y 105 días de edad para las densidades de 0,7 y 1,3 minadores/hoja, respectivamente) con respecto al testigo (85 días de cultivo). En las otras semanas de infestación, las diferencias no fueron estadísticamente significativas; para las plantas infestadas en las semanas 5 y 9 se observó un retardo que puede ser de atención en un cultivo comercial de Gypsophila (entre 2 a 4 días para plantas infestadas la semana 5, y 8 a 9 días para la semana 9). Al 50 y 100% de la cosecha se presentó el mismo comportamiento con las plantas infestadas la semana 1. En las plantas que fueron infestadas con 1,3 adultos/hoja en la semana 9 también se retardó la llegada del 50% de la cosecha, siendo diferente estadísticamente a los otros tratamientos de esa semana (0 y 0,7 minadores/hoja) (Fig. 4).
El comportamiento de la cosecha en el transcurso del tiempo muestra el mecanismo de compensación de las plantas atacadas por las poblaciones de minador en la 1a. semana, cuando las plantas posiblemente intentaron incrementar su área foliar o la tasa de fotosíntesis de las hojas que permanecieron, pero la disminución de la fotosíntesis fue tan drástica que no alcanzaron a producir los asimilados suficientes en el tiempo determinado, siendo necesario que se mantuvieran vegetativas dos a tres semanas más para poder florecer.
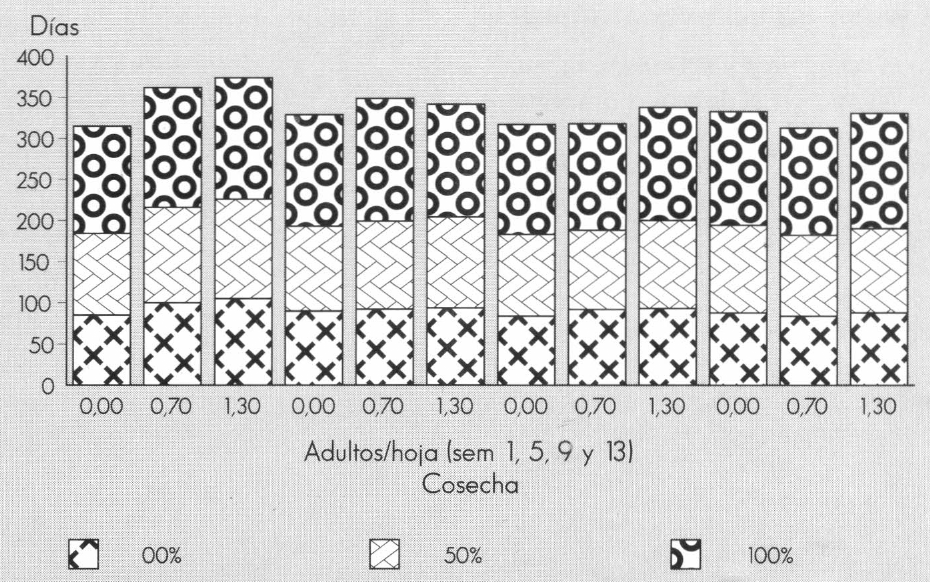
Comportamiento de la producción de Gypsophila paniculata L. al cumplir 0% (comienzo de cosecha), 50% y 100% (fin de la cosecha), de acuerdo a las densidades de infestación del minador.
Número de tallos basales por planta
En esta variable no se encontraron diferencias estadísticas entre los tratamientos (Tabla 3), por lo que, posiblemente, con el daño por el minador sucedió lo mismo que con el daño por defoliación: no se presentó rompimiento de la dominancia apical como mecanismo de respuesta al ataque del insecto.
Biomasa
La biomasa de tallos no se afectó con la presencia de minador en la semana 1. Para la semana 5, la densidad de 0,7 adultos/hoja redujo significativamente la variable (47,8 g), con respecto a las densidades de 0 y 1,3 adultos/hoja (61,2 y 61,0 g, respectivamente). En la semana 9, cualquier presencia del minador (0,7 y 1,3 adultos/hoja) redujo la biomasa de tallos a 47,9 y 53,0 g, respectivamente, con respecto al testigo (68,1 g). En la semana 13 no se afectó esta variable con ningún tratamiento (Fig. 5).
La biomasa de hojas en la semana 1 se redujo, pero no significativamente, con cualquier presencia del minador. En la semana 5 se incrementó la biomasa de hojas con 1,3 adultos/hoja (36,9 g), con respecto al valor obtenido cuando las plantas se infestaron con 0,7 adultos/hoja (25,8 g), mientras que el testigo (29,5 g) fue similar a ambos. Para la semana 9, cualquier densidad del minador redujo la biomasa de hojas, encontrando diferencias estadísticas entre el testigo (38,23 g) y el valor con 1,3 adultos/hoja (28,24 g). En la semana 13, el daño causado por cualquier población del minador no afectó la biomasa de hojas.
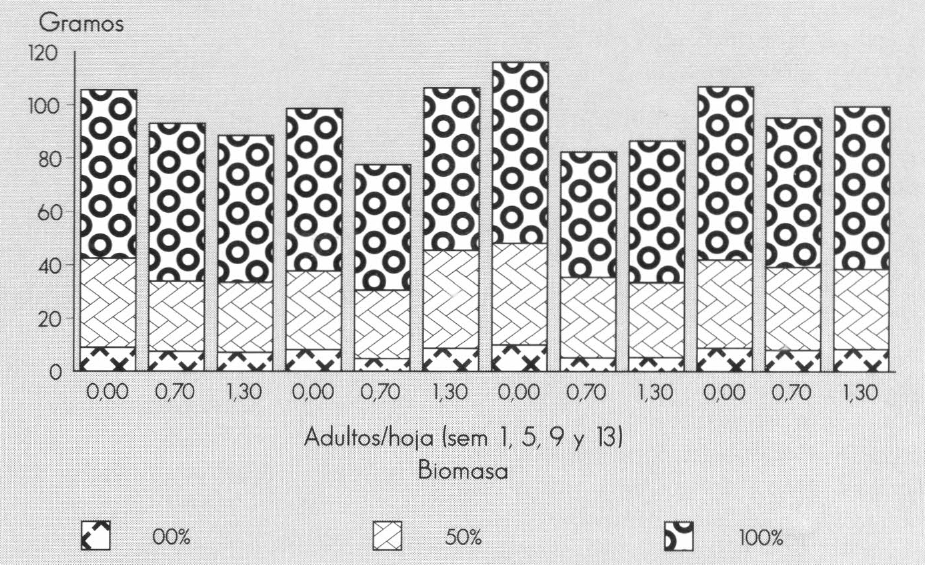
Comportamiento promedio de la biomasa de estructuras (tallos, hojas y flores) por planta de Gypsophila paniculata L. de acuerdo a las densidades de infestación de minador.
La biomasa de flores es importante como parámetro de calidad, ya que a mayor biomasa de flores mayor cantidad de ellas que permiten ramos más llenos y coloridos. En la semana 1, esta variable no se afectó con los tratamientos. En la semana 5 se redujo la biomasa de flores con la población de 0,7 adultos/hoja (4,8 g) con respecto al testigo (8,2 g) y al tratamiento de 1,3 adultos/hoja (8,7 g). En la semana 9, la presencia del minador redujo la biomasa de flores, de 10 g (testigo), a 5,2 y 5,3 g con 0,7 y 1,3 adultos/ hoja, respectivamente. Finalmente, los tratamientos en la semana 13 no afectaron la biomasa de flores con respecto al testigo.
La reducción de la biomasa total, de tallos, de hojas y de flores, en las plantas infestadas la semana 9, mostró su susceptibilidad en esa etapa fisiológica, la cual es de traslocación y acumulación de nutrimentos luego de la inducción floral. La afección de las hojas pudo ser crítica, deteniendo su actividad fotosintética, con lo cual, posteriormente, no alcanzó a recuperarse para permitir una adecuada traslocación de asimilados hacia los tallos y flores.
Conclusiones
Daño por defoliación
La defoliación en la semana 1 no afectó la producción pero retardó la fase de cosecha en 2, 7 y 12 días con 25, 50 y 75% de área foliar perdida.
En la semana 5 disminuyó, no significativamente, la producción en 10,3 у 13,7% con el 25 y 75% de defoliación. El peso del material no comercial se incrementó en 35,6, 28,7 y 30,5% con 25, 50 y 75% de defoliación, respectivamente, y la fase de cosecha se retardó en 6 días con el 75% de defoliación.
Para la semana 9, la producción de las plantas defoliadas el 75% se redujo en 22%; el peso del material no comercial se incrementó en 66,4 y 42,5% con 50 y 75% de defoliación, y la fase de cosecha no se afectó.
En semana 13 se incrementó la producción en 13,8% con el 25% de defoliación. El peso de material no comercial y la fase de cosecha no se afectaron; la biomasa de tallos se incrementó en 32,6% con el 25% de corte del área foliar, así como también la biomasa de flores en 21,1 y 22% con 25 y 50% de defoliación.
Daño de minador
En la semana 1, la producción se redujo 37,3 y 33% con poblaciones de 0,7 y 1,3 adultos de minador por hoja. El peso del material no comercial se redujo en 41,6 y 45,8%, y la fase de cosecha se retardó en 15 y 20 días con los mismos tratamientos.
La infestación en la semana 5 disminuyó la producción en 38 y 27,8% con densidades de 0,7 y 1,3 minadores por hoja; el peso de material no comercial no se afectó, pero se retardó la fase de cosecha entre 2 y 4 días. La biomasa total, de tallos y de flores, se redujo 20,7, 21,9 y 12,6%, respectivamente, con 0,7 minadores/ hoja.
En la semana 9 se redujo la producción en 34,4 y 46,7%; el material no comercial se disminuyó en 45,1 y 30,3%; la fase de cosecha se retardó en 8 y 9 días, con 0,7 y 1,3 adultos/ hoja, respectivamente. La biomasa total se redujo en 28,45 y 25,7%, la biomasa de tallos en 29,7 y 22,2%, la biomasa de hojas en 21 y 26,1% у la biomasa de flores en 48,2 y 47,2% con las mismas densidades, con respecto al testigo.
Al infestar con minador en la semana 13, no se afectó ninguna variable medida.
El número de tallos basales no se afectó con ninguna densidad de minador en cualquier edad del cultivo.
La tasa de producción y crecimiento de hojas a partir de la semana 1 y durante tres semanas, fue similar a la tasa de pérdida de área foliar realizada por las larvas del minador; a partir de las demás semanas de infestación (5, 9 y 13), el incremento del área foliar perdida causada por la actividad del minador, fue mayor.
Recomendaciones
La defoliación puede ser una práctica de manejo del minador, ya que si se quitan las hojas afectadas por el insecto, las pérdidas en producción pueden llegar a ser menores que si el minador permanece en el cultivo, siendo factible reducir la presión química.
En las semanas 1, 5 y 9 es imprescindible cuidar el área foliar producida, aunque en la primera semana se pueda soportar una pérdida de follaje hasta del 49% por punturas y 26% por minas sin una reducción considerable de la producción.