
Abstract
The finding of an insect, responsible for the drop of tomato flower buds, induced basic studies under field and laboratory conditions, where the development of the host and the insect life cycle were followed simultaneously. In order to determine the number of instars, weekly, 100 flower buds were analyzed. Percent parasitism was appraised during the reproductive cycle of tomato, collecting weekly, during one hour, affected buds. Economic aspects were estimated analyzing attack and losses caused by the pest on 33 plants. The insect detected belongs to the new specie Melanagromyza socolena Sanabria de Arévalo y Zenner de Polanía (Diptera: Agromyzidae), which passes its cycle within the flower bud. The adult, a black colored fly with metallic dark green sheen, places one egg within the bud; the egg is white and lasts from 2 - 3 days. The larva, also white, goes through three instars within 5 - 7 days, feeding on anthers, pistil and ovary. The pupa develops between the sepals at the base of the bud, having a mean duration of 11 days. In the moment of pupal formation, the flower bud separates at the abscision point. As parasites, the pteromalids Heteroschema rugosopunctata (Ashmead) and Syntomopus sp., and a genus near Zaeucoila (Eucoilidae) were detected, and toghether cause up to 54.76% parasitism. The bud drop increases as the flowering period advances. In the inferior third of the tomate plant an average of 4.03% of buds is lost, in the middle third 19,88% and in the superior third 28,60%, which corresponds to an average loss of 0.733 kg of possible fruits per plant. Per hectare the losses may reach and exceed 15 tons. Up to now the insect has been detected at altitudes between 1,400 and 1,800 masl in some tomato producing towns of Cundinamarca, but its presence is suspected in all the department.
Introducción
El tomate de huerta (Lycopersicon esculentum Miller) es la hortaliza más popular en Colombia, existiendo una alta demanda para consumo fresco y para procesamiento. La producción para consumo fresco se concentra en la zona cafetera, y en ésta se destaca, por el área cultivada y la productividad, el departamento de Cundinamarca. Los municipios más productores son Fusagasugá, Arbeláez, Anolaima, Cachipay y San Francisco, los cuales abastecen el mercado de Santafé de Bogotá (FEDERACAFE s.f.). Fue precisamente en el último municipio, donde al visitar en 1989 cultivos de tomate se observó una considerable caída de botones florales, no atribuible a causas conocidas. Se logró establecer que cada botón caído contenía en la base, entre los sépalos, la pupa de un díptero. Bajo condiciones de laboratorio del C.I. "Tibaitatá" del ICA, en Mosquera (Cund.), emergieron los adultos que fueron determinados como pertenecientes a la familia Agromyzidae, género Melanagromyza. La revisión de la literatura pertinente, tanto sobre plagas insectiles del tomate como sobre el género Melanagromyza (FEDERACAFE s. f.; Bustillo y Gutiérrez, s.f.; Saldarriaga et al. 1987; García 1988; Spencer 1973, 1973a; Steyskal 1972; Spencer y Steyskal 1986; Spencer y Stegmaier 1973) indicó el hallazgo de una nueva plaga y de una nueva especie para la ciencia, lo cual indujo a los estudios, cuyos resultados se presentan en este trabajo.
Materiales y Métodos
Los estudios de campo se realizaron durante tres cosechas de tomate, variedad "Santa Cruz", tipo Chonto, en una finca de Anolaima (Cund.), a 1.550 msnm y una temperatura promedia de 23°C. Los lotes cubrían un área de 2.500 m2 y en ellos se llevaron a cabo todas las prácticas agronómicas recomendadas para el buen desarrollo del cultivo, incluyendo riego si era requerido. Las enfermedades se controlaron químicamente pero no se realizaron aplicaciones de insecticidas. Las observaciones sobre biología y enemigos naturales y la descripción de los estados de desarrollo del insecto se llevaron a cabo en el laboratorio de Entomología y en un cuarto de cría (T = 24 ± 2°C; H.R. = 70-80%) en el C.I. "Tibaitata" del ICA, en Mosquera (Cund.). Para las mediciones se empleó un estereoscopio con micrómetro incorporado.
Para obtener los datos sobre el ciclo de vida se realizó el seguimiento simultáneo del desarrollo de los botones florales y del insecto, combinando observaciones de campo con trabajos de laboratorio. Con el mismo fin se recolectaron semanalmente 100 botones florales de edad y tamaño específico, los cuales se midieron y abrieron para determinar también el número de instares larvales y hábitos alimenticios de la larva. Además se recolectaron, durante el desarrollo del cultivo, 3.351 botones afectados y ya desprendidos, los cuales se llevaron al laboratorio para determinar la existencia de parasitoides y evaluar el porcentaje de parasitismo. Para la recolección de estos botones se utilizó el método hora/hombre.
Para evaluar las pérdidas por el ataque del insecto se marcaron 33 plantas de tomate en el momento del transplante y en cada una se colocaron tiquetes en cada racimo floral en el momento de su formación. Semanalmente se contabilizaron, en cada racimo, los botones caídos por ataque de la plaga, hasta finalizar la cosecha de los racimos marcados. Para fines de análisis, la planta se dividió en tercios; los datos de botones caídos se transformaron a porcentaje y éstos se agruparon en orden descendente por planta para poder graficar el daño causado a cada una de ellas.
Resultados y Discusión
Melanagromyza socolena Sanabria de Arévalo y Zenner de Polanía es un insecto cuyos hábitos de ataque y alimentación no pueden compararse con otras especies de este género, ya que la larva ataca internamente el botón floral del tomate y no actúa como minador de tallo como lo hacen la mayoría de las especies de Melanagromyza. Aunque existen especies que se alimentan de semillas dentro de las vainas de algunas leguminosas, no se tiene indicio de alguna especie que afecte los botones florales (Spencer 1973a). Esta nueva especie se describió en honor de los 20 años de la Sociedad Colombiana de Entomología, SOCOLEN (Sanabria de Arévalo y Zenner de Polanía 1994). Hasta ahora, el insecto se ha detectado en los municipios de Anolaima, Arbeláez, Fusagasugá, Cachipay y San Francisco, en Cundinamarca, pero se sospecha que se encuentra presente en todo el departamento, cuya área cultivada en tomate es de aproximadamente 2.200 ha.
Biología
Bajo condiciones de campo, el ataque del insecto sólo se nota cuando el botón floral cae al suelo y en el racimo floral se observan los muñones (Fig. 1A); en este momento, la plaga ya se ha desarrollado hasta pupa dentro del botón. El adulto, una mosquita de color negro con visos metálicos verde oscuros y de 2 mm de longitud (Fig. 1B), coloca un solo huevo dentro del cono estaminal del botón floral, cuando éste apenas mide 6,5 a 8,8 mm de longitud. El huevo que queda atravesando una antera y su extremo anterior protrude dentro del cono vacío (Fig. 1C), es de color blanco, con el extremo posterior redondeado; se estrecha hacia el extremo anterior y termina ligeramente curvado en un proceso elongado. Mide, en promedio, 0,4043 mm de largo. Bajo condiciones de campo no se logran distinguir botones con o sin huevos. Bajo microscopio se observa ocasionalmente un tenue halo necrótico en el sitio de oviposición, el cual siempre se encuentra en el tercio superior del botón floral. Este estado del insecto dura en pomedio 2,5 ± 0,5 días.
La larva, también de color blanco, tiene la forma típica de los agromízidos y pasa por tres instares. El primer instar penetra a la antera y consume los granos de polen en formación. A medida que se va desarrollando, se acerca a la base del botón, "minando" dentro de la antera. El segundo instar pasa a otra antera y luego sale para consumir parte del pistilo (Fig. 1D). El tercer instar que se caracteriza por poseer los espiráculos protorácicos de color negro mientras que los caudales son blancos, termina de comer la base del pistilo, penetra al ovario y lo destruye. Las medidas de los tres instares larvales se consignan en la Tabla 1, donde se observa que el ancho, tanto caudal como cefálico, es muy constante dentro del instar, mientras que la longitud de la larva muestra una variabilidad considerable. Mientras la larva cumple su ciclo dentro del botón floral, éste sigue creciendo normalmente y no se nota ninguna diferencia externa con un botón floral no atacado. Así, al alcanzar el botón un promedio de 1,7 cm de longitud y ya los pétalos amarillos que sobresalen, la larva está completamente desarrollada. La duración de este estado es corto, sólo se demora 6 ± 1 días.
La pupa se forma casi siempre entre los sépalos, en la base del botón, dentro de los restos del ovario. Ocasionalmente, la larva antes de empupar sube nuevamente entre los pétalos y empupa en cualquier parte del botón (Fig. 1E). Los espiráculos anteriores siempre sobresalen del botón floral. La pupa tiene forma de tonel, inicialmente es de color blanco cremoso, luego se torna café oscuro. Se caracteriza porque tanto los espiráculos caudales como los anteriores son prominentes y fácilmente visibles (Fig. 1F). Las medidas son bastante constantes como se puede observar en la Tabla 1. La duración de este estado es la más larga, su desarrollo se demora 11±1 días.
Cuando el insecto está próximo a empupar o la pupa ya está formada, pero casi siempre todavía de color claro, se desprende el botón floral en el punto de abscisión y es sólo en este instante que se puede observar directamente el ataque de M. socolena en el campo. Por su daño y la forma de ataque se propone como nombre vulgar del insecto el de "capador del tomate".

Aspectos de Melanagromyza socolena en tomate. A. Daño, B. Adulto, C. Huevo en antera, D. Larva de II instar y restos del botón atacado, E. Pupa sobresaliendo del botón floral, F. Pupa. Nótense los espiráculos.
Dimensiones de la larva y de la pupa de Melanagromyza socolena
En la Figura 2 se relaciona el ciclo de la plaga con el desarrollo del botón floral del tomate. Se nota que existe una perfecta sincronización entre el insecto y el huésped. Este último no se afecta en su crecimiento hasta que se forma la pupa, o sea cuando la plaga ya no necesita tejido vivo y en crecimiento para su desarrollo. La pupa permanece dentro de los restos del botón sobre el suelo, y el adulto emerge aunque los sépalos y pétalos se hayan secado por completo o estén afectados por sequía, exceso de humedad u hongos.
Al correlacionar el ciclo de vida del insecto con el desarrollo de la planta de tomate se obtuvieron los resultados que se indican en la Figura 3. Bajo las condiciones climáticas de Anolaima, a los 17 días, en promedio, después del trasplante ya se han formado los primeros botones florales, llegan los primeros adultos de la plaga y se inicia la oviposición. Esto posteriormente se refleja en la caída de los botones del tercio inferior de la planta. La infestación inicial es baja, pero dada la disponibilidad continua y cada vez mayor de botones para la oviposición y el corto ciclo de vida del insecto, la población aumenta a medida que avanza el desarrollo del cultivo, como se verá más adelante en detalle, al discutir las pérdidas de botones causadas por el insecto a través del tiempo. Al finalizar la cosecha ya han pasado cuatro generaciones completas de la plaga y todavía hay un sinnúmero de botones en formación en el extremo del tercio superior, aptos para ser infestados. De este hecho se desprende la importancia de una total destrucción de los residuos de cosecha, único medio para cortar el ciclo de vida del insecto, ya que aparentemente el tomate es el único huésped.
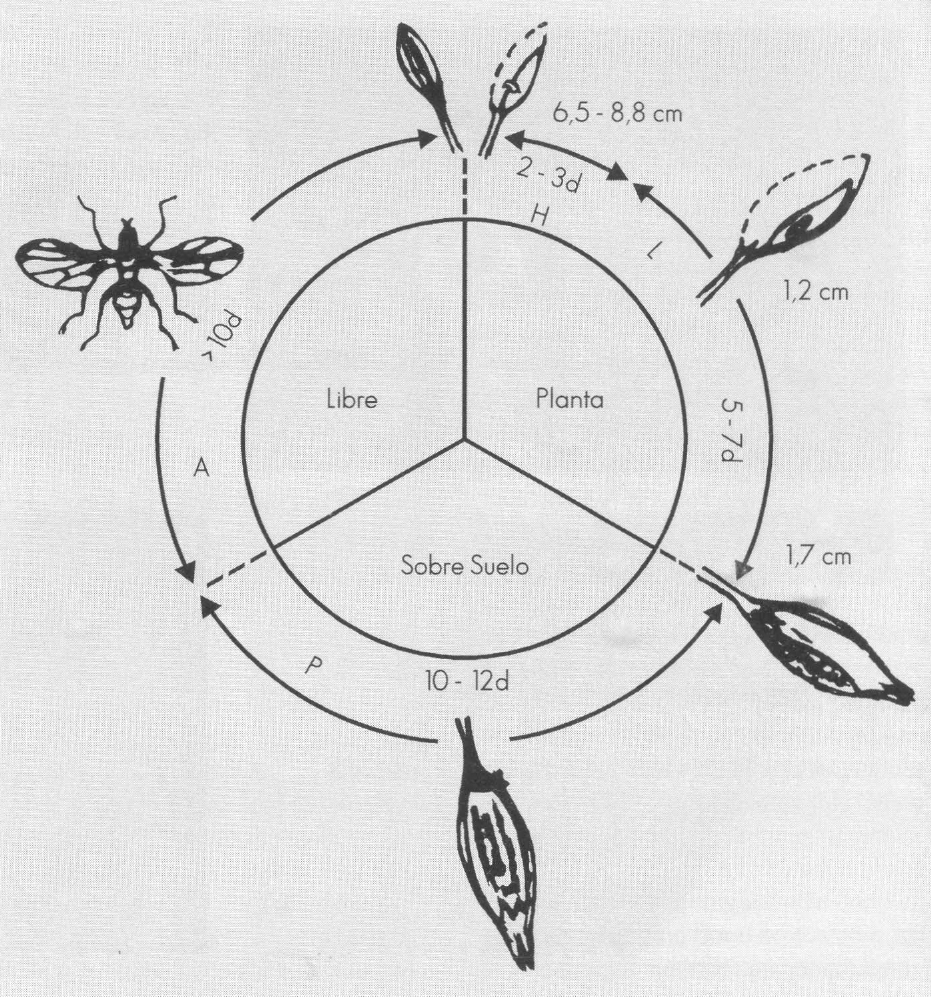
Relación Melanagromyza socolena botón floral del tomate.
Enemigos naturales y Parasitismo
Como parasitoides de M. socolena se han encontrado hasta el momento, en Anolaima, dos especies de la familia Pteromalidae (Hymenoptera): Heteroschema rugosopunctata (Ashmead) y Syntomopus sp., ambos determinados por el Dr. Eric Grissell del SEL-PSI-USDA en Washington, y una de un género cercano a Zaeucoila (Hymenoptera: Eucoilidae), determinado por el Dr. Arnold Menke de la misma institución. Además se han observado ocasionalmente obreras de la hormiga Solenopsis geminata (Fabricius) (Hymenoptera: Formicidae) cargando pupas de la plaga.
H. rugosopunctata fue descrito de especímenes colectados en las islas de San Vicente y Grenada. Es sinonimia de H. prima Gahan, especie que fue criada de M. gibsoni (Malloch) en Tempe, Arizona; otros registros se tienen de Santiago de las Vegas, Cuba (Gahan 1938). El género Syntomopus fue eregido por Walker en 1933, y la especie S. americanus Ashmead se conoce como parasitoide de M. angelica (Frost) y M. virens (Loew) (Muesebeck et al. 1951). Esta última especie es plaga de la compuesta Parthenium argentatum Gray y fue estudiada por Lange (1944), quien registra al parasitoide, pero no menciona el estado del huésped afectado. Syntomopus sp. es el único de los tres enemigos naturales registrados en este trabajo que se crió a partir de muestras de botones afectados procedentes de Arbeláez, y recolectadas en campos de tomate con innumerables aplicaciones de insecticidas. Del tercer parasitoide, cerca a Zaeucoila, solamente se lograron obtener seis especímenes en enero de 1993. No se encontró literatura sobre el género, pero Borror et al. (1976) mencionan que los géneros de la subfamilia Eucoilinae (Cynipidae) son parasitoides de pupas de varios dípteros.
Los porcentajes de parasitismo ejercidos por la sumatoria de H. rugosopunctata y Syntomopus sp., obtenidos desde el inicio hasta el final de la cosecha de tomate, durante 78 días, se ilustra en la Figura 4. A partir del 18 de noviembre y con el fin de determinar el estado del huésped parasitado, se discriminó durante la recolección entre botones florales afectados "frescos" y "viejos". Los primeros hacen referencia a aquellos que todavía se encuentran ligeramente adheridos al racimo, pero se desprenden al tocarlos. En este momento, la mayoría de los individuos del insecto se encuentran como prepupa o pupa de color blanco cremoso. Los botones viejos son aquellos botones caídos y recolectados bien sea del follaje o del suelo. Por la marcada diferencia entre los porcentajes de parasitismo entre botones frescos y viejos se deduce que ambos pteromálidos son parasitoides de pupa - pupa. De la gráfica también se desprende que el parasitismo aumenta a medida que se incrementa la población de la plaga, pues se observaron todos los botones recolectados durante una hora cada semana. No se detectaron parasitoides durante la primera generación de la plaga (Fig. 3) y sólo se obtuvieron los primeros al finalizar la segunda generación.
La proporción entre H. rugosopunctata y Syntomopus fue de 1:1,34. Es factible que ambos benéficos tengan en el área otros huéspedes y que el rango de S. sp., la especie predominante, sea mayor.
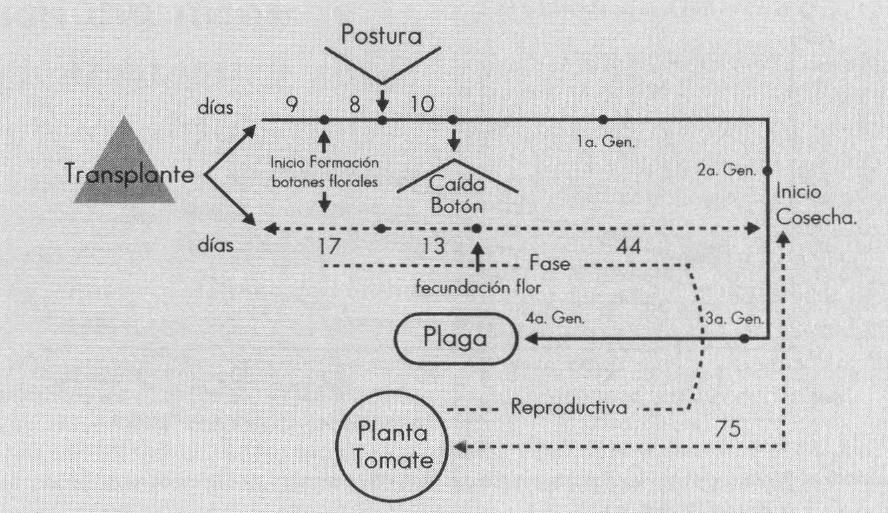
Comparación entre la fenología reproductiva del tomate y las generaciones de Melanagromyza socolena.
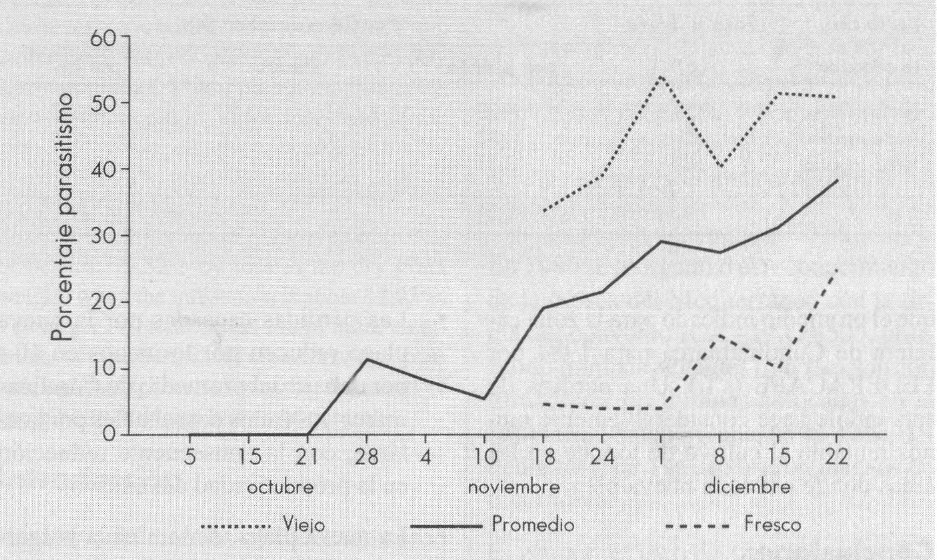
Porcentaje de parasitismo ejercido por Syntomopus sp. y H. rugosopunctata durante tres generaciones de Melanagromyza socolena
Aspectos económicos
Teóricamente, el botón floral que se cae por ataque de la plaga corresponde a un fruto que no se cosecha, lo que automáticamente se traduce en pérdidas para el agricultor. En la variead "Santa Cruz", el promedio de botones florales observado por racimo es de 6 y la caída, a causa del insecto, oscila entre 0 y 100%. Este daño se observó tanto bajo condiciones de no aplicación de insecticidas en Anolaima, como en lotes de Arbeláez, donde el cultivo ya había recibido 18 aplicaciones de una amplia gama de productos.
En la Figura 5 se muestra la caída de botones contabilizados durante el ciclo reproductivo de 33 plantas de tomate. En el tercio inferior de la planta cae, en promedio, el 4,03% de los botones, lo cual se puede considerar bajo y corresponde al ataque de la primera generación de la plaga. El tercio medio sufre la pérdida promedia de 19,88% y en el tercio superior, la caída alcanza, en promedio, el 28,60%. Desde el punto de vista económico, la pérdida más importante corresponde al tercio medio, ya que en él se cosecha el 50% de la producción total de tomate (FEDERACAFE s.f.). El ataque al tercio superior, aunque muy alto, representa una disminución menor de los ingresos, ya que los frutos cosechados en él son de tamaño pequeño y se venden como "parejo" a precios bajos. El ataque en este tercio es, sin embargo, de gran importancia por representar el foco de infección para cultivos vecinos o para la próxima cosecha.
En la Tabla 2 se hace un análisis de las pérdidas en peso por el ataque del insecto. Como base se tomaron 20.750 plantas de tomate por hectárea. Así, los frutos que se dejaron de cosechar por ataque de la nueva plaga puedan alcanzar y sobrepasar las 15 toneladas de tomate, las cuales, con un valor promedio de $1.500 por caja de 10 kg en las plazas municipales, corresponden a aproximadamente $2′250.000. En las tres cosechas de la hortaliza, en los lotes experimentales, se obtuvo un promedio de 20 t/ha, lo cual corresponde al promedio nacional, pero éste es 10 toneladas más bajo que el promedio indicado para la zona cafetera de Cundinamarca para 1984 por FEDERACAFE (s.f.). Una pérdida de esta índole hace considerablemente menos rentable el cultivo de tomate en las áreas donde existe la nueva plaga.
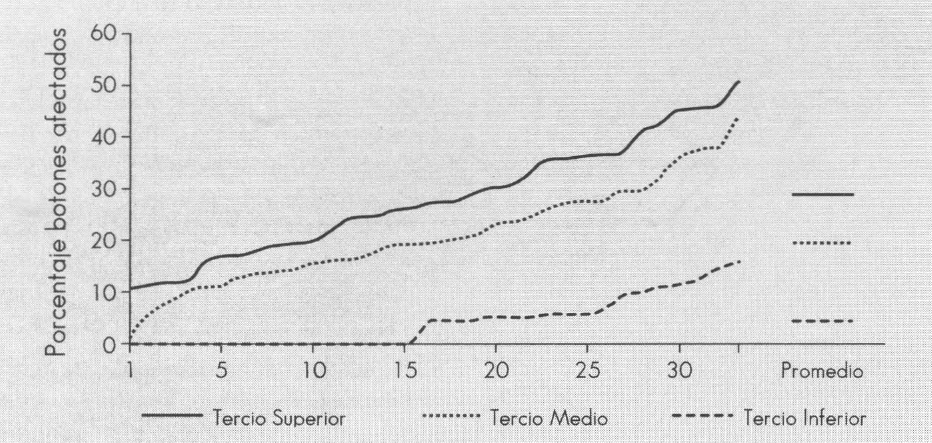
Pérdida de botones florales por ataque de Melanagromyza socolena por tercio de la planta de tomate.
Peso de frutos perdidos a causa del ataque de Melanagromyza socolena
Conclusiones
La plaga de tomate encontrada es una especie nueva para la ciencia, con hábitos desconocidos para una especie de Melanagromyza y fue descrita como M. socolena. Aparentemente tiene al tomate como único huésped.
La población, el daño y los enemigos naturales aumentan a medida que avanza el desarrollo del cultivo, para presentarse el máximo ataque y parasitismo a finales de la cosecha.
Como posibles medidas de manejo de la plaga se vislumbra el control biológico por Syntomopus sp. y Heteroschema rugosopunctata, el trampeo de adultos y la destrucción rigurosa de los residuos de cosecha.
Las pérdidas causadas por la nueva plaga reducen por lo menos en 10 t por debajo del promedio de Cundinamarca, los frutos cosechables por hectárea, con la consecuente reducción en la productividad del cultivo.
La nueva plaga, denominada vulgarmente "capador del tomate", trastorna cualquier programa establecido de manejo integrado de plagas insectiles de esta hortaliza.
Footnotes
Agradecimientos
A Fabio Andrés y Paula Polanía, sin cuya paciencia y colaboración hubiera sido imposible realizar este estudio.