
Abstract
Biological aspects of the tomato leafminer Liriomyza sativae Blanchard (Diptera: Agromyzidae) were studied under laboratory conditions (T=26°C; R.H=67%) during the first semester of 1992. Mean duration of the different developmental stages was: egg 3.5 days; larva 7.0 days; prepupa 4.5 hours and pupa 8.0 days. Mean duration of the life cycle from egg to adult was 18.5 days. Adults males and females fed with 5% honey water solution lasted 6.1 and 10.0 days, respectively. Preoviposition and ovoposition periods were 1.0 and 7.5 days. Fecundity was 24.4 eggs/female, where as fertility was 59%. Three larval instars were registered using Dyar's law and based on the size of the mouth hooks. Sex ratio was 1 male: 1.77 female.
Introducción
El pasador de la hoja de tomate, Liriomyza sativae Blanchard (Diptera: Agromyzidae), constituye uno de los problemas entomológicos más importantes del tomate (Lycopersicon esculentum Mill.) en Venezuela, especialmente en las zonas donde se cultiva durante todo el año, como en el Valle de Quíbor, municipio Jiménez, Estado Lara. Spencer (1973) registra esta especie en Venezuela e indica que presenta altas poblaciones, es polífaga y constituye un serio problema para los agricultores, en una amplia variedad de cultivos.
Su importancia como plaga ha aumentado como consecuencia de las aplicaciones frecuentes y en dosis elevadas de insecticidas organosintéticos, como única medida de control, lo cual ha provocado una reducción drástica en las poblaciones de sus enemigos naturales, situación que ha contribuido a incrementar las pérdidas en las cosechas, a reducir el área sembrada y aumentar los costos del control. Spencer (1973) reporta poblaciones de L. sativae en áreas agrícolas importantes de los estados Aragua y Lara, e indica que las aplicaciones de insecticidas, en la mayoría de los casos, afecta más a los parasitoides que a las larvas y adultos de L. sativae.
Varios estudios se han realizado sobre la biología de especies del género Liriomyza (Hendrickson y Keller 1983; Mora y Mosquera 1984; Schuster y Patel 1985; Parella 1987; Salas et al. 1988), pero pocos realizados con L. sativae, Oatman y Michelbacher (1958) en melón, Parkman et al. 1989 en tártago e Issa y Marcano (1991) en tomate.
El presente estudio pretende contribuir a un mejor entendimiento de la biología de L. sativae y sus relaciones con el cultivo y el ambiente ocupado, para en esa forma diseñar estudios para la generación de alternativas de control dentro del enfoque de Manejo Integrado de Plagas (MIP).
Materiales y Métodos
El estudio sobre la biología del pasador de la hoja del tomate L. sativae se realizó, a nivel de laboratorio, en la Estación Experimental Lara del Fondo Nacional de Investigaciones Agropecuarias (FONAIAP), ubicada en el municipio Iribarren del Estado Lara, Venezuela, bajo condiciones promedio de temperatura y humedad relativa de 26°C y 67%, respectivamente, durante el primer semestre de 1992.
El material biológico inicial se colectó en hojas de tomate provenientes de siembras localizadas en zonas productoras de Quíbor, municipio Jiménez del Estado Lara. Las hojas infestadas con larvas se colocaron en frascos de vidrio de boca ancha, a los cuales se les colocó un pedazo de tela organdi sujetada con una banda de goma, para evitar el escape de los adultos. Se seleccionaron parejas con machos y hembras recien emergidos de la pupa y se hicieron parejas y éstas se colocaron en frascos de vidrio de boca ancha con pecíolos jóvenes de tomate mantenidos dentro de un frasco pequeño con agua y sujetando con goma de esponja. Los pecíolos se usaron como sitio para la ovoposición, asegurándose de que no tuvieran huevos ni otros estados de desarrollo de insectos.
Para la observación de la ovoposición se utilizaron 80 parejas, colocando dos por frasco, los cuales se taparon con tela ordandí sujetada con una banda de goma. Una vez ocurrida la oviposición, los pecíolos se sacaron de los frascos y se transfirieron a otros iguales, para posteriormente hacerles un seguimiento individual diario a 80 huevos seleccionados, utilizando un huevo por folíolo y así determinar el período de incubación.
Para el estudio de la duración del estado larval se utilizaron 100 larvas individuales recien nacidas, observándolas hasta el momento que salían de la mina o permanecían en ella para transformarse en prepupa.
Se seleccionaron 100 prepupas y se colocaron en cajas de Petri, provistas con papel toalla ligeramente humedecido, para observar la formación de la pupa y registrar la duración de este estado. En las mismas condiciones se escogieron 100 pupas recién formadas, se observaron hasta la emergencia del adulto y se registró la duración de este estado. En cada pupa se midió la longitud y la anchura, utilizando una reglilla micrométrica adaptada a un microscopio estereoscópico con un aumento de 40X, y posteriormente se pesó en una balanza analítica. Un grupo de pupas se colocó en cajas de Petri y a su vez en una cámara húmeda, separando dos subgrupos, uno colocado todo el día en la oscuridad y el otro bajo luz artificial, para observar el posible efecto de estos factores en la duración de este estado. La longevidad se estudió empleando 25 hembras y 25 machos recién emergidos, los cuales se colocaron separadamente dentro de frascos de boca ancha cubiertos con un pedazo de tela organdí sujetada con una banda de goma y se alimentaron con una solución acuosa de miel de abejas al 5%. Para la determinación de la fecundidad, se utilizaron 10 parejas recién emergidas, colocando cada una en la unidad de ovoposición, y diariamente se registró el número de huevos depositados por cada hembra, así como la duración de los períodos de preovoposición y ovoposición. La separación de sexos se realizó a nivel de adulto, ya que las pupas resultaron muy similares en cuanto al peso y tamaño, sin presentar evidencias características para la separación de sexos. A los adultos separados por sexo se les midió la longitud y anchura en la parte media del tórax, y además se registró el color del cuerpo, ojos, antenas y alas.
En cuanto a la fertilidad de los huevos, se seleccionaron 80, se registró el número de eclosionados y se consideró como infértiles aquellos que no lograron eclosionar.
Para determinar el número de instares larvales, se tomaron 20 parejas y cada una se colocó en un frasco para que ovipositaran. Una vez se obtuvieron larvas, diariamente se seleccionaron entre 25 y 40 y se les midió la longitud de los ganchos bucales, utilizando una reglilla micrométrica adaptada a un microscopio estereos cópico.
Resultados y Discusión
Descripción de los Estados de Desarrollo
Durante la fase de prepupa, las larvas se contraen y toman una forma elipsoidal, se acortan y cesan todos los movimientos para luego empupar y presentan un color amarillo naranja.
Duración de los Estados
En la Tabla 1 se registra la duración promedio de los direrentes estados de desarrollo de L. sativae. Los resultados indican que la duración del huevo es bastante similar a la reportada por Salas et al. (1988) para L. huidobrensis y difieren mayormente con los obtenidos por Issa y Marcano (1991) para la misma especie. Parella (1987) señala que la duración del estado de huevo es variable dependiendo de la especie, la planta hospedante y la temperatura. En cuanto al período larval, ocurre lo mismo, se observa una gran similitud con los resultados de Salas et al. (1988) y una gran diferencia con los de Issa y Marcano (1991). Otros autores reportan una duración entre 4 y 5 días para L. sativae y L. trifolii, señalando que ésta depende de la especie, el alimento y la temperatura (Oatman y Michelbacher 1958; Schuster y Patel 1985). La duración de la pupa es muy similar a la reportada por Oatman y Michelbacher (1958), Salas et al. (1988) e Issa y Marcano (1991), para L. pictella, L. huidobrensis y L. sativae, respectivamente. La duración promedio del ciclo de vida desde huevo a la formación del adulto fue de 18,5 días, la cual difiere con la reportada por Issa y Marcano (1991) que fue de 15,71 días. Las pupas que permanecieron en la oscuridad duraron 5 días menos que las que permanecieron en la luz. En cuanto a longevidad del adulto también se observó una gran difrencia con los resultados de Issa y Marcano (1991), siendo mucho mayor la reportada en este estudio. Parella (1987) señala que la longevidad disminuye con las altas temperaturas y se incrementa notablemente cuando se alimentan con miel de abejas, alimento usado en este estudio.
Duración de las fases de desarrollo de L. sativae a nivel de laboratorio. Estación Experimental Lara. 1992.
horas.
Para determinar el número de instares de L. sativae, se utilizó la ley de Dyar, citada por Gaines y Campbell (1935). Al agrupar las medidas de la longitud de los ganchos bucales en un histograma de frecuencia (Fig. 1) se observan tres grupos discretos que representan tres instares bien definidos, lo cual coincide con los resultados obtenidos por otros investigadores (Salas et al. 1988; Petitt 1990; Issa y Marcano 1991). En la Tabla 2 se presentan las dimensiones de los ganchos bucales de los tres instares larvales y la relación de crecimiento entre instares (r). Como se observa, los valores promedios de la longitud del gancho bucal aumentan a través de los diferentes instares, tomando la forma de una progresión geométrica como lo indica la ley de Dyar. En cuanto a la relación de crecimiento, ésta tiende a disminuir a través del desarrollo larval, coincidiendo con datos similares de varios autores señalados por Gaines y Campell (1935). Se utilizó el modelo y=ax+b donde y= longitud del gancho bucal y x= número de instar. De esta forma se obtuvo la línea de regresión y= 0,098x + 0,01 con r2= 1 (Fig. 2). Esta línea de regresión permite estimar, con un alto valor predictivo, el instar al cual pertenece una larva, una vez conocida la longitud del gancho bucal.
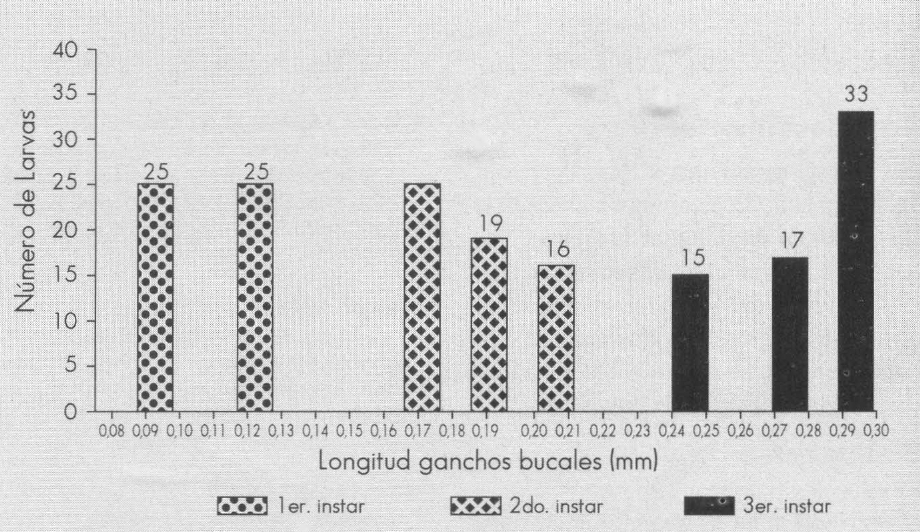
Distribución de frecuencia de las medidas de longitud de los ganchos bucales de L. sativae.
Línea de regresión calculada de la longitud promedio del gancho bucal de Liriomyza sativae.
Longitud promedio (mm) del gancho bucal de larvas de L. sativae y la relación entre el promedio de cada instar y el anterior (r). Estación Experimental Lara. 1992.
Preovoposición y Ovoposición
El período de preovoposición fue de 1 ± 0,01 día (rango de 1-1,05 días), mientras que el de ovoposición fue de 7,5 ± 0,65 días (rango de 1-9,5 días).
Fecundidad y Fertilidad
El promedio de huevos depositados por hembra fue de 24,4 ± 1,42 con un rango de 22 a 26 huevos, mientras que el número de huevos/hembra/día fue de 4,6 ± 1,07. De un total de 80 huevos observados, 47 resultaron ser fértiles, es decir eclosionaron, lo cual representa un 59% de fertilidad. Estos resultados indican una fecundidad mucho mayor que la reportada por Issa y Marcano (1991) para L. sativae alimentado en folíolos de tomate. Oatman y Michelbacher (1958) reportan un promedio de 688/huevos/hembra para L. pictella. La fertilidad en este estudio resultó ser menor, ya que Issa y Marcano (1991) registraron una fertilidad del 69,79%.
Proporción Sexual
La proporción de sexos fue de 1 macho: 1,77 hembras (n:100), resultando ligeramente diferente a los reportados por Oatman y Michelbacher (1958) e Issa y Marcano (1991).
Footnotes
Agradecimientos
Los autores agradecen a R.V. Peterson, del Systematic Entomology Laboratory, del USDA en Washington, D.C. y a I. Sanabria de Arévalo, del Instituto de Ciencias Naturales del Museo de Historia Natural de la Universidad Nacional de Colombia, en Santafé de Bogotá, por la identificación taxonómica de la especie.