
Abstract
Two Beauveria bassiana isolates and eight insecticide formulations were examined in vitro to determine compatible combinations for the Hypothenemus hampei management. Differences were found among the isolates. In addition, the isolates reacted in a different way depending on the insecticide formulation. Diazinon WP, malation and fenitrotion were the most toxic; the other insecticides had variable effects on the isolates. The inhibition effect increased throughout the mixing time. A fungistatic effect of the insecticides on the isolates was observed. The interaction in the spray tank could be avoided by using separate applications of the two products. However, this will represent a higher cost in the borer suppression.
Introducción
La capacidad toxicológica de los productos químicos aplicados en la agricultura para el control de las plagas también afecta otros organismos. Ellos ocasionan el desarrollo de resistencia y resurgencia de insectos plagas, permiten el aumento de las poblaciones de plagas secundarias, eliminan los enemigos naturales, destruyen los insectos polinizadores y afectan la fauna silvestre y los animales domésticos (Vergara 1991).
El hongo entomopatógeno Beauveria bassiana (Bals.) Vuill. (Hyphomycetos) se está usando en el control de la broca del café, Hypothenemus hampei (Ferrari) (Coleoptera: Scolytidae). El uso de mezclas de B. bassiana e insecticidas para el control de esta plaga es una opción atractiva. Muchos estudios han indicado que los insecticidas pueden inhibir la actividad de B. bassiana (Ramarajahe et al. 1967; Olmert y Kenneth 1974; Clark et al. 1982; Osborne y Boucias 1985; Rivera 1992; King et al. 1993), puesto que la exposición prolongada in vitro del hongo a formulaciones de insecticidas es detrimente, la mezcla muestra un proceso que podría ocurrir en el campo.
Algunos investigadores han probado entomopatógenos en mezcla con insecticidas. Gravena et al. (1980) encontraron un mayor control de las plagas lepidópteras del tomate: Phthorimaea operculella (Zeller) (Gelechiidae), Heliothis spp. y Trichoplusia ni (Hubner) (Noctuidae) al combinar la bacteria Bacillus thuringiensis con metomil (0,016 y 0,18 kg i.a./ha, respectivamente) que cuando aplicaron B. thuringiensis solo.
La mortalidad de insectos provocada por los hongos entomopatógenos se incrementa, en general, en presencia de una dosis baja de un insecticida. Pristavko (1966) encontró que la alta eficacia del hongo B. bassiana en asociación con pequeñas dosis de DDT en el control de Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae) parece ser el resultado de una acción conjunta del insecticida DDT (0,00016 mg i.a.), el hongo (5 × 104 conidias) y la microflora intestinal que se combinan en el proceso patológico; además, la variación del pH de la hemolinfa parece ser el factor de debilitamiento fisiológico del insecto que permite el desarrollo de la enfermedad.
En Francia, Fargues (1973) examinó el DDT en combinación con B. bassiana, y halló que el insecticida es un agente que poco favorece las larvas de L. decemlineata a la micosis. En estudios de campo posteriores (Fargues 1975), con los insecticidas azinfosmetil y carbaryl, registró que combinaciones en dosis reducidas de estos insecticidas con B. bassiana no producen acción sinérgica. Durante los primeros cinco días, la efectividad de la bacteria fue similar a la obtenida con dosis reducidas de insecticida solamente; en consecuencia, estos resultados fueron mayores que cuando sólo se utilizó B. bassiana.
Riba et al. (1983) probaron la susceptibilidad de huevos, larvas y pupas de Ostrinia nubilalis (Hubner) (Lepidoptera: Pyralidae) a diferentes cepas de B. bassiana, Metarhizium anisopliae (Metsch.) Sorokin, Nomuraea rileyi (Farlow) Samson, Paecilomyces fumoso-roseus (Wise) Brown & Smith; además, estudiaron la susceptibilidad a la micosis en larvas de quinto instar con la utilización de clorpirifos en mezcla con el hongo, encontrando que con 1 ó 100 ppm, se alcanzaba el 80% de mortalidad, mientras que el testigo sólo alcanzaba el 50%. En relación con el tiempo letal medio (TL50), hallaron que se reducía el 30% (TL50= 11 días con hongo y 7,6 días con el insecticida en mezcla a la dosis de 100 ppm). Respecto a la tasa final de mortalidad total hallaron que oscilaba entre el 90 y 100% en el caso de la mezcla.
Sikura y Sikura (1983) mostraron que Boverin™ en combinación con dosis bajas de fosalone fue altamente efectivo contra el escarabajo de la papa, L. decemlineata.
Anderson y Roberts (1983) probaron seis aislamientos de B. bassiana y 13 formulaciones de insecticidas, en mezcla, para hallar combinaciones de compatibilidad en el manejo de L. decemlineata. Al evaluar la viabilidad del hongo, hallaron poca diferencia entre las respuestas de los diferentes aislamientos a los plaguicidas. Los insecticidas formulados como concentrado emulsionable (CE) que usan solventes aromáticos a base de xileno, fueron más inhibitorios para el hongo. Las formulaciones en polvo mojable a menudo incrementan el conteo de las unidades formadoras de colonias; los piretroides (permetrin y fenalverato) fueron inhibitorios como formulación y como ingrediente activo. En todos los tratamientos, la mayor inhibición ocurrió dentro de las siguientes cuatro horas de la mezcla.
Foschi y Grassi (1985), al probar la efectividad de B. bassiana y M. anisopliae aplicados en forma individual o conjuntamente con dosis bajas de clorpirifos contra O. nubilalis en el campo, hallaron que la adición del insecticida en una proporción de 0,9 kg i.a./ha, incrementaba la mortalidad de las larvas que emergían después del invierno, pero no reducía el número de perforaciones por planta.
Slavchev (1988), en Bulgaria, realizó estudios en el campo con combinaciones de B. bassiana formulado en granulos y las dosis comerciales recomendadas de los insecticidas granulares carbofuran y terbufos, aplicados al suelo para el control de larvas de primero y segundo instar de Bothynoderes punctiventris (Germ) (Coleoptera: Curculionidae), y encontró niveles de efectividad que alcanzaban aproximadamente el 90%. De los resultados se dedujo un efecto sinérgico, puesto que cuando B. bassiana fue aplicado solo, alcanzó una efectividad del 79%. Una adecuada irrigación fue prerrequisito para el desarrollo satisfactorio y efectividad del hongo.
Anderson et al. (1989), al evaluar combinaciones de B. bassiana y formulaciones de los insecitcidas carbaryl, fenvalerato, abamectin (competidor del ácido aminobutírico), triflumuron (benzoilfenilurea que inhibe la síntesis de quítina) y thuringiensin (inhibidor competitivo de ATP en síntesis protéica) en compatibilidad y eficacia para el control de L. decemlineata, in vitro y en el campo, hallaron que las combinaciones del hongo con un insecticida dado, fueron consistentemente más tóxicas que el hongo. Los resultados indicaron que los dos tratamientos tuvieron un efecto aditivo; en el campo, las pruebas con las mezclas mostraron mortalidades mayores que las causadas por los agentes individuales, pero la interación sinérgica no fue aparente.
La urgente necesidad de desarrollar estrategias para un empleo efectivo del Manejo Integrado de la Broca (MIB) han llevado al Laboratorio de Patología de Insectos de CENICAFE a examinar la compatibilidad in vitro de dos aislamientos de B. bassiana en mezcla con varias formulaciones de los insecticidas utilizados en el control químico de la broca del café.
Materiales y Métodos
En este experimento se utilizaron conidias del hongo B. bassiana, de los aislamientos Bb 9002 aislado de H. hampei y Bb 9205 aislado de Diatraea saccharalis (Fabricius) (Lepidoptera: Pyralidae), pertenecientes a la colección del Laboratorio de Patología de Insectos de CENICAFE; las conidias se produjeron en sustrato de arroz cocido, con un tiempo de inoculación de 25 días, a una temperatura promedio de 25 ± 2°C, tiempo en el cual se obtiene la máxima esporulación (Antia y Bustillo 1993). Para la prueba, las conidias se prepararon suspendiéndolas en una solución acuosa del aceite agrícola «Carrier» al 0,1% y su viabilidad se determinó por germinación del 100% a las 48 horas en Agar Sabouraud Dextrosa (ASD) con Extracto de Levadura al 1%, acidificado con ácido láctico al 0,44%.
Se examinó la compatibilidad de B. bassiana, en una concentración de 9,7 × 108 conidias/ml (Antia et al. 1992), en mezcla con tres concentraciones de las formulaciones comerciales de ocho insecticidas probados por CENICAFE para el control químico de la broca: endosulfan (solvente solvesso), clorpirifos, pirimifos-metil, fenitrotion, malation, diazinon (PM y EM) e isazofos. Las suspensiones de los insecticidas se basan en las dosis de aspersión en el campo (DC), recomendadas por los productores para el cultivo del café; también se utilizó la dosis comercial media (1/2 DC) y una dosis menor (DM) para establecer un rango de subdosis (Tabla 1). Todos los tratamientos se compararon con un testigo que consistió en conidias de B. bassiana y agua destilada estéril (ADE) con aceite «Carrier» al 0,1%.
Insecticidas y concentraciones evaluadas en el estudio de compatibilidad en mezcla con B. bassiana
i.a. ingrediente activo
DC dosis comercial
DC1/2 mitad de la dosis comercial
DM dosis menor
CE concentrado emulsionable
PM polvo mojable
EM emulsión mojable
SM solución microencapsulada
Los dos agentes (B. bassiana e insecticida) se mezclaron en matraces, se mantuvieron en agitación (110 rpm) para simular el movimiento del operario en el campo y se evaluó la germinación conidial después de estar en mezcla 1, 3 y 6 horas. Transcurrido cada intervalo de tiempo, se inocularon cajas de petri que contenían SDY, con 10 alícuotas de 5 µl de cada uno de los tratamientos.
El experimento se organizó en un diseño completamente al azar con el matraz como unidad experimental, con 4 repeticiones por tratamiento a un nivel de significancia del 5%. Para evaluar la compatibilidad de los tratamientos con el hongo y la posible fungistasis causada por los insecticidas, se midió la variable germinación de conidias, en porcentaje, a las 24 y 48 horas. Los resultados se analizaron empleando un análisis de varianza y se ajustó una regresión lineal simple a través del tiempo de mezcla con cada producto, ésta tuvo la forma y = a + bx, donde a es el intercepto y b es la pendiente. Con el fin de poder comparar la diferencia entre los productos y dosis utilizados, se calculó el área bajo la curva de la regresión de la germinación en el tiempo, por medio de una integral (Ecuación 1); para tal efecto, se utilizó el procedimiento del modelo lineal general del Sistema de Análisis Estadístico (SAS 1988).
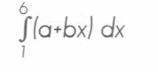
Resultados y Discusión
La dosis en la cual cada uno de los insecticidas inhibió los aislamientos de B. bassiana en los tiempos evaluados se muestra en las Tablas 2 y 3.
Inhibición, en porcentaje promedio, causada por diferentes insecticidas en mezcla con B. bassiana Bb9002, usando tres dosis, tres tiempos de mezcla y dos tiempos de germinación del hongo.
Inhibición, en porcentaje, causada por diferentes insecticidas en mezcla con B. bassiana Bb9205, usando tres dosis, tres tiempos de mezcla y dos tiempos de germinación del hongo.
Al comparar los límites de confianza del 95% (a partir de error estándar = es) de los interceptos del testigo de los dos aislamientos, con los de los interceptos de las mezclas del hongo con la DC de diazinon EM, endosulfan e isazofos, insecticidas que permitieron la germinación a las 24 horas, no se traslapan (Tablas 4 y 5); en teoría esto quiere decir que el efecto inhibitorio de la germinación conidial se produce tempranamente en las mezclas debido al tipo de solvente utilizado en las formulaciones que tienen propiedades desecantes de la membrana citoplasmática, afectando la viabilidad de la conidia. Al observar las pendientes se puede deducir que los valores negativos en la dosis comercial de endosulfan y diazinon EM, en contraste con la pendiente positiva de isazofos, tienen un mayor efecto sobre el hongo, lo mismo se deduce para las otras dosis evaluadas.
Interceptos (a) y pendientes (b) de la regresión de la germinación de B. bassiana Bb9002 a las 24 horas, después de la mezcla a diferentes tiempos, con tres dosis de los insecticidas evaluados.
- no se presentó germinación
Interceptos (a) y pendientes (b) de la regresión de la germinación de B. bassiana Bb9205 a las 24 horas, después de la mezcla a diferentes tiempos, con tres dosis de los insecticidas evaluados.
- no presentó germinación
Al utilizar la prueba de comparación de medias de Tukey, hubo diferencias significativas entre productos con cada uno de los aislamientos (Fig. 1). Se observa que algunas combinaciones de dosis menores que la comercial, no tienen mayor efecto sobre B. bassiana. El uso de estas mezclas puede tener una ventaja en el MIB, ya que ellas introducen multiples factores de mortalidad contra la broca que podrían reducir la dosis a aplicar y tal vez el número de aplicaciones del insecticida, minimizando el peligro de contaminación ambiental; sin embargo, esto implica determinar la participación de cada uno en la mortalidad de la población de broca.
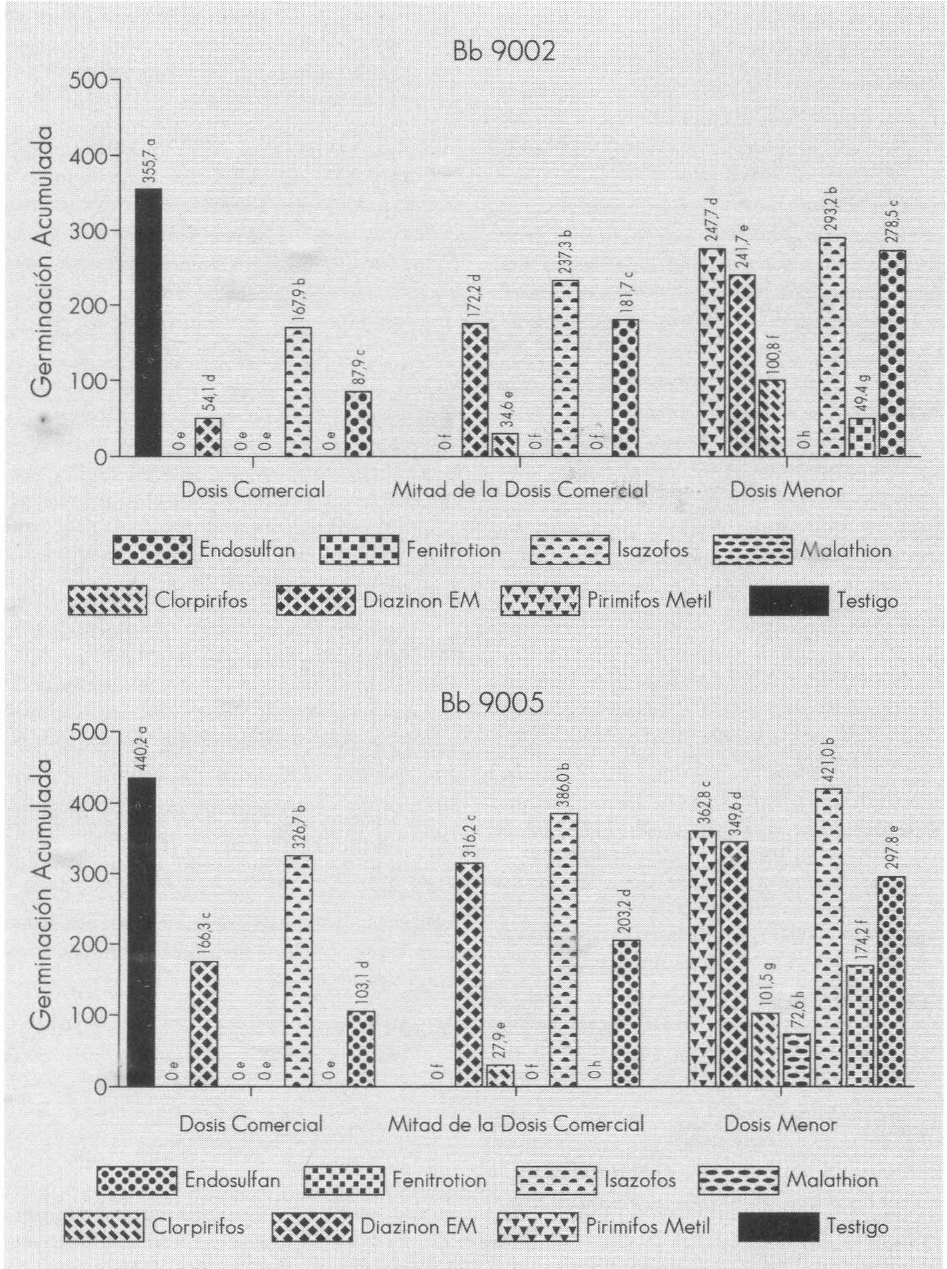
Comparación del área bajo la curva de la germinación de B. bassiana Bb9002 y 9005 (24h), después de la mezcla a diferentes tiempos con tres dosis de insecticidas
Si se compara el área bajo la curva de la germinación acumulada a través del tiempo, la respuesta de los aislamientos frente a los insecticidas fue distinta cuando se evaluaron en SDY (Fig. 1); igualmente, al comparar los límites de los interceptos de las respectivas dosis y productos para cada uno de los aislamientos, se observa que el aislamiento Bb9002 fue más sensible a los insecticidas. Este hallazgo podría implicar que primero se deben seleccionar los aislamientos virulentos de B. bassiana y posteriormente seleccionar por su compatibilidad en mezcla con insecticidas, para así potenciar su acción, asegurando su aprovechamiento en el MIB. El diazinon PM inhibió completamente la germinación en los tiempos evaluados, mientras diazinon EM no; estos resultados están acordes con los hallados por Olmert y Kenneth (1974) y Alves (1986), quienes utilizando una formulación CE no encontraron inhibición causada por el insecticida.
En el presente estudio, el endosulfan tiene como solvente solvesso, y esta formulación no mostró un fuerte efecto sobre la germinación. Al respecto, Anderson y Roberts (1983) y Rivera (1993) encontraron que el endosulfan con solvente aromático de tipo xileno causa virtualmente la inhibición completa de la germinación en pocas horas.
En este estudio y en otro realizado previamente por Anderson y Roberts (1983), se demuestra que diferentes formulaciones de un insecticida dado están típicamente asociadas con la inhibición de la germinación de B. bassiana, como ocurre con diazinon y endosulfan; un fenómeno similar ha sido observado en el campo por Holloway y Young (1943), Fisher y Griffiths (1950) y Hall y Dunn (1959).
El malation inhibió la germinación significativamente. En contraste, Alves (1986) señala que el crecimiento micelial en un medio sólido que contenía el tóxico fue moderadamente compatible, aunque no indica qué concentración fue utilizada; estos resultados no son necesariamente contradictorios, ya que el efecto de los insecticidas sobre la germinación conidial puede ser diferente a aquel sobre el crecimiento micelial. El fenitrotion tuvo un efecto moderado sobre la germinación, y resultados similares con crecimiento micelial fueron hallados por Rivera (1993). El pirimifosmetil y el clorpirifos no tuvieron un mayor efecto sobre el hongo, y lo mismo fue hallado para clorpirifos por Riba et al. (1983) y Foschi y Grassi (1985). El isazofos tuvo muy poco efecto sobre la viabilidad de las conidias.
La fuerte interacción en la mezcla puede ser evitada por aplicaciones separadas de los dos agentes. Sin embargo, estas no serían prácticas ni deseables para el caficultor, puesto que se aumentarían los costos de aplicación.
La evaluación de la germinación a las 24 y 48 horas permite seleccionar los insecticidas menos inhibitorios y determinar su efecto fungistático en mezcla, en contraste con los estudios realizados sobre este mismo tópico, cuya metodología se basa en la evaluación de la viabilidad de la conidia con el método de unidades formadoras de colonia (UFC), imposibilitando el estudio del efecto fungistático. Además, esta metodología podría ser utilizada para diferenciar aislamientos entre sí.
Al observar el comportamiento de la germinación del hongo en los dos tiempos evaluados (24 y 48 horas), se observó que la mayoría de insecticidas tienen un efecto fungistático más que fungicida, pues hay germinación y crecimiento micelial a las 48 horas, excepto para el diazinon PM, el feniotrion y el malation que tuvieron un efecto deletéreo sobre el hongo (Tablas 2 y 3). El efecto fungistático es explicable debido a la difusión de la mezcla en el agar, disminuyendo así la acción inhibitoria del insecticida. Harrison y Gardner (1992) obtuvieron resultados de fungistasis semejantes con herbicidas; además, Groden y Lockwood (1991) hallaron que mediante el análisis Probit de Dosis-Mortalidad contra niveles de fungistasis, revelan una significativa tendencia de aumento en los valores de la dosis letal media (DL50) con el incremento de la fungistasis. Esta fungistasis puede impactar directamente la dosis de inóculo efectivo requerido para el adecuado control de la broca. Un estudio exhaustivo en el campo es necesario para definir el nivel de dosis, la formulación del hongo y la tecnología de aplicación para evitar el efecto fungistático.