
Abstract
In this work was determinated the preference of Sitophilus zeamais for ovipositing in 30 corn genotypes, and it also was studied the relationship between oviposition per seed with the total protein content, triptophan and lysine. The most prefered genotypes for oviposition were: Ejura (1) 7843Q, PII Dia 1'90, Cacahuacintle (susceptible control), and the P3 Dia 1'90. Also they had the highest rate of damaged grains. The less prefered and with lower rate of oviposited grains were the genotypes B25C22- MHS-1-2-b, Across 8765, P7 Dia 1'90 and the Palo mero (resistant control). The protein content, lysine, and triptophan in the sample and in protein, had not influence in the oviposition preference of the insect, the adults atraction or the oviposition rate per kernel.
Introducción
El gorgojo menor de los granos, Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae), es una de las especies más destructivas y ampliamente distribuidas en todo el mundo, cuyos estados de larva y adulto causan grandes pérdidas en maíz almacenado (Widstrom et al. 1972). En trabajos desarrollados para buscar resistencia genética a esta plaga se ha tomado como criterio de selección la preferencia que este insecto muestra para ovipositar en uno u otro genotipo.
Teniendo en cuenta lo anterior, se llevó a cabo este estudio, con el objetivo de evaluar la preferencia del S. zeamais para ovipositar en materiales que previamente habían sido evaluados por resistencia al ataque de esta plaga en pruebas de confinamiento. También se determinó la relación entre el contenido proteínico de los granos y la oviposición recibida por los mismos.
Revisión de literatura
La hembra de S. zeamais deposita de 300 a 500 huevos durante toda su vida y alcanza su máxima oviposición a las 3 ó 4 semanas después de la emergencia; la oviposición en los granos la realiza luego de hacer una cavidad, la cual le permite colocar el huevo en la parte media; posteriormente, la hembra coloca un tapón de material gelatinoso sobre el sitio donde ha colocado el huevo. Esta sustancia gelatinosa queda nivelada con la superficie del grano y se seca en un corto tiempo al entrar en contacto con el aire (Ramírez 1982).
Painter (1951) definió la «no preferencia» como una condición inherente al insecto, cuya conducta le impide alimentarse u ovipositar en una planta. Al respecto, Kogan y Ortman (1978) sugirieron cambiar este término por el de «antixenosis», ya que el vegetal es el que induce en el insecto una conducta que le impide utilizarlo como huésped mediante un «mensaje» que puede ser de tipo químico o físico, o mediante características morfológicas.
Dobie (1974), en evaluaciones para determinar resistencia a S. zeamais, no encontró relación entre el número de oviposiciones en los genotipos y la susceptibilidad mostrada por estos, por lo que considera que la resistencia fue definida por factores que operan después de la oviposición.
Classen et al. (1990) encontraron que el contenido de ácido ferúlico en granos de nueve poblaciones de maíz, del banco de germoplasma del CIMMYT, correlacionó negativamente con las variables: Número de huevos, progenie, preferencia y consumo por S. zeamais; por tal razón, sugieren considerar esta sustancia como una nueva fuente de resistencia; por su parte Gómez et al. (1983) no descartaron la presencia de un estimulante de la oviposición en los granos.
Sharifi y Mills (1971) encontraron que las hembras de S. zeamais pueden ovipositar sobre varias partes del grano y que las posturas son depositadas en pequeñas cavidades abiertas en el endospermo; por otro lado, Rosseto (1972) observó que las hembras de S. zeamais presentan un comportamiento diferente para ovipositar en maíz en mazorca o desgranado.
Materiales y Métodos
Este estudio se realizó bajo condiciones controladas (T=26 ± 1°C; H.R.= 70 ± 5%) en una cámara de cría. La metodología empleada fue, con algunas modificaciones, la utilizada por Aguilera (1989) y VanderSchaaf et al. (1969). Las modificaciones se refieren a la cantidad de insectos y de maíz utilizado, a algunas variables y al tiempo de exposición de las muestras a los insectos.
En las Tablas 1, 2 y 3 se presenta la genealogía, el origen y la textura de los genotipos utilizados en el estudio, los cuales son materiales mejorados para áreas tropicales y subtropicales. Para eliminar la posible infestación de campo, estos maíces se introdujeron durante 10 días en un congelador a -10°C; posteriormente, la humedad se homogenizó entre 12 y 13% con la ayuda de un determinador de humedad marca Dickey-John, y las muestras homogenizadas se mantuvieron en la cámara de cría durante tres semanas antes de iniciar el estudio (VanderSchaaf et al. 1969). Simultáneamente se obtuvo una cría de S. zeamais en maíz Cacahuacintle, utilizando 60 insectos progenitores, de edad y sexo desconocidos, por cada 200 g de maíz. Diez días después, cuando se obtuvo oviposición suficiente, se retiraron los insectos progenitores.
Pruebas de libre elección
Para la realización de las pruebas de libre elección se tomaron 15 g de cada genotipo y se colocaron en cajas de petri plásticas de 9 cm de diámetro por 2 cm de altura. Luego, las cajas de petri se colocaron en unos recipientes o tinas metálicas de 80 cm de diámetro y 15 cm de altura, cuyas tapas tenían una malla circular en el centro para permitir el flujo de aire a su interior. Las muestras se distribuyeron alrededor de las tinas en un diseño experimental de bloques al azar con cuatro repeticiones. En el centro de la estructura se depositaron, sin separar los sexos, 450 insectos de 1 a 3 semanas de edad; la relación de insectos a gramos de maíz, fue de 1:1. Las tinas se sellaron y se llevaron a la cámara de cría. Estas pruebas tuvieron una duración de 10 días, lo cual permitió la oviposición en los diferentes genotipos; al transcurrir ese lapso se registró el número de insectos por grano, el número de adultos por grano y el porcentaje de granos ovipositados.
Determinación de las oviposiciones
Para determinar las oviposiciones en los granos se siguió el procedimiento propuesto por Frankenfeld (1948), citado por Horber (1987), que consiste en teñir las punturas que hacen las hembras al ovipositar. Los componentes del tinte son: Fucsina ácida (0,5 g), ácido acético (50,0 ml) y agua destilada (950 ml). En un vaso de precipitados, con capacidad de 600 ml, se vaciaron 200 ml del tinte y luego se agregaron 15 gramos de uno de los materiales a probar y después de 5 minutos de tinción se retiró la muestra. Así se procedió con cada una de las muestras. Con el fin de eliminar el exceso de colorante en los granos, estos se pasaron a otro vaso de precipitados de la misma capacidad que sólo contenía agua; una vez realizado el lavado, las muestras se secaron sobre papel toalla. Posteriormente, y con la ayuda de una lupa de mano y de un bisturí, se determinó el número de oviposiciones en cada muestra.
En cada genotipo se determinó: el porcentaje de proteína mediante el método autoanalizador de Technicon 2, el porcentaje de triptofano empleando el método de Opienska Blauth modificado por Hernández, y para determinar la lisina se usó un analizador de aminoácidos marca Bekman 120c. Estas metodologías fueron descritas por Villegas et al. (1985).
Variedades y sintéticos de maíz para áreas tropicales, utilizados para evaluar preferencia a S. zeamais. Chapingo, México.
= Número dado a cada genotipo en los experimentos.
TL = Tlaltizapan
PR = Poza Rica
Variedades y sintéticos de maíz con alto contenido proteínico, para áreas subtropicales, utilizados para evaluar preferencia a S. zeamais. Chapingo, México.
= Número dado a cada genotipo en los experimentos.
= Testigo susceptible
= Testigo resistente
TL = Tlaltizapan
Líneas con diferentes grados de endocría de poblaciones de maíz con alto contenido proteínico, para áreas tropicales, utilizadas para evaluar preferencia a S. zeamais. Chapingo, México.
= Número dado a cada genotipo en los experimentos.
PR = Poza Rica
Resultados y Discusión
Los resultados correspondientes al análisis de proteína demuestran que el 33,3% de los materiales tuvo un porcentaje de proteína entre 10,0 y 11,3; otro 30% entre 9,0 y 8, y el 36,6% restante osciló entre 7,5 y 8,9 (Tabla 4).
Porcentaje de proteína, triptofano y lisina (en proteína y en muestra) en granos completos de 30 genotipos de maíz utilizados para evaluar preferencia a S. zeamais. Chapingo, México.
= Testigo susceptible
= Testigo resistente
Cuadrado medio del error, valor y significancia del F calculado de las variables utilizadas para determinar preferencia a la oviposición del S. zeamais en 30 genotipos de maíz.
P > F = 1%
En la Tabla 5 se presentan los valores de F calculado, los cuales resultaron significativos al 1% de probabilidad. Esto indica que hubo diferencia de respuesta entre los genotipos a las variables estudiadas.
Los genotipos con mayor número de oviposiciones por grano fueron el 14 (Ejura (1) 7843Q) y el 30 (P11 Día 1'90) (Fig. 1), los cuales recibieron 18,66 y 9,74% más oviposiciones que el Testigo susceptible 37 (Cacahuacintle), y 233,6 y 257,9% más que el Testigo resistente 38 (Palomero). Coincidencialmente, estos dos genotipos fueron los que presentaron el nivel de susceptibilidad más alto en relación con la pérdida de peso en las pruebas de confinamiento realizadas previamente. Los genotipos 6 (Across 8765) y 36 (B25C22MH5-1-2-b) presentaron menor número de oviposiciones por grano, en cantidades de 69,3 y 71% menos que el Testigo susceptible, y 7,5 y 12,6% menos que el Testigo resistente.
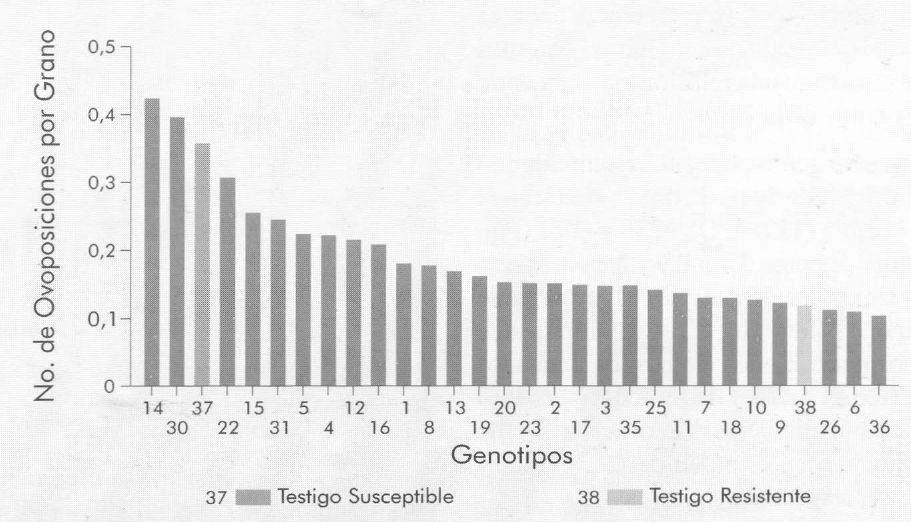
Número promedio de huevos de S. zeamais por grano en genotipos de maíz libremente elegidos, para determinar preferencia para oviposición.
La variable adultos por grano se midió para determinar la atracción del S. zeamais por los diferentes maíces. Los resultados indican que los genotipos 30 (P11 Día 1'90) y 22 (P3 Día 1'90) atrajeron mayor número de insectos por grano y estos fueron dos de los cuatro genotipos que tuvieron mayor pérdida de peso en las pruebas previas de confinamiento. Entre los menos atractivos se destacan los genotipos 9, 19, 17, 36, 35 y 26, con un total de 0,44; 0,045; 0,056; 0,057; 0,69 y 0,088 insectos por grano, respectivamente; con excepción del genotipo 36, estos genotipos en las pruebas de confinamiento presentaron la más baja pérdida de peso.
Al comparar los genotipos 30, 22, 4, 18 y 31 con el Testigo susceptible (37) (Fig. 2) se observa que no obstante el mayor número de insectos atraídos por los primeros, en cuanto al número de oviposiciones éstos fueron superados por el segundo; esta situación, dificulta concluir que la presencia de adultos en cada muestra coincide con su preferencia hacia la oviposición, aunque en ciertos genotipos se mantenga una relación estrecha entre estas dos variables.
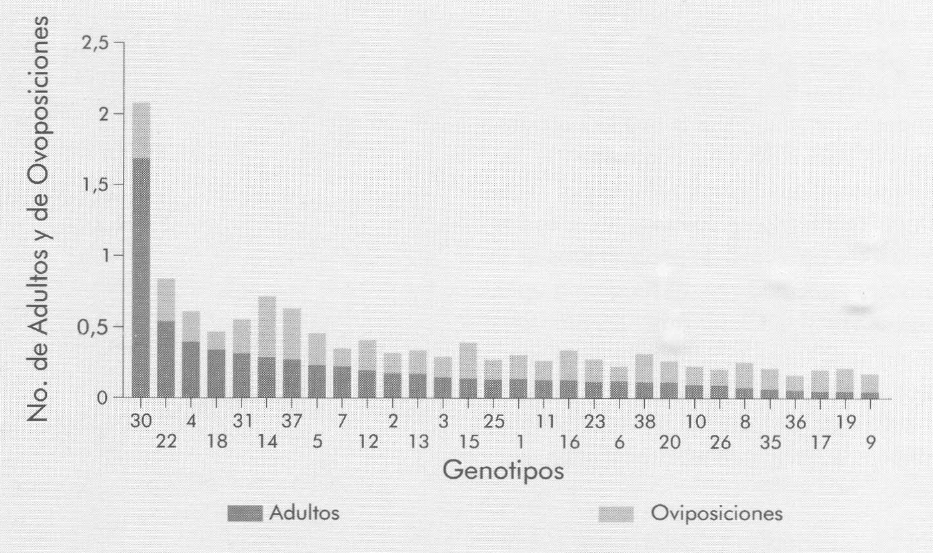
Número promedio de adultos y de oviposiciones de S. zeamais por grano en genotipos de maíz libremente elegidos, para determinar preferencia por la oviposición.
Entre los genotipos con mayor porcentaje de granos ovipositados se destacan el 14 (Ejura (1) 7843Q), el 30 y el 22 (Fig. 3) que tuvieron 3,48; 2,9 y 2,3 veces más granos ovipositados que el Testigo resistente (Palomero). Los genotipos que mostraron menos granos ovipositados fueron el 6, 26 y 18, y todos sufrieron daños ligeramente inferiores a los de Palomero en el orden del 21,7; 15,3 y 7,5%, respectivamente.
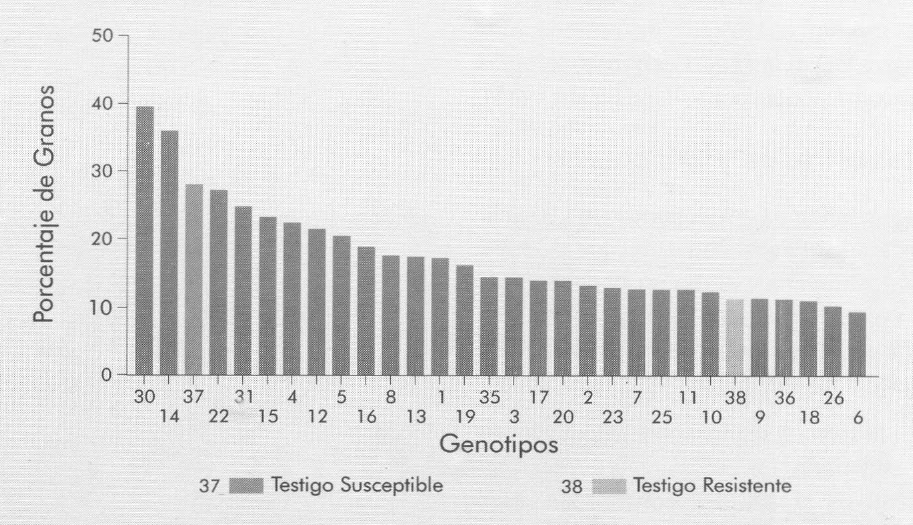
Porcentaje de granos con oviposturas (x) de S. zeamais en genotipos de maíz libremente elegidos, para determinar preferencia por la oviposición.
De un análisis conjunto entre esta variable y la de oviposiciones por grano se desprende que en la mayoría de los genotipos, los que tuvieron el mayor número de oviposiciones también tuvieron el mayor porcentaje de granos ovipositados y viceversa; esto muestra una tendencia del insecto a colocar un huevo por grano, al menos durante los 10 días en que estuvieron expuestas las muestras. También se puede decir de estas variables que su importancia está dada en la medida en que sean utilizadas para observar preferencia del insecto a la oviposición, pero no es recomendable su uso para concluir que existe resistencia, porque se ingnora que pueda pasar con los huevos y las larvas, y también el efecto que los diferentes genotipos tengan en la mortalidad, fecundidad y fertilidad de los insectos que lleguen al estado adulto. Dobie (1974) coincide con esta apreciación, al no encontrar evidencias que le permitieran relacionar el número de oviposiciones dejados por los insectos en los granos con la susceptibilidad, y por el desconocimiento de los factores que operan después de la oviposición.
En la Tabla 6 se muestra que no hubo correlación entre las oviposiciones por grano, los adultos por grano, el porcentaje de granos ovipositados y las variables que aluden al contenido proteínico; por lo tanto se afirma que el porcentaje de proteínas y su calidad (lisina y triptofano) en las muestras no tienen influencia en la preferencia de insecto para ovipositar en uno u otro genotipo.
Correlaciones entre las variables: oviposiciones por grano, adultos por grano, porcentaje de granos ovipositados y contenido proteínico en 30 genotipos de maíz evaluados por la preferencia para ovipositar del S. zeamais.
Conclusiones
Los genotipos más preferidos por el S. zeamais en pruebas de libre elección fueron: Ejura (1) 7843Q, P11 Día 1'90, Cacahuacintle y P3 Día 1'90. Igualmente, estos genotipos tuvieron el mayor porcentaje de granos dañados.
Los genotipos P11 Día 1'90 y P3 Día 1'90 atrajeron el mayor número de insectos adultos y los genotipos S86p15Q3 y S89S545Q atrajeron el menor número de insectos adultos.
Los genotipos B25C22MH5-1-2-b, Across 8765, P7 Día 1'90 y Palomero fueron los menos preferidos para la oviposición y también los que tuvieron el menor porcentaje de granos ovipositados.
Durante el tiempo de exposición de las muestras, el Sitophilus zeamais tuvo tendencia a colocar un huevo por grano.
El contenido proteínico, la lisina y el triptofano en la muestra y en proteína no presentaron influencia sobre la preferencia de los insectos para la oviposición, ni tampoco en la atracción de adultos y el porcentaje de granos ovipositados.
Es necesario determinar la relación existente entre la dureza y otras sustancias presentes en los granos y la preferencia del S. zeamais para ovipositar.