
Abstract
Currently, there is an incresing utilization of tropical savanna areas with established pastures of introduced grasses of the genus Brachiaria. Despite their excellent adaptation to acid and infertile soils, Brachiaria grasses are highly susceptible to the spittlebugs Aenelamia spp.. Here we report on the results of a study about ants associated with introduced and native pastures and their role as predators of spittlebugs. We found a group of generalist predatory ants that consume spittlebug eggs artificially placed in the field. Ants of the genera Solenopsis, Wasmannia and Pheidole were observed most persistently preying upon spittlebug egg masses and first-instar nymphs. Other species such as Paratrechina sp., Ectatomma ruidum y Camponotus sp.1 were also observed consuming spittlebug eggs. To study the diversity, species composition and abundance of ants associated with introduced and native pastures, we sampled plots by placing baits at different distances. Baits placed every 5 m captured up to 83% of the species collected with more time-consuming methods such as manual collection and nest digging. A total of 41 ant species associated with all pastures were collected with all methods. Among the introduced pastures, B. decumbens cv. Basilisk had the greatest number of associated ants (18 species). The patterns of spatial distribution of nests of Brachymyrmex sp., E. ruidum and Camponotus sp.1 in a plot before and after soil preparation for sowing Brachiaria hybrids were also documented. After soil preparation, Camponotus sp.1 nests disappeared, but nests of one Solenopsis species were found in the plot.
Introducción
En los trópicos americanos, las sabanas ocupan grandes extensiones, como es el caso de los Llanos Orientales entre Colombia y Venezuela, que cubren una superficie de aproximadamente medio millón de kilómetros cuadrados (Sarmiento 1984; Sarmiento y Monasterio 1975). Actualmente existen programas de manejo de sabanas con miras a optirnizar su utilización agrícola. Las pasturas nativas tienen baja productividad y calidad de forraje, lo que ha llevado al establecimiento de pasturas introducidas que se adaptan a las condiciones de baja fertilidad y a los suelos ácidos (CIAT 1989, 1990). Pastos del género Brachiaria han mostrado buena adaptación a las condiciones edáficas de América tropical; sin embargo, su uso es limitado por su alta susceptibilidad a varios géneros de la familia Cercopidae (Homoptera), siendo las especies de Aeneolamia las más limitantes en los Llanos Orientales de Colombia (Ferrufino y Lapointe 1989; Lapointe et al. 1989).
El programa de Forrajes Tropicales del CIAT está implementando una metodología de selección de híbridos de Brachiaria resistentes al salivazo de los pastos, Aeneolamia spp., infestando artificialmente con huevos obtenidos de una colonia masal. Esta metodología, en la que se disponen los huevos superficialmente en el campo, facilitó el aprovechamiento de los huevos por especies de hormigas depredadoras, dificultando así la evaluación de los híbridos de Brachiaria.
Algunos autores reconocen a las hormigas como uno de los grupos de artrópodos más abundantes en las sabanas tropicales (Lamotte 1975; Lavelle 1983; Lévieux 1983; Chamorro y Soto 1986). Aunque no se conoce a fondo la composición de su espectro trófico, se sabe que las hormigas tienen un papel importante en el flujo de energía en los ecosistemas de sabanas actuando como detritívoras y como consumidoras primarias y secundarias (Bucher 1982; Gandar 1982; Lamotte 1975).
Dentro de las consumidoras primarias se encuentran hormigas altamente especializadas y de gran importancia económica como las pertenecientes a la tribu Attini, especialmente Atta y Acromyrmex, siendo A. landolti (Forel) una plaga importante, por consumir gramíneas de alto valor forrajero como Andropogon gayanus Kunth (Eiten 1982; Fowler 1985; Lapointe et al. 1990). Del grupo de consumidoras secundarias es poco lo que se conoce. Existen algunos trabajos realizados principalmente en la sabanas africanas por Lamotte (1975) y Lévieux (1983). En Suramérica, a excepción de las Attini, no se conoce aún la fauna de hormigas que ocupa los ecosistemas de sabanas, aunque existen trabajos muy generales como el de Zuccaro y Bulla (1985) y el de Chamorro y Soto (1986).
En el grupo de hormigas consumidoras secundarias hay especies depredadoras oportunistas, que se han denominado así por su capacidad de explotar recursos alimenticios temporales y disponibles (Brian 1983; Holldobler y Wilson 1990). Estas especies se han caracterizado, además, por su gran capacidad de colonización, encontrándose asociadas a sistemas de cultivos y áreas perturbadas (Risch y Carroll 1986).
En general, las actividades agrícolas tienen un efecto sobre la dinámica natural de los ecosistemas; la preparación del suelo, así como otras prácticas agronómicas, afectan la fauna de artrópodos, eliminando especies y dando paso a la colonización por otras (Saks y Carroll 1980; Risch y Carroll 1982). La fauna de invertebrados en sabanas tropicales es todavía desconocida, así como también es desconocido el efecto en general del establecimiento de pasturas introducidas sobre tal ecosistema. Con la presente investigación se quiere contribuir al conocimiento de la fauna de hormigas asociadas con sabanas nativas y pasturas introducidas, estableciendo diferencias en la composición de especies en áreas nativas y perturbadas. También se examina el posible efecto de un grupo de especies depredadoras sobre los huevos y ninfas del salivazo de los pastos.
Materiales Y Métodos
El trabajo se realizó en el Centro de Investigación "Carimagua" del ICA-CIAT, ubicado en San Pedro de Arimena (Meta), con una localización geográfica de 4° 30' de latitud norte y 7° 30' de longitud oeste y con una altura sobre el nivel del mar de 200 m. Corresponde a la zona de los Llanos Orientales caracterizada como sabana isohipertérmica bien drenada con una pluviosidad promedio anual de 2.240 mm. Los suelos son infértiles y ácidos (Fletcher y Cortez 1983).
En Carimagua se encuentran áreas dedicadas a la investigación en forrajes tropicales, calidad y productividad de pasturas y manejo de sabanas nativas entre otros proyectos. Parte del área se encuentra sembrada con ensayos de plantas forrajeras introducidas como pastos del género Brachiaria y A. gayanus, así como también se encuentran áreas de sabana nativa, las cuales están siendo caracterizadas en su composición florística. El trabajo de inventario de especies de hormigas se realizó principalmente en estos tres tipos de vegetación: sabana nativa, A. gayanus y Brachiaria spp.
Colección de Hormigas
Se realizaron colecciones de hormigas en los siguientes tipos de forrajes: Brachiaria decumbens Stapf. cv. Basilisk, B. dictyoneura cv. Llanero, B. humidicola cv. Pasto Humidícola, B. brizantha (Hochst) Stapf. cv. Marandú, A.gayanus cv. Carimagua 1 y en la sabana nativa. El material se obtuvo por medio de colección manual, excavación de nidos y a través de muestreos con cebos, como se describe a continuación.
- Colección manual y excavación de nidos
La colección manual de hormigas se realizó revisando la superficie del suelo, la parte aérea y radicular de las plantas. Se excavaron los nidos reconocidos en las diferentes praderas, colectando, en lo posible, miembros de las diferentes castas de la sociedad (obreras, reinas y machos).
- Muestreo con cebos
Se utilizaron trampas cebadas que consisten en cajitas circulares de 3,5 cm de diámetro (área 7,1 cmβ), las cuales tienen perforaciones de 2 x 2 mm de lado, un total de 44 en la cara inferior y 34 en la cara superior. En su interior se dispuso sardina enlatada como cebo. Las trampas se colocaron sobre la superficie del suelo, distribuyéndolas de forma uniforme en el área muestreada, a distancias que variaron entre 3 y 15 m. El tiempo de exposición de las trampas en el campo fue de cuatro horas, luego del cual se recogieron en cajas petri y se sellaron con una película parafinada. Las muestras luego se llevaron al laboratorio para su separación e identificación. El material colectado se determinó, al menos, hasta el nivel taxonómico de género, empleando las claves de Holldobler y Wilson (1990), Mackay y Mackay (1989) y Fernández (1990). Algunos especímenes fueron enviados a especialistas para su confirmación e identificación a especie.
- Evaluación de una Metodología de muestreo de hormigas en ecosistemas de sabanas.
En una sabana nativa, en una sabana derivada (sabana intervenida que presenta plantas nativas e introducidas) y en un lote de híbridos de Brachiaria, se realizó un mayor esfuerzo de colección, combinando los diferentes métodos de muestreo: cebos colocados en varias fechas, excavación de nidos y colección manual. Además se evaluó la eficiencia de captura del muestreo con cebos, seleccionando una densidad de cebos que permitiera colectar un porcentaje conocido de las especies capturadas empleando los diferentes tipos de muestreo. Con este fin, se realizó un ensayo con tres densidades de cebos en un diseño de bloques al azar. Cada una de las vegetaciones arriba mencionadas se dividió en tres bloques, cada uno con tres subparcelas con una densidad diferente de cebos. En la sabana nativa y en la sabana derivada los tratamientos evaluados fueron 4, 9 y 36 cebos por subparcela de 30 x 30 m (i.e., 5, 10 y 15 m de distancia entre cebos).
En el lote de híbridos de Brachiaria se aumentó la densidad de los cebos reduciendo el tamaño de las parcelas a 18 x 18 m y variando las distancias entre los cebos a 3, 6 y 9 m.
El ensayo se repitió en tres fechas diferentes para los tres tipos de vegetación. El análisis de los datos se realizó mediante un análisis de varianza para un diseño de bloques al azar, con estructura factorial (SAS 1982). Las fuentes de variación fueron día, evaluación y tratamientos (densidad de cebos).
Se calculó el índice de diversidad de Shannon (Ludwing y Reynolds 1988) para las especies de hormigas colectadas en los diferentes tipos de vegetación. Para la sabana nativa y para un lote de híbridos, la diversidad fue estimada en tres ocasiones y los valores obtenidos fueron comparados por medio de la prueba de Manny Whitney (Siegel 1978).
Reconocimiento de Nidos
Para el reconocimiento de nidos de las especies de hormigas de la sabana se identificaron en la superficie del suelo, las aberturas y entradas a las galerías subterráneas pertenecientes a hormigas, identificando la especie que la habita. Al reconocer las entradas, estas se marcaron y posteriormente se realizaron excavaciones para buscar cámaras con crías, así también como para colectar muestras de los miembros de la sociedad.
Distribución Espacial de los Nidos
Por el fácil reconocimiento de sus nidos y su alta frecuencia en los muestreos, se seleccionaron tres especies de hormigas para estudiar su distribución espacial y ver el efecto de la preparación del suelo sobre dicha distribución. Se aplicó el método recomendado por Ludwing y Reynolds (1988), que consiste en marcar un transecto en la mitad de una área y dentro de esta marcar unidades muestrales. Se realizó un transecto de 217 m de largo x 3 m de ancho, y se marcaron unidades muestrales de 3 x 3 m. En estas unidades se señalaron y contaron los nidos de tres especies de hormigas: Ectatomma ruidum (Roger), Brachymyrmexsp. y Camponotus sp. 1. Con los datos obtenidos se estableció el patrón de distribución, computando la varianza con el número de nidos a diferentes tamaños de bloques. Los bloques se formaron uniendo varias unidades muestrales contiguas. Se hicieron gráficos de la varianza contra el tamaño de las unidades muestrales y se obtuvieron los patrones de distribución espacial de los nidos de las especies antes y después de la preparación del suelo para la siembra de un lote de híbridos de Brachiaria.
Hormigas Depredadoras
Masas de huevos y ninfas del salivazo de los pastos, A. reducta Lallemand, fueron colocadas sobre papel filtro. Los huevos y ninfas se obtuvieron de una colonia del insecto mantenida en el CIAT, en Palmira (Valle) (Lapointe et al. 1989). Las masas se dispusieron en la superficie del suelo en lotes de B. decumbens cv. Basilisk y de híbridos de Brachiaria. Se realizaron observaciones sobre la depredación por hormigas y se colectaron especímenes para su identificación.
Resultados Y Discusión
Hormigas Asociadas con Forrajes Tropicales
Se colectó un total de 41 especies de hormigas pertenecientes a 19 géneros de las 6 subfamilias neotropicales (Tabla 1). El mayor número de especies correspondió a la subfamilia Myrmicinae, en la cual se encontraron varias especies de los géneros Solenopsis y Pheidole.
Especies de hormigas colectadas en diferentes tipos de vegetación en "Carimagua". San Pedro de Arimena (Meta).
B. de. = Brachiaria decumbens; B. Br. = B. brizantha; B. dy. = B. dyctioneura; B. h. = B. humidicola; A. ga. = Andropogum gayanus; S. na. = Sabana nativa; Hib. = Híbridos; S, d. = Sabana derivada. (X = colectadas con cebos; + = otro tipo de colección).
Con todos los métodos de colección empleados se colectaron 15 especies en la sabana derivada y un total de 12 especies de hormigas en la sabana nativa y en el lote de híbridos de Brachiaria. Al evaluar las diferentes densidades de cebos se encontraron diferencias significativas en el número de especies y de individuos colectados con 4, 9 o 36 cebos en las dos áreas evaluadas: 900 mβ (Fig. 1; ANOVA: F = 39,3; G.L. = 2,24; P = 0,0001), y 324 mβ (Fig. 2; ANOVA: F = 45,94; G.L. = 2,12; P = 0,0001). El mayor número de especies se colectó con 36 cebos en 900 mβ y 36 cebos en 324 mβ. El mayor porcentaje de especies obtenido con cebos del total de especies de hormigas colectadas, se obtuvo con la densidad de 36 x 900 mβ, o sea con cebos a 5 m de distancia. Con este método se obtuvo hasta el 83% del total de las especies. El método de muestreo con cebos fue el más efectivo y rápido, por lo cual se siguió utilizando en los muestreos posteriores.
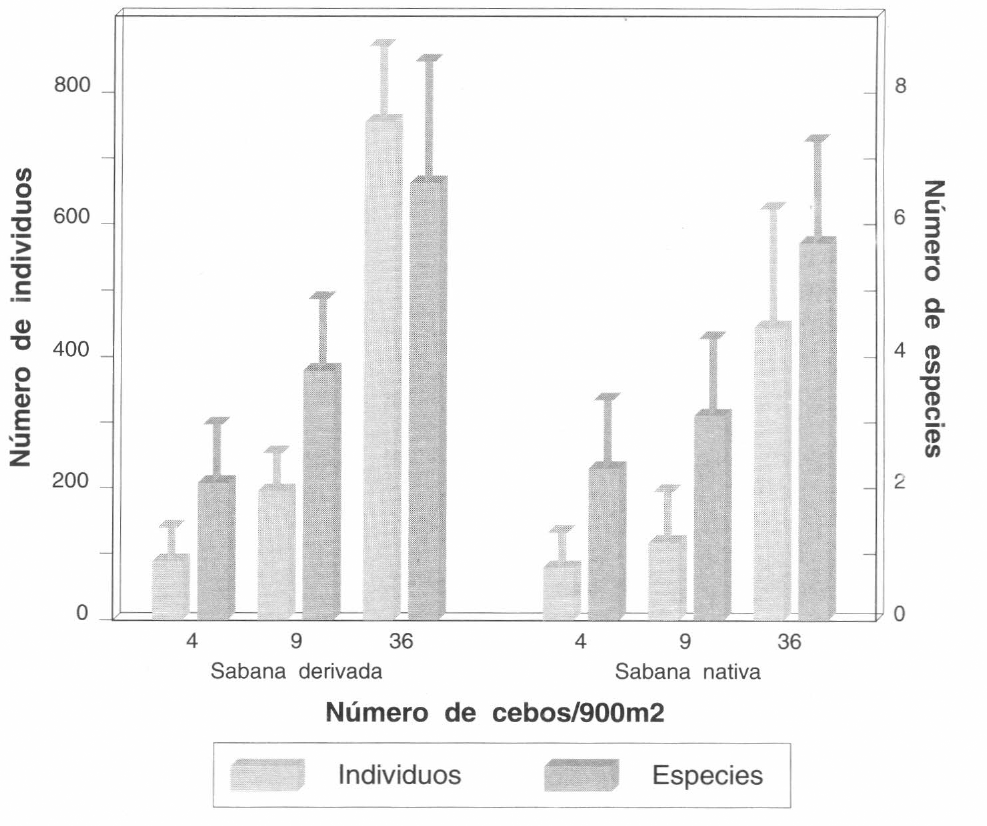
Número promedio de especies e individuos de hormigas colectados con 4, 9 у 36 cebos colocados en un área de 900 mβ, en una sabana derivada y en una nativa. Las barras de error corresponden a la desviación estándar (n=9).
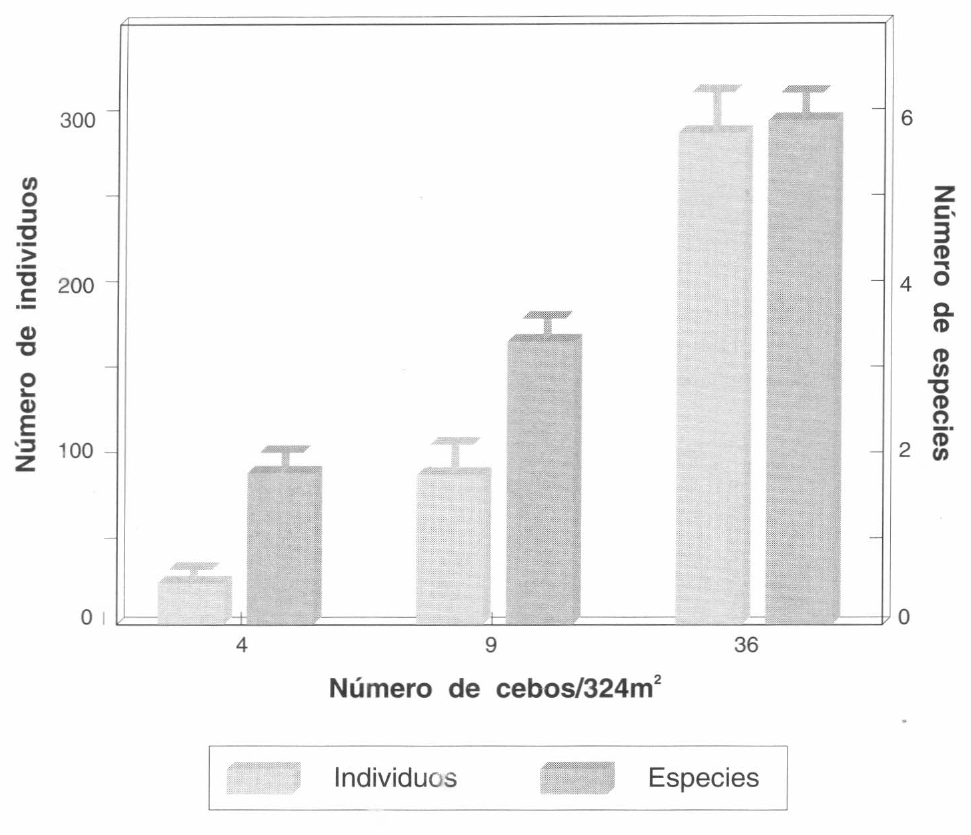
Número promedio de especies e individuos de hormigas colectados con 4, 9 y 36 cebos en un área de 324 mβ en un lote de híbridos de Brachiaria. Las barras de error corresponden al error estándar (n=9).
Se encontraron diferencias tanto en el número como en la composición de especies de hormigas en las diferentes praderas muestreadas, encontrándose el mayor número de especies en la sabana nativa, la sabana derivada y en A. gayanus. El menor número de especies se encontró en las praderas de Brachiaria (Tabla 1).
La diferencia en la composición de especies se relacionó con las diferencias en el tipo de vegetación que conforman las praderas estudiadas. Las especies depredadoras dependen de la fauna de fitófagos que visitan las plantas, y para las especies consumidoras de material vegetal son importantes los atractivos que la planta ofrece, tales como los nectarios u otro tipo de exudados, además de forraje, semillas e inflorescencias.
La sabana nativa esta conformada por una gran diversidad de plantas entre gramíneas, leguminosas y otras. En lugares con plantas nativas se encontraron especies de hormigas que no se encontraron en las praderas con un sólo tipo de forraje. Algunos pastos como A. bicornis, A. selloanus y A. gayanus presentaron nectarios en la base de las láminas foliares, por lo cual se observaron soportando altas poblaciones de por lo menos dos especies de Camponotus. En este tipo de plantas también se encontraron escamas y cóccidos (Homoptera) que atraían especies de hormigas como Camponotus y Paratrechina. Se encontraron otras especies como las del género Pheidole y la especie Pseudomyrmex termitarius (F.R. Smith), las cuales se observaron depredando termitas. Estas especies no se encontraron en los otros tipos de vegetación.
Las especies más abundantes y ampliamente distribuídas fueron E. ruidum Roger y Brachymyrmex sp., las cuales se encontraron en todos los tipos de vegetación estudiados. Solenopsis sp. 4 se encontró en todos los tipos de vegetación menos en la sabana nativa. Se encontraron dos especies nuevas del género Cyphomyrmex colectadas en lotes de híbridos de Brachiaria, las cuales se observaron llevando inflorescencias a sus nidos. Se colectaron especímenes del género Ephebomyrmex, recientemente registrado en Colombia*. Esta especie fue colectada en una zona caracterizada por suelos con un alto contenido de arcilla y se encontró alimentándose de semillas principalmente de B. dictyoneura y Axonopus purpusii.
I. Zenner de Polanía. Comunicación personal. C.I. "Tibaitatá", ICA, Apartado Aéreo 151123 Eldorado. Santafé de Bogotá, Colombia.
En la Tabla 2 se muestran los valores de los índices de diversidad calculados para los diferentes tipos de vegetación estudiados. B. decumbens cv. Basilisk fue la especie de Brachiaria con mayor índice de diversidad y número de especies de hormigas, 18 en total. Esta planta presenta macollas grandes y a su alrededor se acumula mucho material vegetal y hojarasca, lo que crea un sitio ideal para los nidos temporales de muchas especies de hormigas. B. humidicola cv. Pasto Humidícola y B. dictyoneura cv. Llanero fueron las que presentaron el menor número de especies de hormigas encontradas y el índice de diversidad más bajo. Se compararon los índices de diversidad para la sabana nativa y un lote de híbridos de Brachiaria. No se encontraron diferencias estadísticamente significativas (Prueba de Mann y Whitney: U = 9; n₁ = n₂ = 3; P = 0,1; prueba de dos colas).
Indice de Diversidad de hormigas y ocupación de cebos en siete tipos de vegetación en Carimagua. San Pedro de Arimena (Meta).
Promedio de tres muestras. No significativamente diferentes, Prueba de Mann y Whitney.
Reconocimiento de nidos
Todas las especies de hormigas se encontraron nidificando en el suelo. Algunas especies presentaron galerías definidas con entradas características, mientras que otras, especialmente varias especies de Myrmicinae, se encontraron en sustratos temporales.
Entre las especies que se encontraron en nidos definidos estan: E. ruidum, con una entrada caracterizada por una torrecita de barro con un agujero circular de aproximadamente 5 mm de diámetro; Camponotus sp. 1 también presenta una entrada circular de menor tamaño que la de E. ruidum, pero bien definida; Brachymyrmex se reconoció por los restos de subsuelo de color rojizo en forma de montoncitos pequeños; y Pheidole sp. forma sociedades grandes con galerías hasta de un metro de profundidad, cuya entrada es una abertura de forma irregular y oblicua.
Distribución Espacial de los Nidos
Las Figuras 3 y 4 muestran los patrones de distribución espacial de los nidos de E. ruidum, Brachymyrmex sp., Camponotus sp. 1 y Solenopsis sp. 2, en una pradera, antes y después de la preparación del suelo. El patrón agregado se reconoce por una relación varianza promedio σβ/x>1 y en la Figura 1 la varianza aumenta formando un pico de agregación. Este patrón lo presentó Brachymyrmex sp. antes y después de la preparación del suelo. Una distribución al azar se reconoce por una curva irregular, donde los valores de varianza σβ fluctúan al azar dentro de los diferentes tamaños de unidades muestrales. Este patrón lo presentó E. ruidum antes de la preparación. El patrón de distribución uniforme se caracteriza por una varianza baja que no fluctúa en los diferentes tamaños de unidades muestrales. Camponotus sp.1 presentó este tipo de distribución antes de la preparación del suelo. Después de la preparación del suelo y la siembra de un lote de híbridos de Brachiaria, los nidos de Camponotussp. desaparecieron completamente y un mes después la especie no apareció en los muestreos ni fueron observados sus nidos. Camponotus sp. 1 se ha observado alimentándose de los nectarios de plantas de A. gayanus y A. bicornis. Al reemplazar una pradera nativa por un lote de híbridos de Brachiaria desaparecen este tipo de gramíneas influyendo posiblemente en el desplazamiento de Camponotus. Después de la preparación del suelo aparecieron nidos de una especie de Solenopsis, los cuales presentaron un patrón de distribución al azar. Esto mostró la gran capacidad de colonización y adaptación a áreas modificadas de especies de tipo generalista como son las Solenopsis.
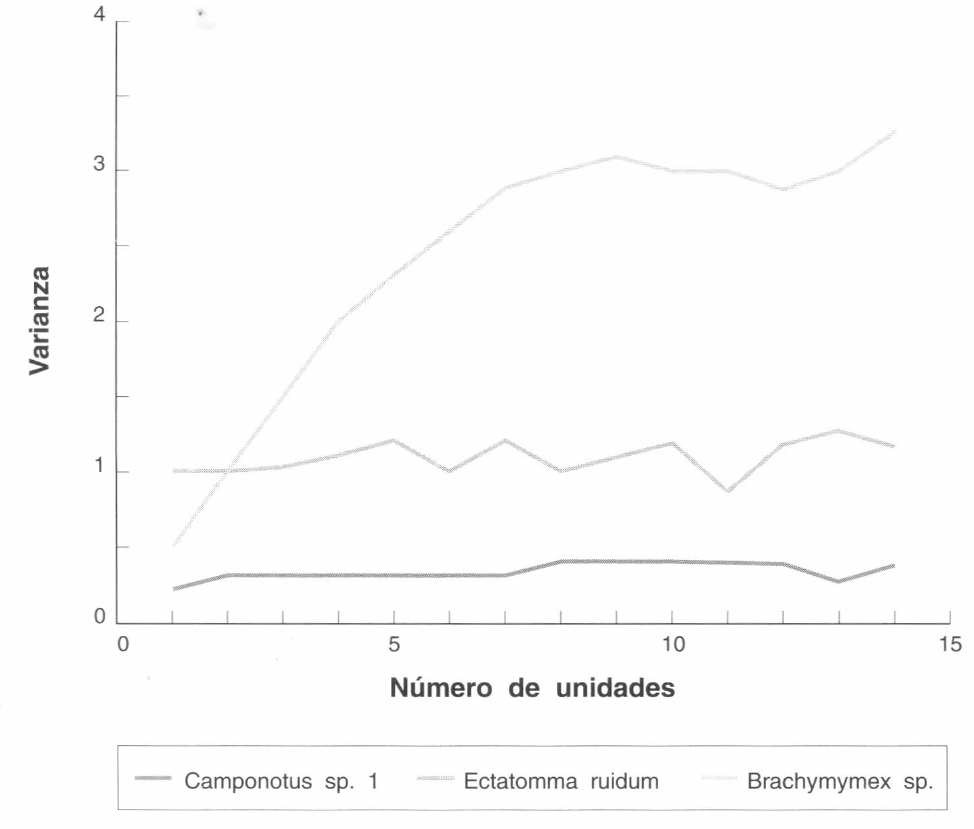
Patrones de distribución espacial de nidos de E. ruidum, Brachymyrmex sp. y Camponotus sp.1 en una pradera antes de la preparación del suelo en Carimagua.
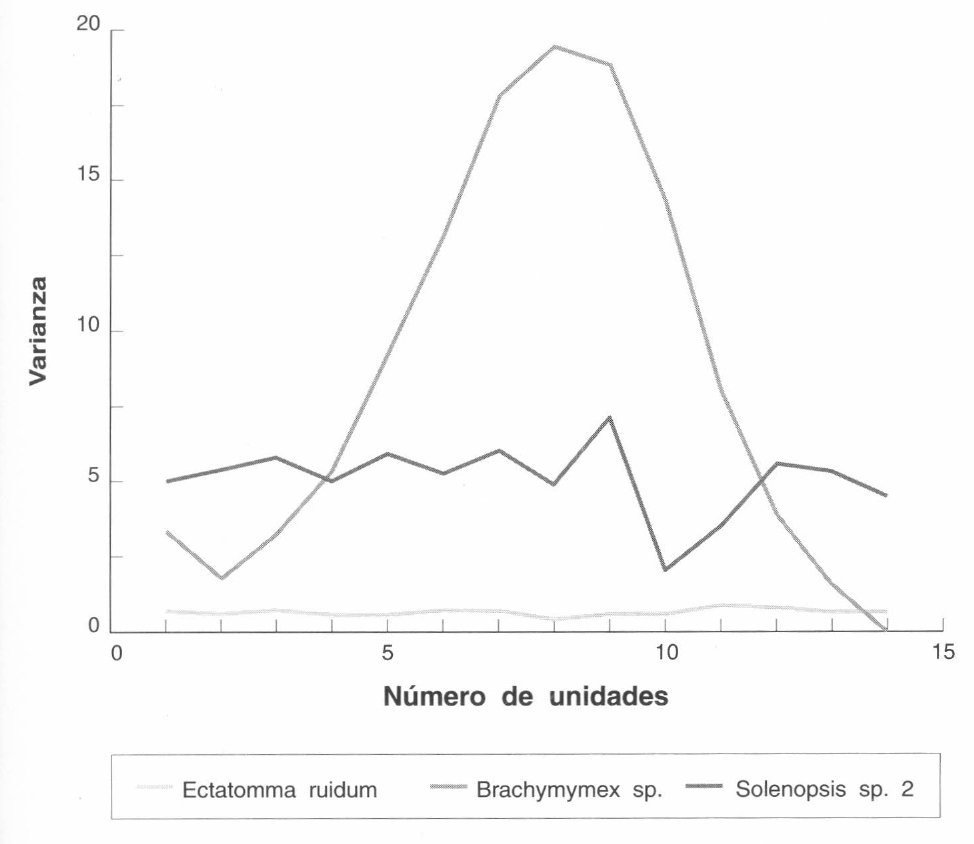
Patrones de distribución espacial de nidos de E. ruidum, Brachymyrmex sp. y Solenopsis sp.2 después de la preparación del suelo y la siembra de un lote de híbridos de Brachiaria.
Hormigas Depredadoras
En el estudio se encontró un grupo de especies de hormigas depredadoras generalistas principalmente de los géneros Solenopis, Wasmannia y Pheidole. Estas especies se observaron llevando huevos y ninfas del salivazo de los pastos de las masas locadas superficialmente. Se realizaron observaciones repetidas de una hora, en las cuales especies como Solenopsisy Pheidole llevaron al nido masas de más de 200 huevos. Las diferentes especies de Solenopsis (subgénero Diplorhoptrum) fueron las que más rápido localizaron la fuente de alimento, forrajeando en gran número sobre los huevos. Estas especies tienen un sistema de comunicación avanzado, lo que les permite localizar el alimento y reclutar rápidamente a los otros miembros de su colonia (Holldobler y Wilson 1990).
Paratrechina sp.1 y E. ruidum se observaron sobre la masa de huevos de una forma menos persistente. A pesar de que E. ruidum consume presas grandes, también se le observó llevando masas de huevos. Camponotus sp.1 se observó consumiendo huevos directamente sobre la masa sin llevarlos al nido.
Aunque no se midió el efecto de estas especies depredadoras sobre la plaga bajo condiciones naturales, se conoció de la capacidad de estas especies para aprovechar diferentes tipos de recursos alimenticios, sobre todo si estos se presentan de manera abundante. La infestación artificial con huevos de salivazo en los híbridos de Brachiaria se ha visto obstaculizada por la depredación de estas especies de hormigas sobre los huevos utilizados en el ensayo.
Bajo condiciones naturales, el salivazo protege sus huevos al enterrarlos en el suelo, lo cual hace poco probable que las hormigas los depreden. Las ninfas de primer instar están más expuestas a la depredación por hormigas mientras buscan un lugar para fijarse a la planta y secretar la espuma que les dará protección hasta el estado adulto. Hasta ahora no se conoce que las hormigas tengan un efecto sobre la plaga, pero en este trabajo, como en el de Hewitt y Nilakhe (1986) en Brasil, se reconoce que las hormigas son potenciales depredadoras de huevos y ninfas de varias especies del salivazo de los pastos. Bajo condiciones naturales, las hormigas pueden estar teniendo efectos significativos sobre la plaga al depredar ninfas de primer instar, sin que este efecto haya sido evaluado.
Ultimamente, en diferentes partes del mundo se están realizando investigaciones sobre la dinámica de especies de hormigas depredadoras generalistas en agrosistemas (Saks y Carroll 1980; Way y Khoo 1992; Perfecto y Sediles 1992). Estas investigaciones muestran resultados similares en cuanto a que existen especies de hormigas con gran capacidad de colonizar áreas perturbadas y que en la mayoría de los casos están actuando como controladoras de otros insectos plagas en diferentes sistemas de cultivos.
Footnotes
Agradecimientos
Los autores agradecen al Centro de Investigaciones "Carimagua" y la colaboración prestada por el equipo de Protección de Plantas, en especial la ayuda de Humberto Zamora en el trabajo de campo; a Fernando Fernández del Instituto de Ciencias Naturales, Universidad Nacional de Colombia, en Santafé de Bogotá; a Edgar Palacios y Philip S. Ward de la Universidad de California, quienes colaboraron con la identificación de las hormigas. También se agradece a Miguel S. Serrano por su apoyo en el desempeño del trabajo, así como a todo el personal del laboratorio de Entomología de Forrajes Tropicales del CIAT, en Palmira. Agradecemos a J.W. Miles por todos sus aportes y a Fernando Zapata por sus sugerencias y comentarios.