
Abstract
Direct count of tetranychid mites in field or laboratory is a very unefficient and tedious task, due to the high populations that develop on cassava. For this reason a faster and confiable method needs to be developed. On these bases a work was carried out to established the relationship between the population scale used in the field and direct countings of mites in the laboratory, and to observe the stability of such a scale on different plant strata and crop ages. Concerning the phytoseiid mites, it was pretended to verify the presence-absence score between field and laboratory counts. This study was developed during two crop cycles al CIAT-Palmira, using the cassava variety CMC-40. Counts were made every two weeks in 20 plots of 144 plants each. A central plant was chosen and every other leaf was picked after the first completely developed leaf. On individual leaves from 5 randomly selected plants phytophagous mites were counted under the stereo-microscope discriminating stadia and species. All phy- toseiid mites present were collected on each evaluated node; on each level of the plant a three minute sample was taken of all the tetranychids mites for taxonomic purposes. It was possible to determine the coefficients that allow conversion of field scores into real population estimates of cassava tetranychid mites. Confidence limits for the sampling procedure were also determined. An error percentage was established for presence-absence evaluations of phytoseiid mites. Additional information was obtained on the estability of the field scale on the various plant strata.
Introducción
El manejo de ácaros plagas, según Hoyt 1969, Croft et al. 1976 y Hoy 1984, citados por Jones (1990), a menudo requiere de un cuidadoso monitoreo de la plaga y de sus enemigos naturales. Desafortunadamente, el tamaño pequeño de los ácaros y la densa telaraña asociada con algunas especies hace que su conteo sea una de las tareas más arduas y demoradas en el manejo de los ácaros plagas. Para solucionar este problema, los investigadores han desarrollado y usado, en varios cultivos y para determinadas especies, planes de muestreo secuencial o binomial, o ambos, los cuáles son más rápidos y seguros que aquellos en los cuales todos los ácaros son contados sobre un número predeterminado de hojas por planta (Jones 1990).
El uso de planes de muestreo para el monitoreo de la población y la estimación del daño está bien desarrollado para ácaros de la familia Tetranychidae es sistemas agrícolas en las zonas templadas. En el cultivo de la yuca se han realizado varios procedimientos de muestreo para Mononychellus sp., utilizando métodos cuantitativos, tanto en Africa como en Colombia.
Braun et al. (1985) desarrollaron el sistema presencia-ausencia para Mononychellus spp. y ellos lo consideran como un método preciso y sensible a cambios en el nivel de la población de los ácaros y que además no requiere de equipos sofisticados. En este método, la unidad de muestra (la hoja) es examinada para verificar la presencia de uno o más individuos de una especie en particular, sin tener en cuenta el número presente.
Entre tanto en Africa, Yaninek et al. (1989) desarrollaron un protocolo de monitoreo de M. tanajoa (Bondar), con el cual se midió la incidencia y la magnitud de la infestación del ácaro verde y los síntomas de daño en la planta de yuca. Según los autores, este procedimiento permite examinar cuantitativamente un gran número de plantas en corto tiempo. El nivel de abundancia de los ácaros es determinado por evaluación de la primera hoja, totalmente expandida o desarrollada en 30 plantas seleccionadas al azar. La primera hoja totalmente desarrollada se distingue de las hojas del cogollo por el color oscuro y de las hojas más viejas porque el pecíolo está unido al tallo con un ángulo de 90 grados.
En el presente trabajo se intentó adoptar el sistema utilizado en Africa, es decir, hacer uso de "Quick Counts" a través de una escala poblacional bajo las condiciones del CIAT y con un complejo de especies de Mononychellus un poco diferente. Para desarrollar el trabajo se propusieron los siguientes objetivos:
Establecer la relación entre una escala poblacional de uso en campos de yuca y el conteo directo de ácaros del complejo Mononychellus en el laboratorio.
Probar la validez de la escala para la cuantificación de las poblaciones en los distintos estratos de la planta a diferentes edades del cultivo.
Determinar el grado de acierto en la aplicación del método "Presencia-Ausencia" en la valoración de ácaros fitoseiidos bajo condiciones de campo y laboratorio.
Realizar un seguimiento cuantitativo de las especies de Tetranychidae y Phytoseiidae en dos ciclos de cultivo.
Materiales Y Métodos
El experimento se realizó en el CIAT-Palmira (Valle), en la variedad CMC-40 clasificada como susceptible a las especies de Tetranychidae que atacan la yuca.
El primer ciclo del ensayo se realizó entre enero y octubre de 1990, en el lote Punta de Lanza y el segundo entre octubre de 1990 a mayo de 1991 en el lote del Gallinero. El área de cada ensayo fue aproximadamente de media hectárea, distribuída en 20 parcelas de 12x12 m, con 144 plantas cada una y una distancia de siembra de un metro. De cada parcela se escogieron 36 plantas centrales dejando tres surcos de barrera.
Muestreo de campo
Los muestreos se hicieron cada 15 días a partir de los tres meses de edad del cultivo. En cada parcela se tomó una planta y de las 20 plantas se escogieron 5 al azar, de las cuales se colectaron todas las hojas, se colocaron en bolsas plásticas y se llevaron al laboratorio.
Las plantas usadas en cada fecha para el muestreo se marcaron, para así no ser tenidas en cuenta en las evaluaciones posteriores.
Las hojas se colectaron a partir de la primera hoja superior más desarrollada, tal como lo define Yaninek et al. (1989 a,b), y de ahí se colectó una hoja de por medio hasta la parte basal de la planta.
En el campo, para cada hoja evaluada se utilizó la siguiente escala poblacional de M. tanajoa, sugerida por Yaninek et al. (1989):
Esta escala se utilizó para el complejo de ácaros del género Mononychellus presente en CIAT y para Tetranychus urticae Koch. Para la evaluación de los fitoseiidos se usó el sistema de "Presencia-Ausencia", y además los ácaros se colectaron y contaron en su totalidad. Para Oligonychus peruvianus (McGregor) se tuvo en cuenta el área del lóbulo central y se contaron sólo las telarañas infestadas.
Conteo en el laboratorio
En el laboratorio y con la ayuda de un estereoscopio, sobre las hojas de las 5 plantas escogidas en el campo se hizo conteo de los huevos de T. urticae y de Mononychellus spp., así como de los estados móviles de Mononychellus representados por las larvas, ninfas y adultos.
Toma de muestra de Tetranychidae
Una vez realizado el conteo total, las hojas se separaron por niveles de la siguiente manera: de la hoja 1-10; 11-20; 21-30; 31-40; 41-50 у 51-60, y de cada grupo se colectaron los especímenes de Tetranychidae para fines taxonómicos. Esta colección se hizo en un período de tres minutos.
Resultados
Primer ciclo de cultivo-Lote Punta de Lanza
Vale la pena destacar, en primer lugar, el complejo de especies de las familias Tetranychidae y Phytoseiidae que se presentó en este primer ciclo y su distribución a lo largo de las plantas. En la Figura 1 se muestra el porcentaje en el cual se encontró cada una de las especies en los diferentes niveles de la planta.
En este primer ciclo se pudo constatar que la especie fitófaga predominante fue M. mcgregori (Flechtman & Baker). La presencia de esta especie fue algo muy llamativo, debido a que su aparición, desde hace por lo menos 10 a 15 años, siempre fue algo esporádica y en bajas densidades. Sin embargo, durante este ciclo se expresó en altas poblaciones, desplazando casi por completo a M. tanajoa que fue la especie predominante hasta entonces. M. mcgregori se encontró distribuída a lo largo de toda la planta, es decir en hojas de todo tipo, pero su mayor concentración se registró en los niveles alto y medio.
Asociados con M. mcgregori se encontraron, en bajos porcentajes, M. tanajoa, T. urticae y O. peruvianus.
Con relación al complejo de los ácaros depredadores, la especie más frecuentemente encontrada en este ciclo fue Typhlodromalus limonicus (Garman & McGregor), distribuída también a lo largo de la planta, aunque sus mayores poblaciones se registraron en los niveles superiores. En menor proporción se encontraron Euseius ho, E. naindaimei (Chant & Baker), E. concordis y Amblyseius aerialis (Muma).
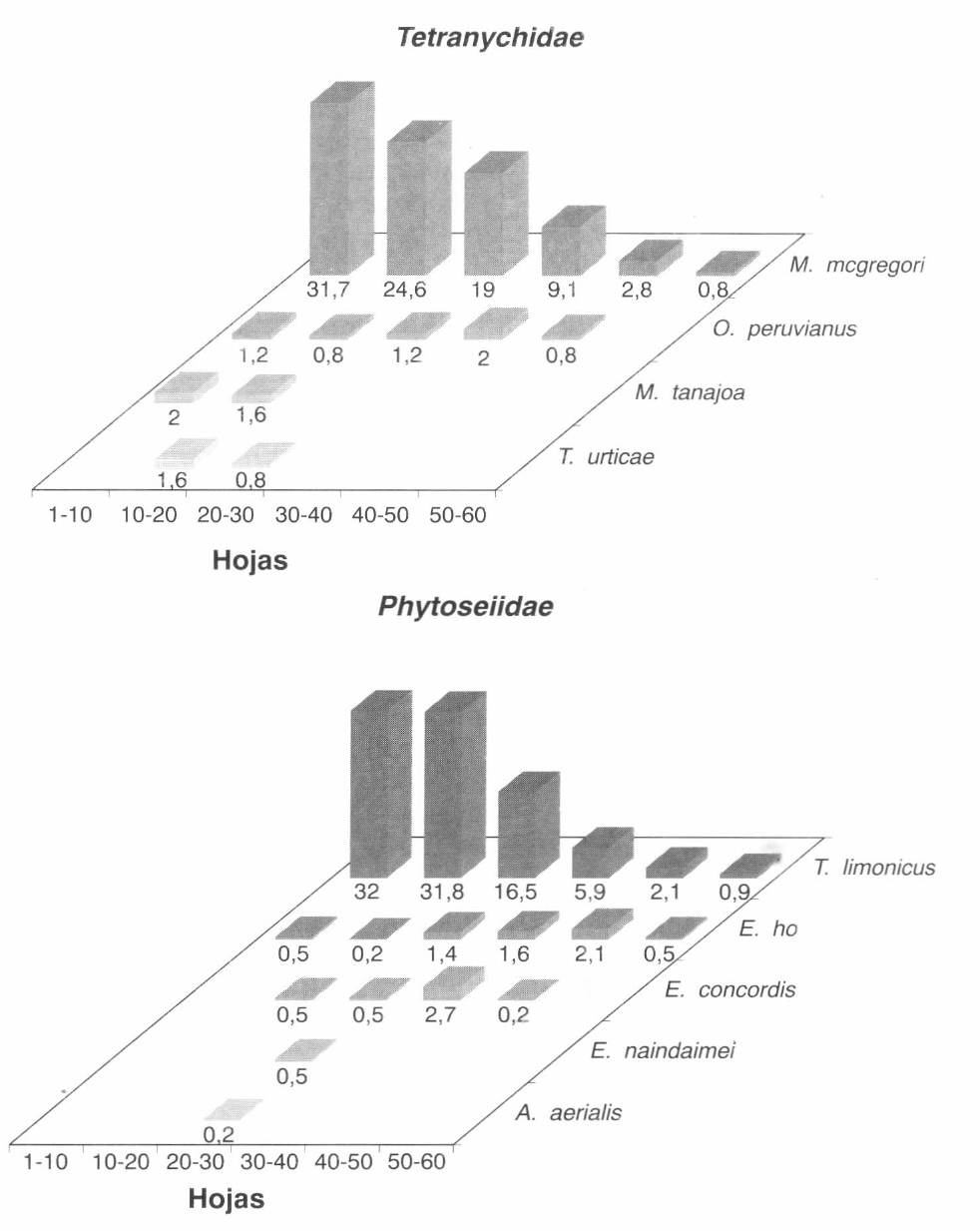
Distribución vertical de las especies de Tetranychidae y Phytoseiidae durante el desarrollo del primer ciclo del cultivo (Lote Punta de Lanza).
Los resultados relacionados con la distribución vertical de Mononychellus son similares a los registrados por Braun et al. (1985), quienes para M. tanajoa expresan que en términos del promedio de Mononychellus spp./hoja, se encuentran ácaros en toda la planta sin tener en cuenta la fase de la infestación, y en cuanto a la importancia del nivel de la hoja en la distribución de la población del ácaro demuestran que la infestación se concentra en los nudos intermedios de la planta durante la fase ascendente. También indican que no hay una diferencia significativa entre las distribuciones de la fase ascendente, en cambio, en la fase descendente, la población aparentemente se redistribuye hacia la parte superior de la planta.
Por otra parte, Yaninek et al. (1989a), en condiciones de Africa, mencionan que la distribución de M. tanajoa en las plantas, durante la estación seca, es más abundante en las hojas jóvenes sin tener en cuenta la edad de la planta.
Calculo de coeficientes
Aunque uno de los propósitos del trabajo fue validar el sistema de la escala poblacional bajo las condiciones de CIAT, se hicieron modificaciones metodológicas que no permiten compararlos. En síntesis, mientras el objetivo del estudio de Yaninek et al. (1989) fue el de seleccionar una hoja que pudiera ser usada como indicador o estimativo de la densidad de Mononychellus en toda la planta, en este trabajo se calculó un coeficiente para cada nivel de hojas a lo largo de la planta, o sea que para cada nivel de la planta se tiene un coeficiente diferente, es decir se logra una estimación de la población para cada nivel en cada grado de la escala.
Por otra parte, en este trabajo, como se apreciará más adelante cuando se presenten los resultados del segundo ciclo, se obtuvieron coeficientes diferentes para cada una de las especies de Mononychellus que se encontraron.
En el primer ciclo, al relacionar la escala poblacional con el conteo directo en el laboratorio se pudo observar que para los tres primeros grados de la escala se presentaron suficientes datos, lo cuál permitió calcular coeficientes de confiabilidad. Esto se puede explicar, posiblemente, por la capacidad de incremento de esta especie que no alcanza niveles muy altos.
Lo anterior se confirma con los datos obtenidos para el grado 4 de la escala. En este caso, el número de observaciones fue insuficiente, es decir, en muy pocos casos se encontraron hojas con poblaciones tan altas que permitieran dar una clasificación de 4 (más de 200 estados móviles por hoja).
De acuerdo con estos resultados se puede afirmar que M. mcgregori es una especie que no alcanza altas densidades de población y por tanto la escala no se puede utilizar con sus cuatro grados.
Es importante resaltar que lo expresado anteriormente es válido hasta la hoja número 30, ya que de la hoja 31 hasta la 60 los coeficientes no arrojan ninguna confiabilidad, pues aunque se presentan suficientes datos, la información es inestable. Esto se puede explicar por razones de tipo fisiológico, o sea por el deterioro de las hojas que no permite hacer una evaluación acertada o posiblemente también por la dispersión que presentan los ácaros en los estratos basales.
En la Tabla 1 se presentan los resultados por nivel de la planta, incluyendo la escala y la población correspondiente a cada grado, el número de observaciones para cada caso y el coeficiente calculado con su respectivo error estándar.
Relación entre la escala poblacional del complejo Mononychellus en campo y el conteo directo en laboratorio. Población de estados móviles (larvas, ninfas y adultos) para 6 niveles de la planta de yuca. Primer ciclo. Lote Punta de Lanza.
Distribución vertical de M. mcgregori
En la Figura 2 se muestra la fluctuación de la población total de los estados móviles de M. mcgregori durante el primer ciclo del cultivo. Como se puede observar, las menores poblaciones coinciden con los mayores valores de precipitación acumulada (Fig. 3), y la población tiende a aumentar durante el período seco, alcanzado algunos picos poblaciones al final del cultivo. Es interesante también resaltar que casi durante todo el desarrollo del cultivo, la población de esta especie se presentó en niveles muy bajos, posiblemente debido a factores climáticos como se dijo antes, o a la capacidad innata de crecimiento de la especie, o quizás a la presencia de insectos y ácaros fitoseiidos depredadores.
Corroborando lo dicho anteriormente, se puede apreciar en la Figura 2 que M. mcgregori se encuentra localizado a lo largo de toda la planta, pero las mayores poblaciones se concentran en los niveles superior y medio.
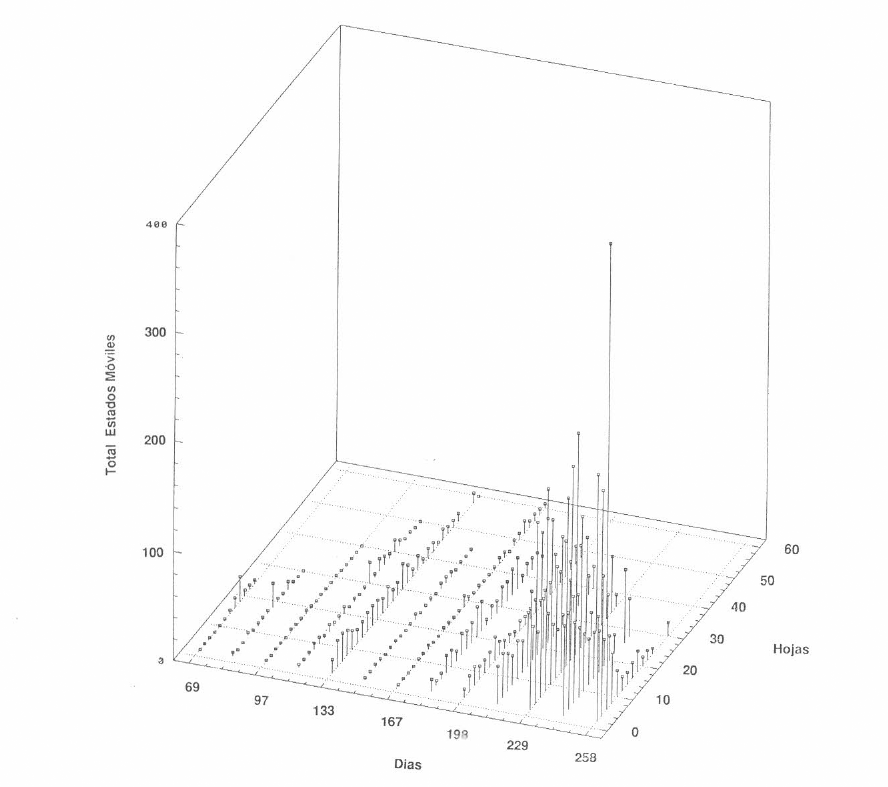
Distribución vertical de los estados móviles del complejo Mononychellus en el primer ciclo de cultivo (Lote Punta de Lanza)
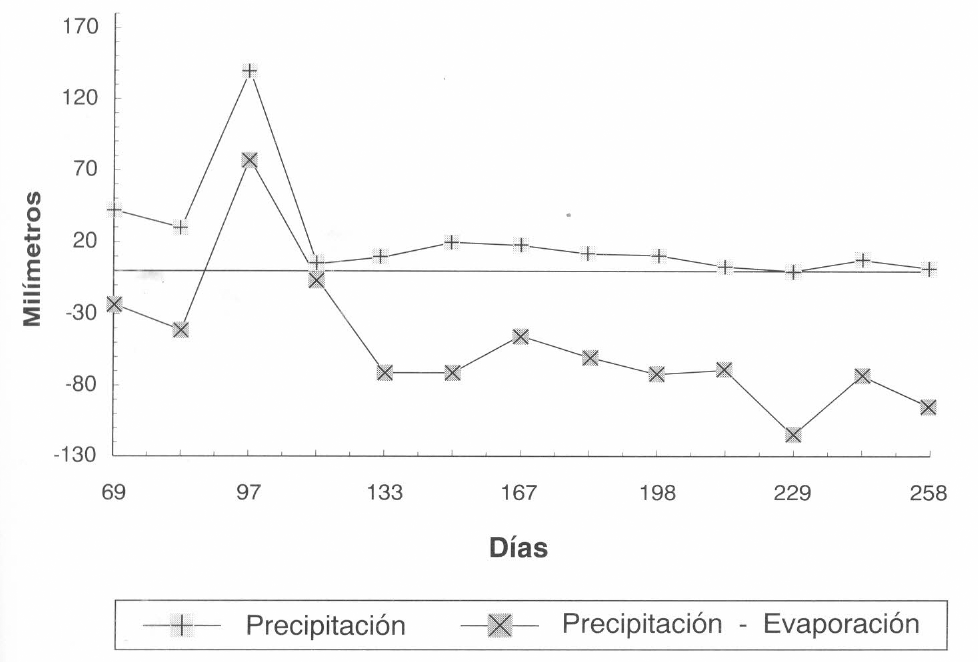
Información meteorológica en el primer ciclo de cultivo (Lote de Punta de Lanza).
Distribución vertical de la población de Phytoseiidae
Las poblaciones de las especies de Phytoseiidae en este primer ciclo, al igual que la de los ácaros fitófagos, se distribuyeron en toda la planta, observándose la misma tendencia en relación con su concentración, es decir en los niveles superior y medio. Es interesante observar que aunque no se obtuvieron números muy altos de fitoseiidos por hoja, estos ácaros siempre estuvieron presentes durante el desarrollo de este ciclo.
Si se relaciona la gráfica de distribución de M. mcgregori (Fig. 2), en la cual el fitófago estuvo en niveles muy bajos, con la frecuencia de los fitoseiidos (Fig. 4), se podría afirmar que posiblemente estos enemigos naturales ejercieron un buen control sobre la población del fitófago, o por el contrario que los fitoseiidos siempre se presentaron cuando la densidad de la presa fue baja, ya que pueden hacer uso de otras fuentes de alimento.
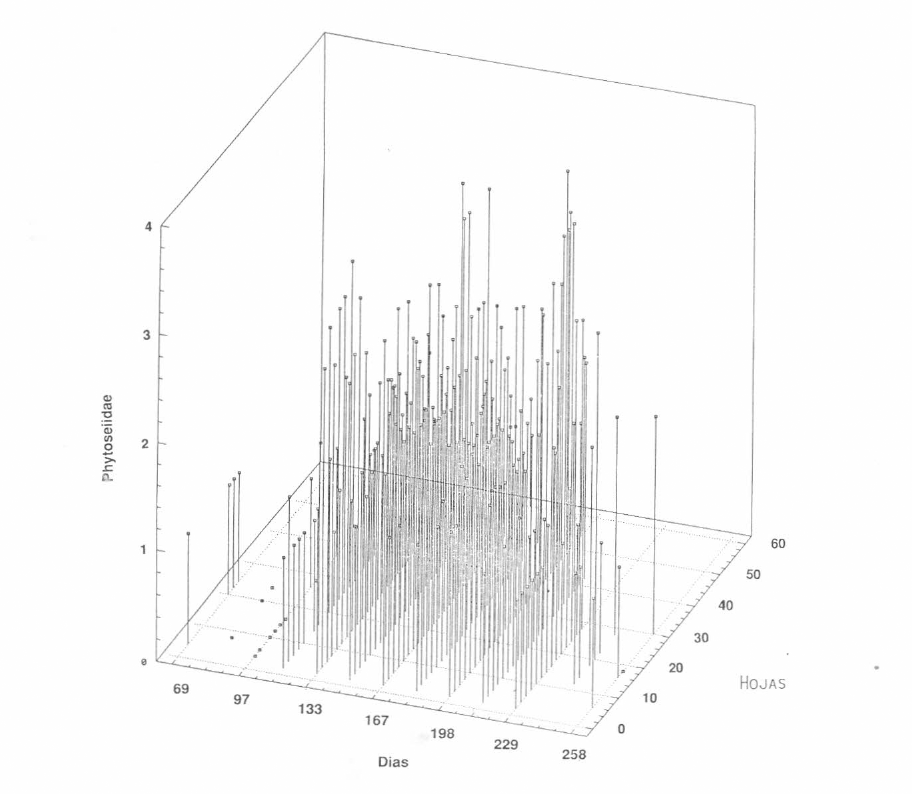
Distribución vertical de la población de los Phytoseiidae en el primer ciclo de cultivo (Lote de Punta de Lanza).
Distribución vertical de la población de Oligonychus peruvianus
Con relación a la especie O. peruvianus se observó que, aunque es posible encontrarla distribuída a lo largo de toda la planta, sus mayores concentraciones se presentaron en los niveles superior y medio (Fig. 5). Durante este primer ciclo no se registró una gran cantidad de telarañas infestadas por muestreo y el mayor valor promedio obtenido fue de 16 telarañas por hoja. Es interesante también resaltar que las poblaciones de O. peruvianos aumentaron inmediatamente después de que finalizó la época de lluvia.
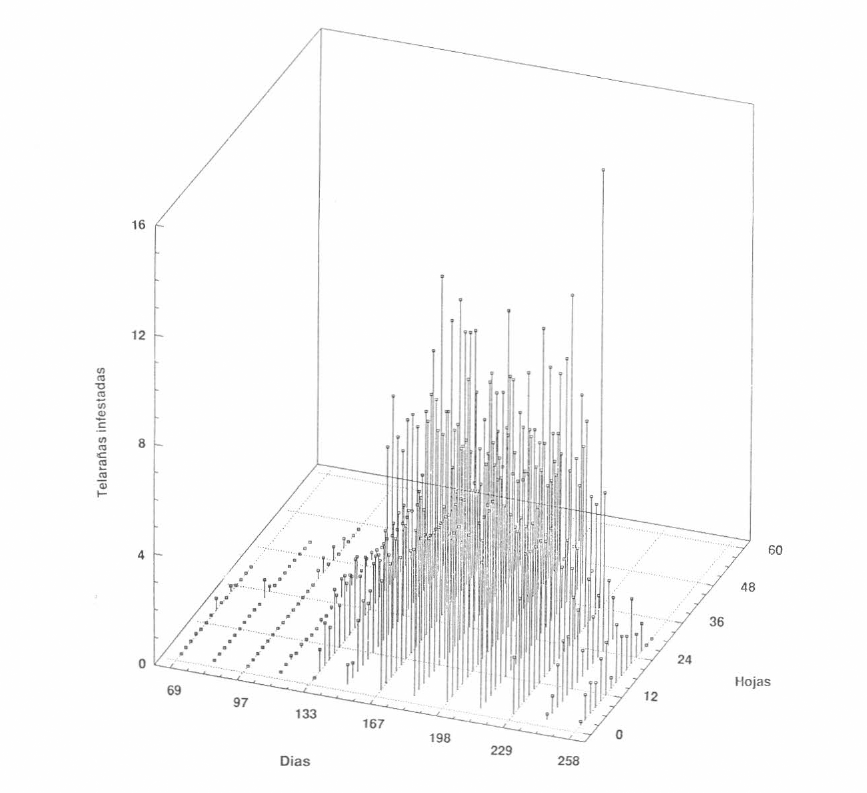
Distribución vertical de la población de Oligonychus peruvianus. Telarañas infestadas en el primer ciclo de cultivo (Lote Punta de Lanza)
Distribución de edades de M. mcgregori
En la Figura 6 se presenta la distribución de los estados de desarrollo del complejo Mononychellus en toda la planta durante el primer ciclo de muestreo. Con relación a la distribución de edades, expresado en porcentaje, se observa que en todas las fechas de muestreo se encontraron todos los estados móviles de desarrollo (larvas, ninfas y adultos); sin embargo, hace falta hacer un análisis por hoja o por nivel, para poder precisar en qué forma se concentran los diferentes estados de desarrollo en la planta, es decir, si los estados adultos y ninfas se desplazan hacia la parte terminal buscando una forma de dispersión y la otra parte de la población, hembras que están ovipositando, permanecen en las hojas más desarrolladas.
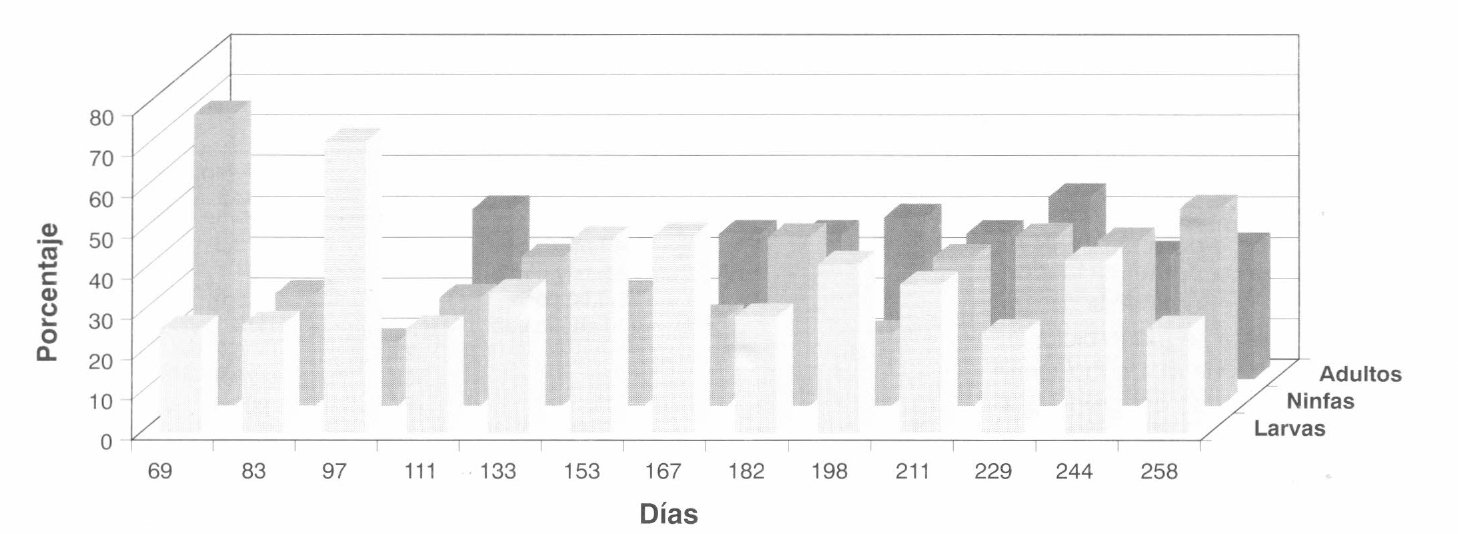
Distribución de edades (en porcentaje) del complejo Mononychellus durante el primer ciclo del cultivo (Lote Punta de Lanza).
Correspondencia entre la Presencia-Ausencia de las poblaciones de Phytoseiidae
De este tipo de muestreo se pueden esperar dos tipos de situaciones:
Un nivel de acierto que se presenta en dos formas.
Se encontraron fitoseiidos en el campo y se colectaron en su totalidad.
Simplemente no se presentaron fitoseiidos y lógicamente no se colectó nada.
Un nivel de error que se presenta así:
Error parcial. Existe población en el campo, pero no es totalmente colectada, ya que los ácaros se dispersan y no se ven.
Error total. No se observan en el campo y cuando se observan en el laboratorio sí se encuentran fitoseiidos.
En la Tabla 2, para el primer ciclo del cultivo se obtuvo un 78,05% de acierto, contra un 21,9% de error, lo cual indica que el método funcionó, ya que fue mayor el porcentaje de éxito.
Correspondencia entre la Presencia-Ausencia de poblaciones de Phytoseiidae en campo y laboratorio. Primer ciclo (Lote Punta de Lanza).
Los casos en los cuales se cometieron errores se pueden explicar por el tamaño reducido de algunas especies, por la velocidad a la cual se desplazan, o por el reflejo del sol que no permite visualizarlos bien.
Segundo ciclo de cultivo-Lote Gallinero
En la Figura 7 se presentan los porcentajes en los cuales se encontraron las diferentes especies de ácaros en los diferentes niveles de la planta, durante el segundo ciclo. En este ciclo se encontró que la especie M. caribbeanae (McGregor) predominó dentro del complejo que normalmente se presenta en CIAT. La presencia de esta especie en forma tan abundante se empezó a registrar en los lotes de CIAT a partir de este trabajo, desplazando a otras especies.
Tal como en el caso de M. mcgregori, la población de M. caribbeanae se concentra en los niveles superior y medio de las plantas, y acompañado esta especie se presentaron M. tanajoa y T. urticae en bajas densidades; durante este ciclo del cultivo no se presentó O. peruvianus, posiblemente por el desplazamiento que hizo M. caribbeanae sobre el complejo de Tetranychidae nativos.
En relación con el complejo de los depredadores, las especies Neoseilus anonymus (Chant & Baker) y T. limonicus fueron las más frecuentes y también en los niveles superior y medio. Durante este ciclo se destacó la presencia abundante de T. aripo de León en los cogollos y en los niveles superiores. La especie E. naindaimei se registró sólo en el nivel superior y en forma muy esporádica.
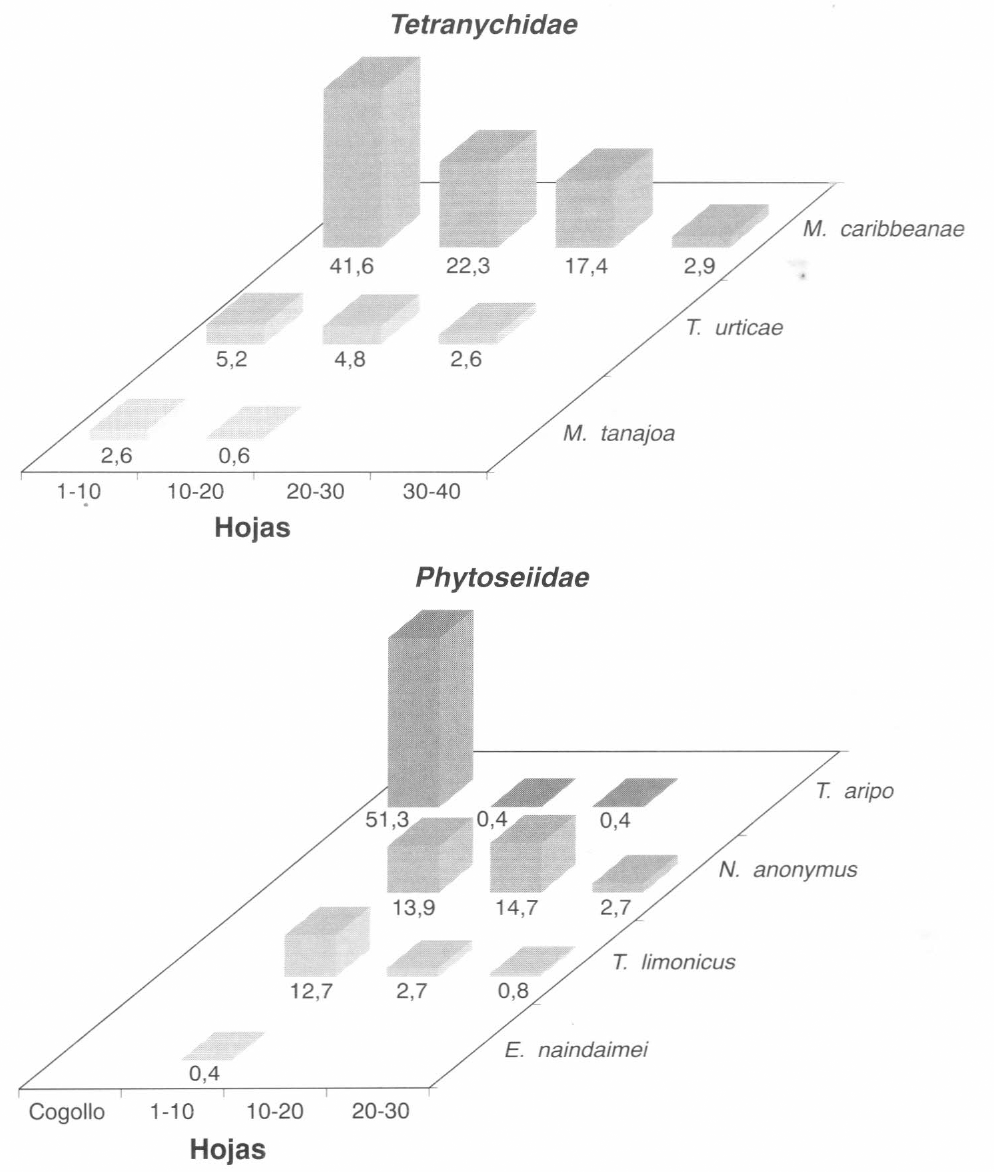
Distribución vertical de las especies de Tetranychidae y Phytoseiidae durante el desarrollo del segundo ciclo del cultivo (Lote Gallinero).
Calculo de coeficientes
Los coeficientes calculados son básicamente para M. caribbeanae. En este ciclo de cultivo se obtuvo suficiente información (número de datos) para realizar el cálculo de los coeficientes en los 4 grados de la escala, en forma confiable, hasta la hoja número 20, pues de la hoja 21 a la 40 esta información no fue suficiente, debido posiblemente a las razones de tipo fisiológico dadas para el primer ciclo.
En contraste con el primer ciclo, donde la especie predominante fue M. mcgregori, en este segundo ciclo, las poblaciones altas de M. caribbeanae permitieron hacer uso del grado cuatro de la escala; esto posiblemente se debe a su tasa de incremento que es mucho mayor y, probablemente, a su capacidad de adaptación a las condiciones del CIAT.
Es interesante resaltar que las plantas de este segundo ciclo, posiblemente por haber soportado poblaciones más altas de M. caribbeanae, no alcanzaron mucha altura y el número de hojas en promedio llegó hasta 40.
En la Tabla 3 se presenta la relación entre la escala de población y el conteo directo en el laboratorio, y como en el ciclo anterior, se calcularon los coeficientes para cada grado de la escala y para los niveles de hojas.
Relación entre la escala poblacional del complejo Mononychellus y el conteo directo en laboratorio. Población de estados móviles (larvas, ninfas y adultos) para cuatro niveles de la planta de yuca. Segundo ciclo. Lote Gallinero.
Distribución vertical de M. caribbeanae (estados móviles)
En la Figura 8 se presenta la distribución de los estados móviles de M. caribbeanae a lo largo de la planta durante todos los muestreos. Como se puede observar, la población se concentra hacia la parte superior de la planta. En este ciclo se alcanzan promedios mucho más altos de ácaros por hoja (más de 600 estados móviles). Aunque al inicio del ciclo, en los primeros muestreos no se presentaron altas poblaciones, después de realizar la infestación, la población se dispara y permanece durante todo el ciclo.
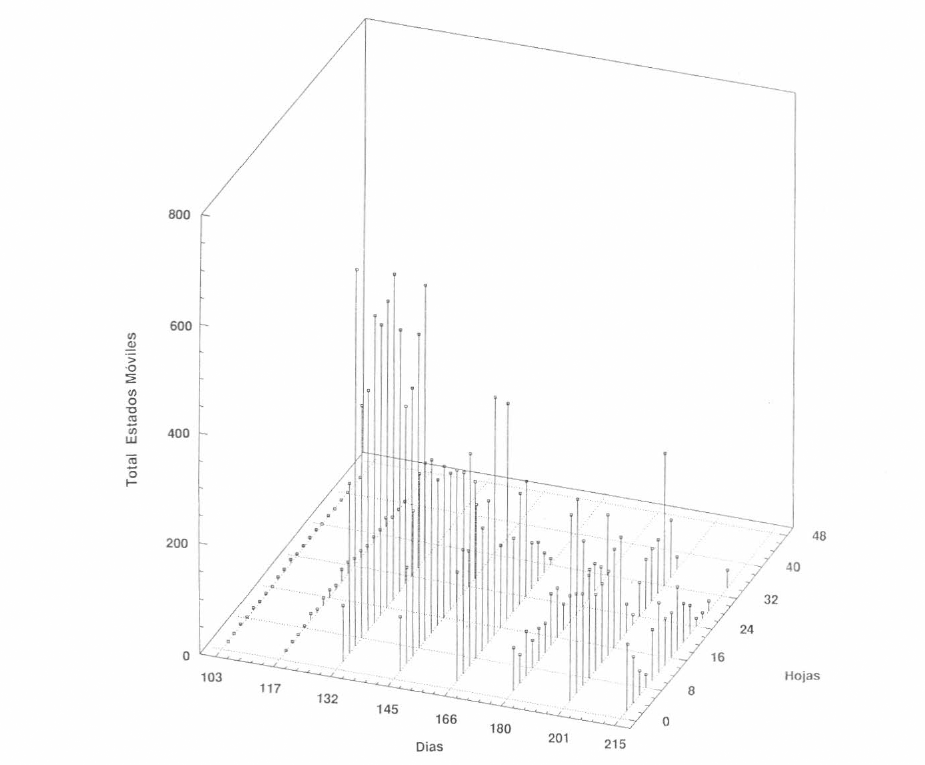
Distribución vertical de los estados móviles del complejo Mononychellus en el segundo ciclo del cultivo (Lote Gallinero).
Al relacionar esta fluctuación de la población con la precipitación acumulada (Fig. 9), se observó que aunque el experimento estuvo sometido a mayores precipitaciones, las poblaciones de M. caribbeanae siempre permanecieron. Esto corrobora lo expresado sobre la capacidad de adaptación de la especie y su grado de agresividad.
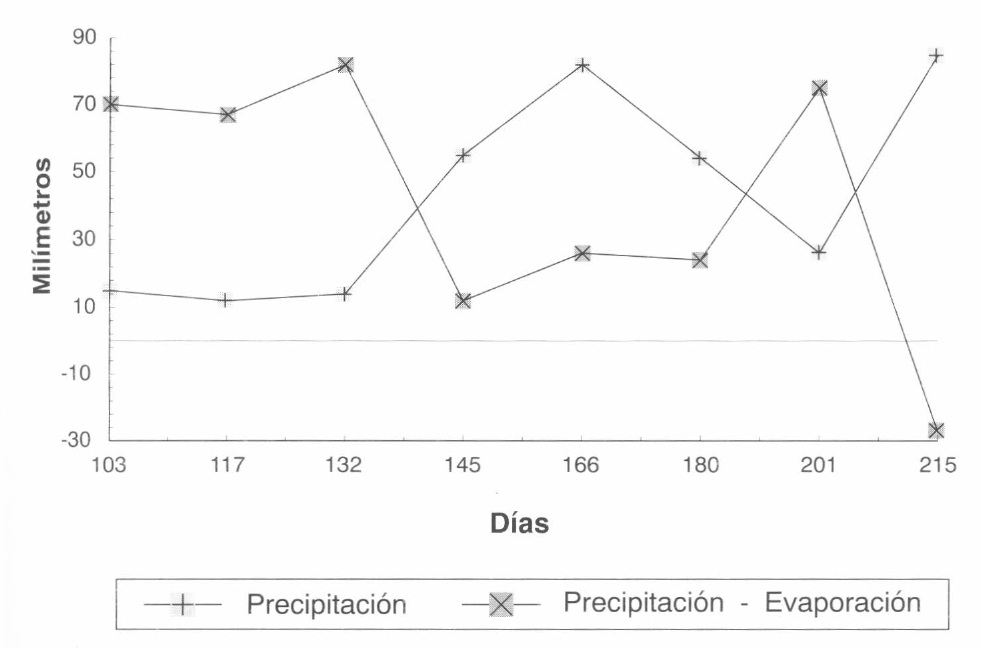
Información meteorológica en el segundo ciclo del cultivo (Lote Gallinero).
Distribución vertical de la población de los Phytoseiidae
En contraste con el primer ciclo, en el segundo, la población de fitoseiidos sólo se presentó en forma abundante en el último muestreo, y durante el desarrollo del cultivo las apariciones fueron esporádicas (Fig. 10). En este ciclo aparecieron dos especies diferentes en mayor proporción: N. anonymus y T. aripo; además es interesante tener en cuenta que durante este ciclo, en el cual la población del fitófago fue más abundante, las poblaciones de los depredadores fueron mínimas.
Especulando un poco sobre esta interacción, se podría pensar que el complejo de fitoseiidos presentes en CIAT no ha coevolucionado con M. caribbeanae y por esa razón no alcanzan altas densidades, aunque tengan abundante alimento. También vale la pena resaltar que los fitoseiidos en este segundo ciclo mostraron el mayor pico en población y en diversidad, cuando las poblaciones del fitófago estaban decreciendo, y hacia el final del cultivo.
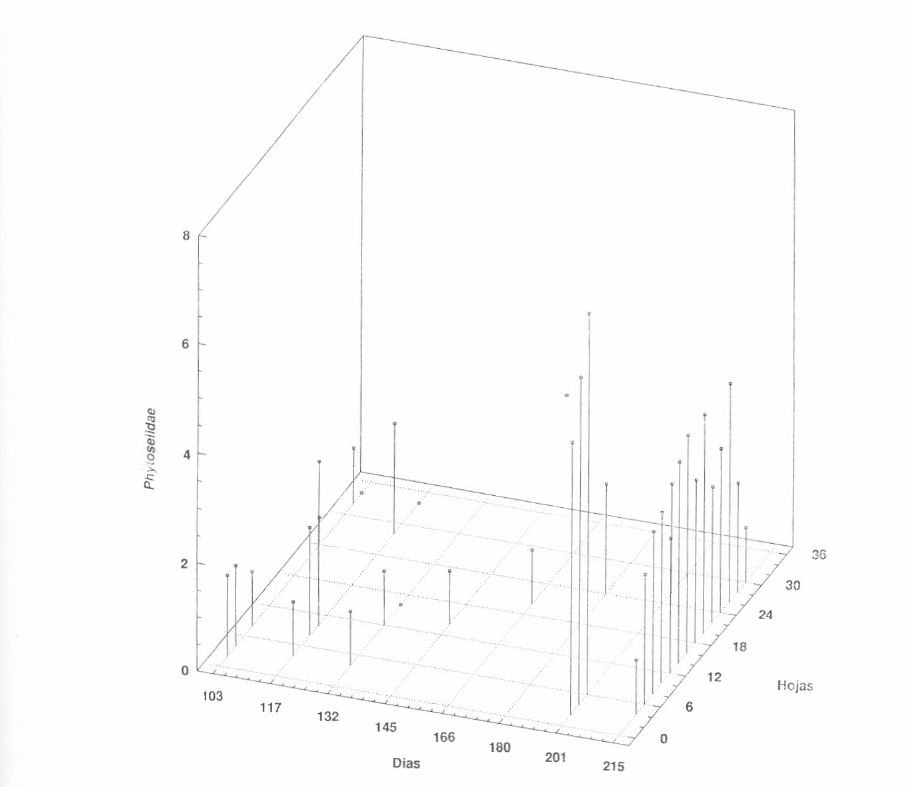
Distribución vertical de la población de los Phytoseiidae en el segundo ciclo de cultivo (Lote Gallinero).
Correspondencia entre la Presencia-Ausencia de las poblaciones de Phytoseiidae
Los resultados para el segundo ciclo se presentan en la Tabla 4. Se pudo establecer un 91,26% de acierto frente a un 8,7% de error en este segundo ciclo. Este alto porcentaje de acierto se puede explicar por la menor densidad de población de fitoseiidos encontrada, es decir, hubo menos posibilidad de equívoco en la evaluación.
Correspondencia entre la Presencia-Ausencia de poblaciones de Phytoseiidae en campo y laboratorio, segundo ciclo (Lote Gallinero).
Conclusiones
El complejo Mononychellus, representado por las especies M. mcgregori, M. tanajoa y M. caribbeanae, estuvo presente en los dos ciclos del cultivo, distribuyéndose sus poblaciones a lo largo de toda la planta y concentrándose principalmente hacia la parte alta y media.
El 88% de la población de los Tetranychidae registrados en el primer ciclo del cultivo, correspondió a la especie M. mcgregori, mientras que en el complejo de los fitoseiidos estuvo representado en el 89,2% de los casos por T. limonicus.
En el segundo ciclo del cultivo, la especie fitófaga predominante fue M. caribbeanae (83,2%), asociada con los depredadores T. aripo, principalmente en el cogollo (52%), у M. anonymus, distribuido en toda la planta (32,3%).
Se establecieron coeficientes de correspondencia entre las escalas poblaciones y el conteo directo para las especies M. mcgregori y M. caribbeanae en la variedad de yuca CMC-40, para las condiciones del CIAT-Palmira.
En cuanto al uso de la escala, se observó que está directamente relacionado con la especie de Mononychellus presente. Las dos especies estudiadas tienen tasas de incremento y patrones de distribución espacial diferentes. La confiabilidad de la escala fue válida en los tres primeros niveles de la planta, es decir de la hoja 1 a la 30, lo cual coincide con la mayor concentración de la población. Para M. mcgregori la escala fue confiable hasta el grado 3, mientras que para M. caribbeanae funcionó hasta el grado 4.
El porcentaje de error para la evaluación del método Presencia-Ausencia en la valoración de ácaros fitoseiidos, en condiciones de campo y laboratorio, fue de 22 y 8,8%, para el primero y segundo ciclos, respectivamente.
Realmente, utilizar la escala poblacional trae beneficios como es el ahorro de tiempo. En un muestreo directo de 20 plantas los conteos directos requirieron 96 horas/hombre, mientras que con estas mismas plantas evaluadas con escalas poblacionales se necesitaron 12 horas/hombre, es decir, alrededor de 1:8.