
Abstract
Introduccion
La resistencia animal a la infestación por garrapatas tiene bases inmunes y genéticas. Hoy se indica con evidencia que los componentes salivares de la garrapata son importantes en la estimulación del sistema inmune del huésped (Wikel y Allen 1982). Por ésta razón, las glándulas salivares de la garrapata y sus secreciones son una fuente importante de material antigénico para los estudios modernos en este campo.
La mayoría de estudios sobre resistencia, realizados hasta ahora, han usado extractos de glándulas salivares o extractos de garrapatas completas (Wikel et al. 1978; Willadsen 1980), mientras que los estudios de intoxicación por garrapatas, transmisión de enfermedades y fisiología de artrópodos se han basado en saliva obtenida artificialmente mediante el uso de drogas parasimpaticomiméticas que estimulan la salivación (Purnell et al. 1969; Tatchell 1967; Gregson 1973). La saliva obtenida de esta manera es extremadamente diluida y no es necesariamente el equivalente a saliva producida naturalmente (Binnington y Kemp 1980).
La secreción natural de saliva puede lograrse mediante estimulación táctil del hipostoma, colocando un tubo capilar sobre él (Gregson 1973), pero a pesar de haber sido ampliamente usado en estudios de intoxicación por garrapatas, este método es muy laborioso y sólo es posible obtener pequeñas cantidades de saliva.
El propósito de este trabajo fue intentar inducir la secreción natural de saliva en hembras de la garrapata
Revision De Literatura
Adherencia y Alimentación de la garrapata.
Las glándulas salivares de la garrapata son un par de estructuras con apariencia de racimo de uvas, localizadas lateralmente en el hemocelo, que se extienden hasta la altura de los espiráculos (Till 1961). En las hembras se presentan tres tipos de acinos glandulares: Acino tipo ló piramidal y acinos granulares tipo II y III (Batungbacal 1974; Binnington 1978). En los machos existe un acino adicional tipo IV.
Se ha demostrado que existen siete tipos de células granulares en el macho de
Una vez que la garrapata encuentra un sitio apropiado para su adherencia, penetra la piel del huésped mediante movimientos de los quelíceros (Gregson 1973), esto es seguido por la secreción de cemento en los siguientes diez minutos (Kemp et al. 1982). Entonces, rápidamente se inicia un proceso de ingestión de fluidos y salivación. Este patrón es más complejo que las simples alteraciones entre ingestión de sangre y salivación. Los procesos son discretos inicialmente, para luego incrementarse en frecuencia con períodos largos de secreción de saliva (Gregson 1969; Arthur 1970). La salivación más copiosa ocurre hacia el final del proceso de alimentación y regresa al huésped excesos de agua, concentrando la sangre ingerida (Kemp et al. 1982).
Se ha demostrado que las diferencias en los patrones de ingestión y salivación están relacionadas con la composición química del medio ofrecido bajo membranas artificiales. Esta respuesta parece ser debida a la presencia de quimiorreceptores en las piezas bucales de la garrapata (Waladde y Rice 1982). Así mismo, Waladde et al. (1979) demostraron que los fagoestimulantes trifosfato de adenosina (ATP) y glutatión reducido (GSH) incrementaron la frecuencia del patrón simple de alimentación en
Estudios histoquímicos de las glándulas salivares de la garrapata y de la saliva han demostrado que estas secreciones contienen enzimas hidolíticas débiles, como esterasas y aminopeptidasas y agentes farmacológicos, como prostaglandinas y antagonistas de la histamina. Sin embargo, su papel en el proceso de alimentación de la garrapata no ha sido completamente estudiado (Schleger y Lincoln 1976; Binnington y Kemp 1980). Las glándulas salivares intervienen además en la regulación de iones y en la eliminación de excesos de agua (Tatchell 1969; Kaufman y Sauer 1982).
Métodos artificiales de alimentación de garrapatas y técnicas para obtener su saliva.
Los métodos artificiales para la alimentación de garrapatas han sido ampliamente usados. Para argásidos, Edelsten (1974) describió métodos que utilizan la membrana corioalantoidea de huevos embrionados. Para ixódidos, Gregson (1973) logró inducir la adherencia de hembras de
El método de tubos capilares ha sido principalmente usado en estudios de transmisión de enfermedades. Se ha indicado que las garrapatas sólo ingieren la sangre ofrecida en los capilares por un tiempo limitado (Gregson 1973). La garrapata
Se ha concluido que la salivación ocurre en los tubos capilares de manera suficiente, como para recolectar partículas infectivas de
En cuanto a la obtención de saliva, Gregson (1957) encontró que las garrapatas podrían ser inducidas a salivar, si un tubo capilar se colocaba cuidadosamente sobre el hipostoma. De acuerdo con Tatchell (1967), para esta técnica es crítico que el diámetro interno del tubo comprima los quelíceros.
El uso de drogas parasimpaticomiméticas para obtener secreciones salivares ha sido ampliamente descrito. Se ha reportado que inyecciones de 5-10 microlitros de pilocarpina al 10% en salina, causan un incremento en seis veces la secreción de saliva en
Estudios fisiológicos de las glándulas salivares de la garrapata han demostrado que existe un control neural de la salivación (Kaufman y Phillips 1973), pero ésta puede ser también estimulada por la inyección de líquidos isotónicos en la hemolinfa, tales como cloruro de sodio, sucrosa, urea o agua destilada (Kaufman y Sauer 1982).
Materiales Y Metodos
Alimentación artificial
Las hembras de
Tubos capilares de vidrio de 75 mm de largo y 0,8-0,9 mm de diámetro interno fueron moldeados al calor para reducir el diámetro de los extremos a 0,2-0,5 mm, con el fin de adaptarlos a las piezas bucales de la garrapata (Burgdorfer 1956; Tatchell 1967; Gregson 1973).
Para los medios artificiales se utilizó la solución balanceada de sales de Hank (Gibco, Europa), a la que se le adicionaron tres concentraciones diferentes de Albúmina Plasmática Bovina (BPA). De acuerdo con la concentración de proteína utilizada, los medios se denominaron de la siguiente manera: HBSS (Sin proteína), HO,5PA (0,5% de proteína) y H5% BPA (5% de proteína). El pH de los medios se ajustó a 7,2, mediante la adición de bicarbonato de sodio.
Los medios se esterilizaron mediante la filtración a través de un filtro Swinnex con una membrana de acetato de celulosa de un tamaño de poro de 0,22 µm y se repartieron en alícuotas de 5 ml, las cuales se conservaron congeladas a -20°C. Previamente a su uso, los medios fueron descongelados y usados a temperatura ambiente en los experimentos. Como fagoestimulantes en los medios se utilizó el Glutatión reducido (GSH) (Sigma Labs.) a la concentración de 10−2 M.
Para la alimentación artificial, las garrapatas fueron adheridas ventralmente a un soporte Perpex especialmente diseñado, en donde los tubos capilares que contenían el medio respectivo se colocaron cuidadosamente sobre las piezas bucales de la garrapata (Fig. 1). En cada experimento se trabajó con grupos de 10 garrapatas.

Alimentación artificial de garrapatas con un medio contenido en tubos capilares. Observe como el extremo del tubo encaja cpn el hipostoma que penetra en él.
El peso individual de cada garrapata se registró utilizando una balanza Oerteling (R.20, d=0,1 mg). Así mismo, se anotó el nivel de la columna de fluido en los tubos capilares, antes y después del proceso de alimentación. Este período fue generalmente de 4-5 horas, durante las cuales las garrapatas se colocaron en el interior de una cámara con una humedad relativa del 100% dentro de un incubador a 37°C. Además, se realizaron algunas pocas observaciones con períodos de 14 horas de alimentación.
Luego del período de alimentación artificial, el medio remanente en los tubos capilares se recolectó en recipientes plásticos de acuerdo con el grupo experimental y se conservó congelado a -20°C.
Se realizaron dos grupos de experimentos utilizando garrapatas prealimentadas en conejos sin previa exposición a garrapatas. El primer grupo incluyó la comparación de tres diferentes niveles de albúmina en el medio (0, 0,5% y 5%) con la inclusión o no de GSH como fagoestimulante.
El segundo grupo consistió en comparar garrapatas alimentadas en el medio sin proteína y con 5% de ella, pero sin utilizar fagoestimulantes. Además, en algunas replicaciones se utilizaron garrapatas prealimentadas en conejos que habían sido previamente expuestos a la garrapata
Como estimulante de la secreción salivar se utilizó dopamina. En dos ocasiones las garrapatas se inyectaron con una solución de dopamina 10−2M en PBS. La inyección se efectuó en el centro del dorso de las garrapatas, aplicando 2-10 µl de solución por espécimen, a intervalos de 15-20 minutos (Kaufman y Sauer 1982; Walker et al. 1979). La saliva producida se colectó mediante la colocación de tubos capilares sobre las piezas bucales de la garrapata, 15-20 minutos después de la segunda inyección. La saliva se recolectó en tubos Eppendorf y se conservó congelada a -20°C.
Detección de componentes salivares en el medio después de la alimentación artificial
Con el fin de detectar material antigénico que pueda permanecer en el medio después de la alimentación de garrapatas, se utilizó una prueba de hipersensibilidad en la piel de conejos sensibilizados a la garrapata. En ellos se comparó la reactividad dérmica producida por los diferentes medios luego de la alimentación de garrapatas en ellos, contra la reactividad producida por la inyección de saliva obtenida por la estimulación con dopamina.
A cuatro conejos Nueva Zelanda se les permitió desarrollar inmunidad a la garrapata por medio de tres infestaciones con adultos machos y hembras, utilizando un esquema similar al descrito por Wikel et al. (1978), con períodos de reposo de 7 días entre una y otra infestación. La prueba dérmica se realizó a los 12 y 13 días de finalizada la tercera infestación.
Los cuatro conejos inmunizados y uno sin exposición previa a las garrapatas fueron rasurados en el dorso y el área de piel dividida en cuadros de aproximadamente 4 cm X 4 cm con la ayuda de un marcador indeleble. En cada conejo se inyectó intradérmicamente 0,1 ml de las diluciones del material a probar. Las reacciones de hipersensibilidad inmediata producidas por estos materiales se valoraron por el tamaño de los diámetros de la reacción de edema producida dos horas después de la inyección. El producto de dos mediciones en ángulo recto se tomó como representativo del área de reacción (Willadsen et al. 1978).
Con el fin de detectar reacciones de hipersensibilidad retardada, se realizaron lecturas adicionales a las 24 y 48 horas post-inoculación, registrando en este caso el diámetro del área de eritema e induración (Roitt 1980). Los medios, así como la saliva obtenida mediante estimulación por dopamina, fueron ensayados si diluir y en diluciones de 1/10 y 1/ 100 en PBS.
Resultados Y Discusion
Cambios de peso en garrapatas alimentadas artificialmente en diferentes medios
La habilidad de las garrapatas para ingerir el medio ofrecido en los tubos capilares, se midió por el aumento de peso registrado durante el período de alimentación. Sin embargo, no todas las garrapatas se alimentaron exitosamente y los valores obtenidos mostraron gran dispersión y ausencia de normalidad en los datos crudos. Así, los valores negativos fueron eliminados y los datos sometidos a transformación logarítmica (log10[x=1]), previo al análisis de varianza (Sard 1978). El efecto de cada tratamiento en el éxito para incrementar el peso se evaluó mediante una prueba de chi-cuadrado (Bailey 1981).
La Figura 2 ilustra los resultados obtenidos en el primer experimento, al comparar tres niveles de proteína en el medio y la adición de GSH como fagoestimulante. El incremento de peso de las garrapatas alimentadas en medio que contenía 5% de proteína fue diferente (P<0,01) del incremento logrado en las garrapatas alimentadas en medios que contenían 0,5% ó sin proteína. La adición de GSH no demostró efecto significativo sobre estos incrementos de peso.
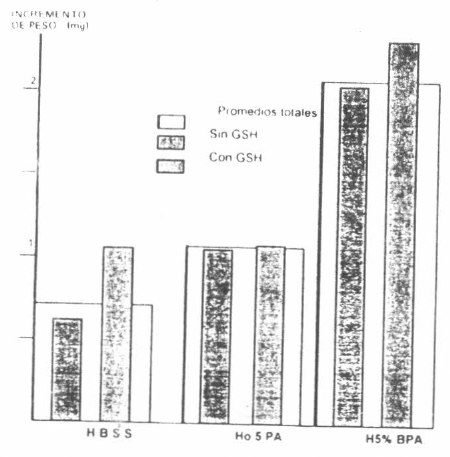
Efecto de tres concentraciones de proteína en el medio y la adición de GSH, sobre el incremento de peso de garrapatas alimentadas artificialmente en tubos capilares. La diferencia en H5% BPA es altamente significativa, (P<0,01). La adición de GSH no demostró efectos significativos.
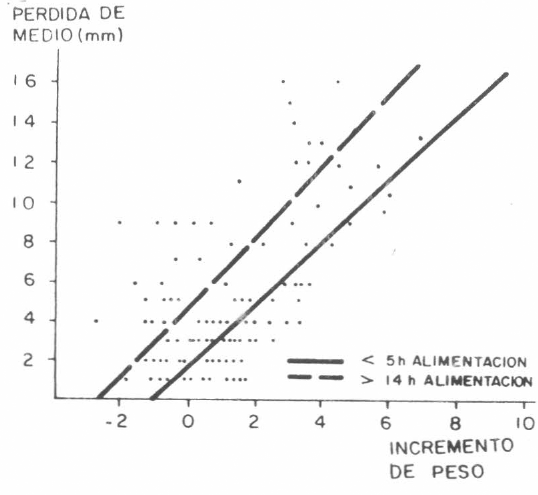
Correlación entre el incremento de pesos y la pérdida de medio en los tubos capilares en garrapatas alimentadas artificialmente por períodos de menos de 5 horas (r=0,877) o más de 14 horas (r=0,863). Las pendientes son similares pero difieren (P<0,01) en nivel.
La calibración volumétrica de los tubos capilares demostró que 1 mm de reducción en la columna de fluido, representaba un volumen de 0,726 µl. El análisis de la relación entre el aumento de peso y la reducción de la columna de fluido (Fig.3) mostró una correlación altamente significativa en todos los tratamientos. Esto indica que la pérdida de líquido en los tubos fue causada por la ingestión por parte de las garrapatas y que la evaporación o escape no fueron factores importantes en la variación.
Durante el segundo grupo de experimentos, al analizar las diferencias entre garrapatas alimentadas en dos medios (proteína al 5% de concentración o no proteína), y utilizando garrapatas prealimen-tadas en conejos con y sin exposición previa a las garrapatas, se encontró que los mayores valores de aumento de peso se dieron en las garrapatas alimentadas en medio que contenía 5% de proteína y a su vez prealimentadas en conejos susceptibles (1,93±0,86 mg) y los menores valores se observaron en garrapatas alimentadas en medio sin proteína y prealimentadas en conejos inmunes a las garrapatas (0,39±0,18 mg).
El análisis del efecto combinado de la fuente de prealimentación de las garrapatas y el tipo de medio usado para la alimentación, se ilustra en la Figura 4. El aumento de peso en garrapatas prealimentadas en conejos susceptibles a la garrapata (1,11±0,36 mg para HBSS y 1,41±0,68 mg para H5%BPA) fue mayor (P<0,01) que el de garrapatas prealimentadas en conejos inmunes (0,64±0,37 mg para HBSS y 0,81±0,5 mg para H5%BPA).
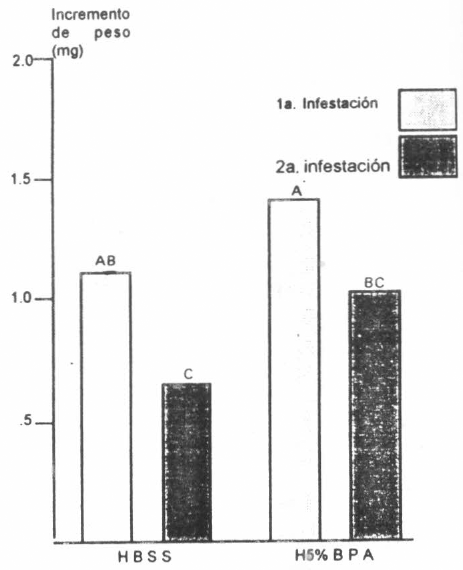
Efecto del nivel de resistencia a garrapatas de los conejos en que se prealimentaron las garrapatas, sobre el incremento de peso en hembras de
En resumen, el medio que contiene 5% de proteína demostró ser el más efectivo para una alimentación artificial exitosa de la garrapata, aunque el medio sin proteína también permite cierto grado de alimentación. Estos resultados coinciden con los reportados por Kemp et al. (1975), quienes demostraron que para alimentar larvas de
Lafalla del GSG para crear un incremento en el patrón de alimentación, puede ser explicada por los hallazgos de Waladde et al. (1979), quienes demostraron que la adición de GSH o ATP a sangre entera, incrementó la frecuencia de las fases de ingestión, pero este efecto sólo fue notorio durante los primeros 7 minutos de alimentación. Período insignificante, si se tiene en cuenta que en estos experimentos el período de alimentación fue de cuatro horas.
El menor aumento de peso observado en las garrapatas prealimentadas en conejos con exposición previa al parásito, podría explicarse por una inhibición en la ingestión de líquidos, causada por agentes farmacológicos liberados por el huésped en el sitio de adherencia (Willadsen 1980). Se ha demostrado que ocurren alteraciones en la composición de la ingesta en garrapatas que se alimentan en conejos inmunes (Wikel y Allen 1982), creando un aumento en la ingestión de leucocitos, principalmente basófilos. Su ingestión y la posterior liberación de compuestos farmacológicamente activos, explicaría la interferencia con la alimentación de la garrapata.
Detección de componentes salivares en el medio después de la alimentación artificial.
Las reacciones de hipersensibilidad inmediata se caracterizaron por el desarrollo de una reacción edematosa prominente, la cual fue notoria al momento de la lectura, dos horas luego de la inoculación (Fig. 5). Con la medición de estas reacciones se obtuvo, para cada inóculo, el promedio de los registros en los cuatro conejos inmunizados.
Reacción de hipersensibilidad inmediata a antígenos salivares de garrapata, en la piel de conejos inmunes. De izquierda a derecha se observan las reacciones producidas por saliva sin diluir, y diluciones 1/10, 1/100 y por PBS.
La Figura 6 presenta los resultados obtenidos al analizar los medios obtenidos durante la segunda fase de experimentos de alimentación artificial. Estos se presentan como regresión lineal entre el área de reacción edematosa desarrollada en los conejos y el logaritmo del recíproco de la dilución del medio utilizada (Willadsen et al. 1978).
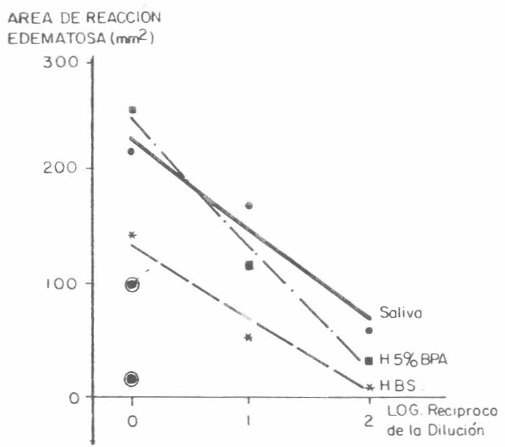
Relación entre la respuesta de hipersensibilidad inmediata y diluciones de los medios del experimento 2, o saliva. Las pendientes son similares, pero el nivel de la constante es diferente (P<0,01). Los puntos representan el promedio para cada alergeno en cada dilución (♦) saliva, (*) HBSS, (■) H5% BPA. Los puntos dentro de un círculo representan las reacciones desarrololadas en el conejo control para saliva (◊) y H5% BPA (□).
Cada regresión por sí misma fue altamente significativa. El análisis de varianza entre las diferentes regresiones demostró que la pendiente era similar, pero mostró diferencias (P<0,01) en el tamaño de la reacción producida por el antigeno sin diluir, de la siguiente manera: Saliva=226 mmβ, H5%BPA=242 mmβ y HBSS=124 mmβ. Estos resultados sugieren la presencia de componentes salivares en los medios, con una concentración mayor en H5%BPA, similar a la observada en la saliva obtenida mediante inyección de dopamina.
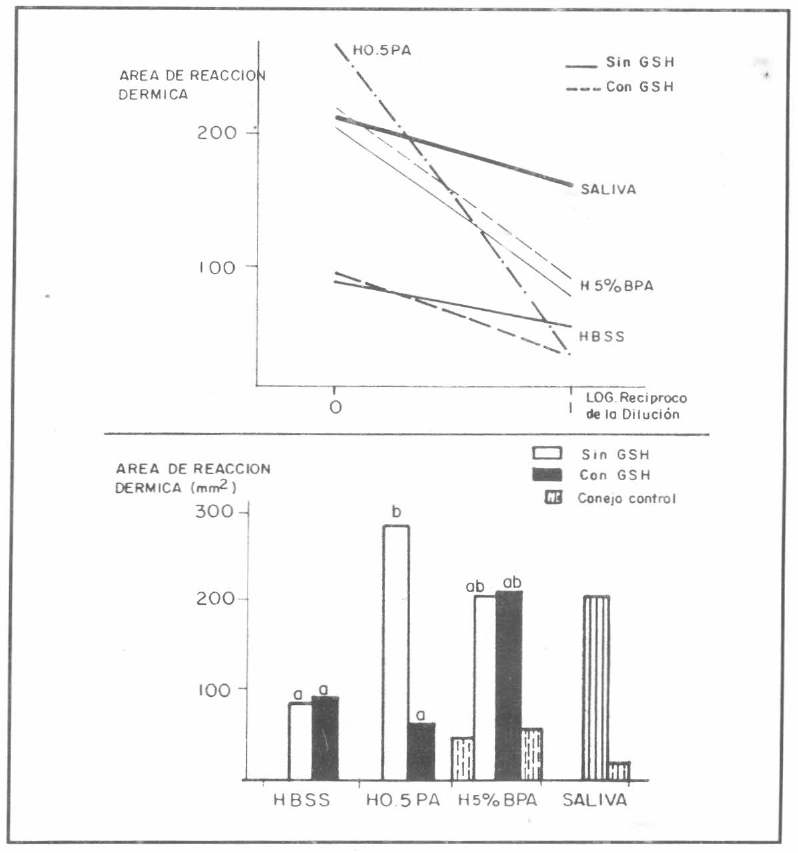
Reacciones de hipersensibilidad inmediata producidas en conejos inmunes por los medios usados para alimentar artifricialmente garrapatas en el experimento1. A. Relaciones entre dosis y área de reacción edematosa comparadas a la reacción producida por saliva obtenida por inyección de Dopamina. B. Promedio de las reacciones producidas saliva y medios, sin diluir, comparados con la reacción producida en el conejo control, hay diferencias altamente significativas entre columnas con letas diferentes.
El conejo control (no inmune) desarrolló una ligera reacción a la saliva y a H5% BPA, sugiriendo la presencia de un factor no inmune en la reacción total, como es la presencia de compuestos vasoactivos (Wikel et al. 1978).
Las reacciones producidas por los medios obtenidos del experimento 1, se ilustran en la Figura 7. Estas reacciones presentaron mayor dificultad para su interpretación, ya que sólo dos diluciones fueron evaluadas. En este caso, no todas las regresiones fueron significativas, entonces, las reacciones fueron comparadas mediante un análisis de varianza, el cual demostró diferencias significativas (P<0,05) entre los promedios para los diferentes medios. H0,5PA produjo las más fuertes reacciones (284 mmβ), nivel que no difiere del producido por saliva o H5%BPA.
Las reacciones de eritema e induración producidas por los inóculos, en general fueron más marcadas a las 24 horas post-inoculación. Con los datos obtenidos a las 24 horas se realizaron pruebas de regresión, de manera similar a como se analizaron las reacciones inmediatas. El análisis de la información procedente del experimento 1 presentó los mismos problemas que los descritos para las reacciones inmediatas.
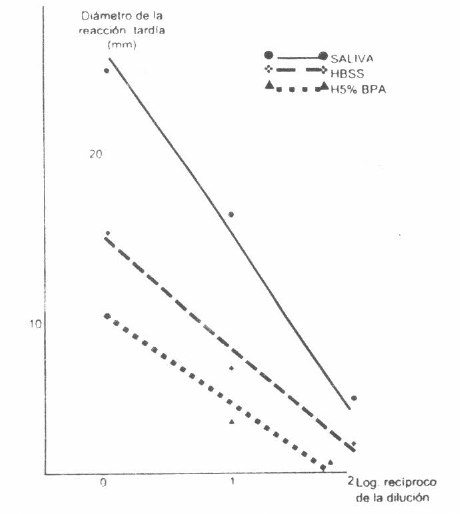
Relación entre la respuesta en la reacción de hipersensibilidad tardía, y la dilusión de saliva o medios del experimento 2. La diferencia entre pendientes es significativa (p<0,05) y la diferencia entre constantes es altamente significatavia (p<0,01). Los puntos representan los promedios para alergeno en cada dilución. El conejo control no desarrolló ninguna reacción.
Las reacciones de hipersensibilidad retardada presentaron un patrón similar al de las reacciones inmediatas (Fig. 8 para el experimento 2 y Fig. 9 para el experimento 1), pero la saliva, obtenida mediante la inyección de dopamina en las garrapatas, produjo reacciones marcadamente diferentes de las producidas por los medios. H0,5%PA produjo niveles mayores de reactividad. El conejo control no desarrolló ningún tipo de reacciones.
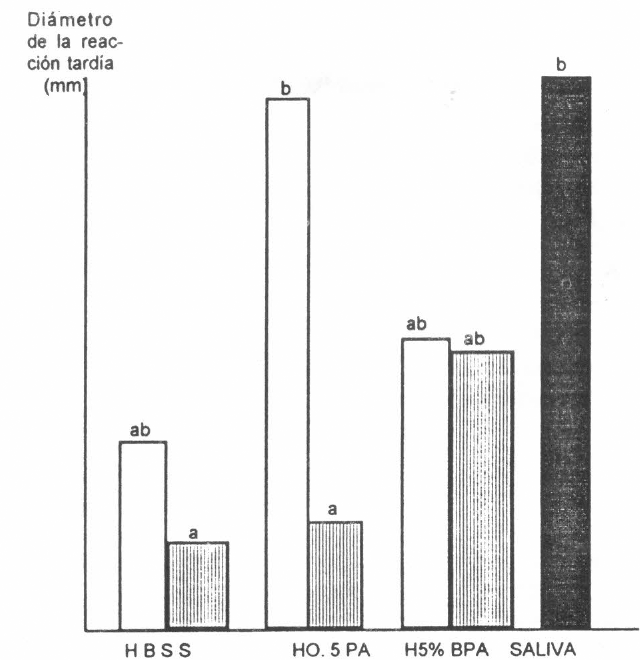
, Efecto del tipo de medio durante la alimentación artificial, sobre el nivel de reacción de hipersensibilidad retardada desarrollada en conejos sensibilizados por los alergenos sin diluir. Columnas con diferente letra son diferentes. (p<0,01). El conejo control no desarrolló ningún tipo de reacción.
Conjugando los resultados de las evaluaciones de las respuestas inmediatas y retardadas se puede afirmar que existen componentes antigénicos tanto en la saliva obtenida mediante inyección de dopamina, como en los diferentes medios en los que las garrapatas fueron alimentadas artificialmente. Estos antígenos desencadenan reacciones de hipersensibilidad inmediata y retardada en conejos inmunes. Ambos tipos de reacciones han sido descritas en animales resistentes a la garrapata (Willadsen et al 1978; Wikel et al. 1978).
Además, se demostró la presencia de factores no inmunes que forman parte de la reacción inmediata, tanto en la saliva como en algunos de los medios utilizados. Se cree que estos son compuestos vasoactivos secretados por la garrapata para facilitar su alimentación en el huésped (Wikel et al. 1978).
Conclusiones
Durante estos experimentos se demostró que:
Garrapatas hembras de
Las garrapatas tuvieron un mayor incremento de peso cuando la solución balanceada de sales usada como medio fue adicionada de albumina bovina a una concentración de 5% (peso/volumen). La adicción de proteína a la concentración de 0,5% ó del fagoestimulante GSH no produjo cambios en los aumentos de peso.
Conejos inmunizados a la garrapata, por medio de infestaciones repetidas, desarrollaron reacciones de hipersensibilidad inmediata y tardía a la saliva obtenida mediante la inyección de dopamina a la garrapata. Una reacción similar se encontró en los conejos cuando fueron inoculados con los diferentes medios, después de que las garrapatas se habían alimentado en ellos. El tamaño de la reacción desarrollada demostró un patrón común de reducción a medida que aumenta la dilución del medio, haciendo posible de esta manera estimar la cantidad de alergenos contenidos en cada medio.
De esta manera se demostró, que luego de la alimentación artificial de garrapatas en diferentes medios, el medio que contenía albumina bovina a una concentración de 0,5% contenía alergenos en una cantidad similar a la hallada en la saliva de la garrapata y en mayor cantidad a los alergenos detectados en el medio adicionado de 5% de proteína o en el medio sin aditivos. A pesar de esto, aún fue posible demostrar componentes salivares en el medio simple sin proteina. La importancia de este hallazgo radica en el hecho de que la purificación de estos componentes de la saliva puede ser menos laboriosa en este medio, así estos se encuentren en menor cantidad.
Como la alimentación artificial de garrapatas con tubos capilares es un procedimiento muy laborioso, cualquier intento posterior para alimentar artificialmente gran número de garrapatas tratando de obtener secreciones salivares, podría incluir la inducción de la alimentación artificial en membranas colocadas sobre un medio simple, de manera similar a la reportada por Kemp et al. (1975).