The effect of six different grasses on the excretion rate of the cercopid Aeneolamia reducta was studied in a screenhouse and a growth chamber. The main objective was to measure the excretion rate of adult A. reducta on B. decumbes c.v. Basilisk (susceptible), B. dictyoneura cv. Llanero (tolerant), B. jubata CIAT 16531 (antibiotic), and B. brizantha cv. Marandú (antibiotic). Wax membrane sachets were used for collecting insect excreta. Female A. reducta produced significantly greater quantities of excreta when placed on cv. Basilisk (1.6 ml/insect/day) and cv. Llanero (1.5 ml/insect/day) and cv. Marandú (0.6 ml/insect/day). Gas chromatography of lyophilized samples of excreta of insects showed low sugar content indicating that adults fed from the xylem.
Introduccion
En la Sección de Entomología, del Programa de Pastos Tropicales del CIAT, se han identificado fuentes de resistencia a cercópidos en pastos del género Brachiaria y se pretende elucidar sus mecanismos de resistencia para facilitar el mejoramiento genético. Se han encontrado altos niveles de antibiosis a Aeneolamia reducta (Lallemand) y Zulia colombiana (Lallemand) por medio de bioensayos basados en la sobrevivencia de las ninfas (Ferrufino y Lapointe 1989; Lapointe et al. 1989a; Arango et al.1989). Pathak et al. (1982) utilizaron bolsas de membrana parafinada (sachets) para medir la excreción diaria de las hembras del saltahojas del arroz, Nilaparvata lugens (Stal) (Homoptera: Delphacidae), que se alimentaron sobre variedades susceptibles y resistentes de arroz, y encontraron que el insecto excretó significativamente más en las variedades susceptibles.
Valerio y Nakano (1987) establecieron las tasas de excreción de hembras y machos de Zulia entreriana (Berg) (Homoptera: Cercopidae) en Brachiaria decumbens Stapf cv. Basilisk, y descartaron la posibilidad de que los adultos se alimenten del floema, debido al bajo contenido de azúcar en la excreción. Sin embargo, presentaron una relativamente baja tasa de excreción por gramo de peso del insecto, como evidencia de que los adultos de esa especie no necesariamente se alimentan en forma exclusiva del xilema de la planta.
En el presente trabajo se utilizó el método de Pathak et al. (1982) para estudiar el efecto de accesiones resistentes de Brachiaria sobre el comporta- meiento alimenticio de los adultos de A. reducta. Se analizó su empleo futuro como método de evaluación de germoplasma por su rápidez y bajo costo. Se presentan datos de la tasa de excreción, su gravedad específica, así como también del contenido de azúcares en muestras liofilizadas de excreción.
Materiales Y Metodos
Los experimentos se realizaron en una casa de malla (27 ± 1°C y 70 ± 7% H. R.) o en un cuarto ambiental a 25 ± 1°C y 70 ± 25% H.R. y un fotoperíodo de 12 horas luz, en las instalaciones del Centro Internacional de Agricultura Tropical, CIAT, en Palmira, Colombia. En todos los experimentos se usó el diseño completamente al azar; se hicieron análisis de varianza, pruebas de rango múltiples de Duncan al 5% de probabilidad (Duncan 1955) y regresiones lineales usando el paquete estadístico SAS (SAS Institute Inc. 1985).
Se utilizaron las siguientes gramíneas de acuerdo a su resistencia a ninfas de Z. colombiana y A. reducta (Ferrufino y Lapointe 1989; Lapointe et al. 1989a; CIAT 1989):
Brachiaria jubata CIAT 16531 (antibiótica)
B. brizantha cv. Marandú (antibiótica)
B. dictyoneura cv. Llanero (tolerante)
B. ruziziensis CIAT 654. (susceptible)
B. decumbens cv. Basilisk (susceptible)
Andropogon gayanus cv. Carimagua 1 (resistente a nivel de campo pero susceptible en condiciones de invernadero).
El material vegetativo se sembró en materos de cartón de 6 cm de altura y se trasplantó, un més después, a materos plásticos de 21 cm. Sesenta días después de la siembra, las plantas estaban en condiciones de ser utilizadas en los experimentos.
Los adultos de A. reducta se tomaron de la colonia de la Sección de Entomología del Programa de Pastos Tropicales del CIAT (Lapointe et al. 1989b). La excreción se colectó en bolsas de 7,5 cm de largo por 6,0 cm de ancho, elaboradas de acuerdo a Pathak et al. (1982) y adheridas a los tallos de las plantas (Figura 1). En cada bolsa se confinó un solo adulto durante 24 horas. Después de retirados los insectos a bolsas con su contenido, se pesaron en una balanza analítica. El líquido se extrajo con una jeringa graduada y se anotó el volumen. Si quedaban gotitas de residuo, estas se secaron con papel filtro y se pesó la bolsa vacía. La diferencia entre estos pesos se usó para calcular el peso del líquido excretado y su densidad específica.
Bolsa de menbrana parafinada (sachet) colocado en el tallo de Andropogon gayanus. Nótese la excreción de una hembra de Aeoneolamia reducta durante 24 horas.
Se realizaron tres experimentos. En el primero, se estudio la tasa de excreción de hembras y machos. En el segundo, la tasa de excreción de hembras de varias edades y, en el tercero, se comparó la tasa de excreción en gramíneas con diferentes niveles de resistencia.
Para estudiar la tasa de excreción de machos y hembras, se confinaron individualmente 10 adultos de cada sexo, sin tener en cuenta su edad, en plantas de cv. Marandú, B. ruziziensis CIAT 654 y cv. Carimagua 1, y se colectó la excreción producida diariamente durante cinco días. Los insectos muertos se reemplazaron con otros.
Para determinar la tasa de excreción asociada con la edad del insecto se utilizaron hembras recién emergidas, las cuales se dejaron adaptar y alimentar en cada una de las seis gramíneas durante 1, 2 y 3 días. Antes de confinarlas en las bolsas se matuvieron sin alimento una hora y no se utilizaron más de una vez. Para cada edad y cada gramínea se hicieron 10 repeticiones (hembras).
La tasa de excresión asociada con la resistencia de la planta se midió empleando hembras recién emergidas, las cuales se dejaron adaptar y alimentar durante 3 días en cada gramínea. Para cada gramínea se utilizaron 60 hembras.
A muestras liofilizadas de la excreción de A. reducta, obtenidas en cada gramínea se le hizo un análisis de azúcares totales por cromatografía de gases. El análisis se llevó a caba en los laboratorios del Departamento de Química de la Universidad del Valle y se usó como método indirecto para esclarecer el sitio de alimentación del insecto.
Resultados
1. Tasa de excreción por sexo
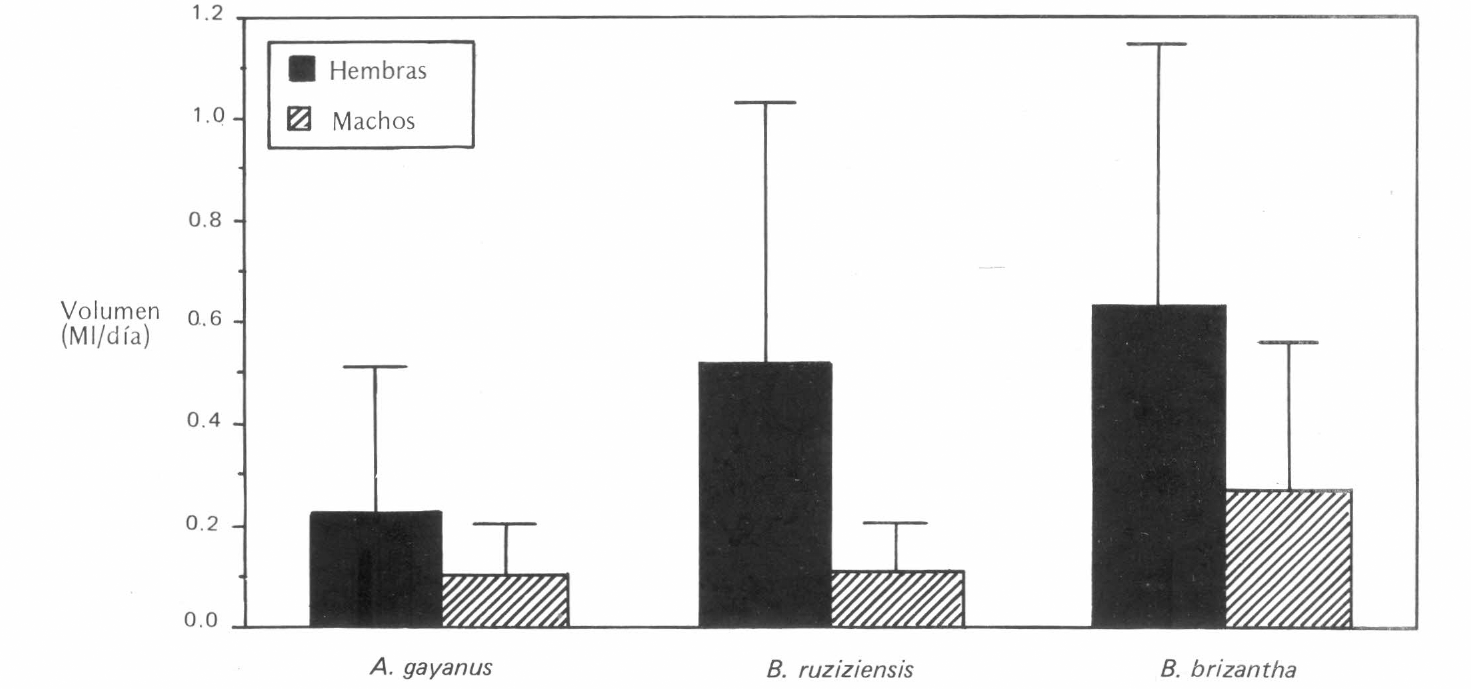
El volumen promedio excretado durante cinco días fue significativamente mayor en las hembras (0,46 ml) que en los machos (0,17 ml) de A. reducta (Figura 2). El análisis de varianza mostró un efecto significativo de la especie de gramínea y del sexo del insecto sobre la tasa de excreción pero no se presentó interacción entre las dos variables. Este resultado coincide con lo obtenido para otros cercópidos como Philanus spumarius (L.) (Horsfield 1978) y para Z entreriana (Valerio y Nakano 1987).
Volumen diario excretado por hembras y machos de Aeneolamia reducta en tres especies de gramíneas. Las barras corresponden a las desviaciones estándar.
Se estableció que para futuros experimentos se debe utilizar un solo sexo para reducir una fuente adicional de variación.
2. Tasa de excreción asociada con la edad.
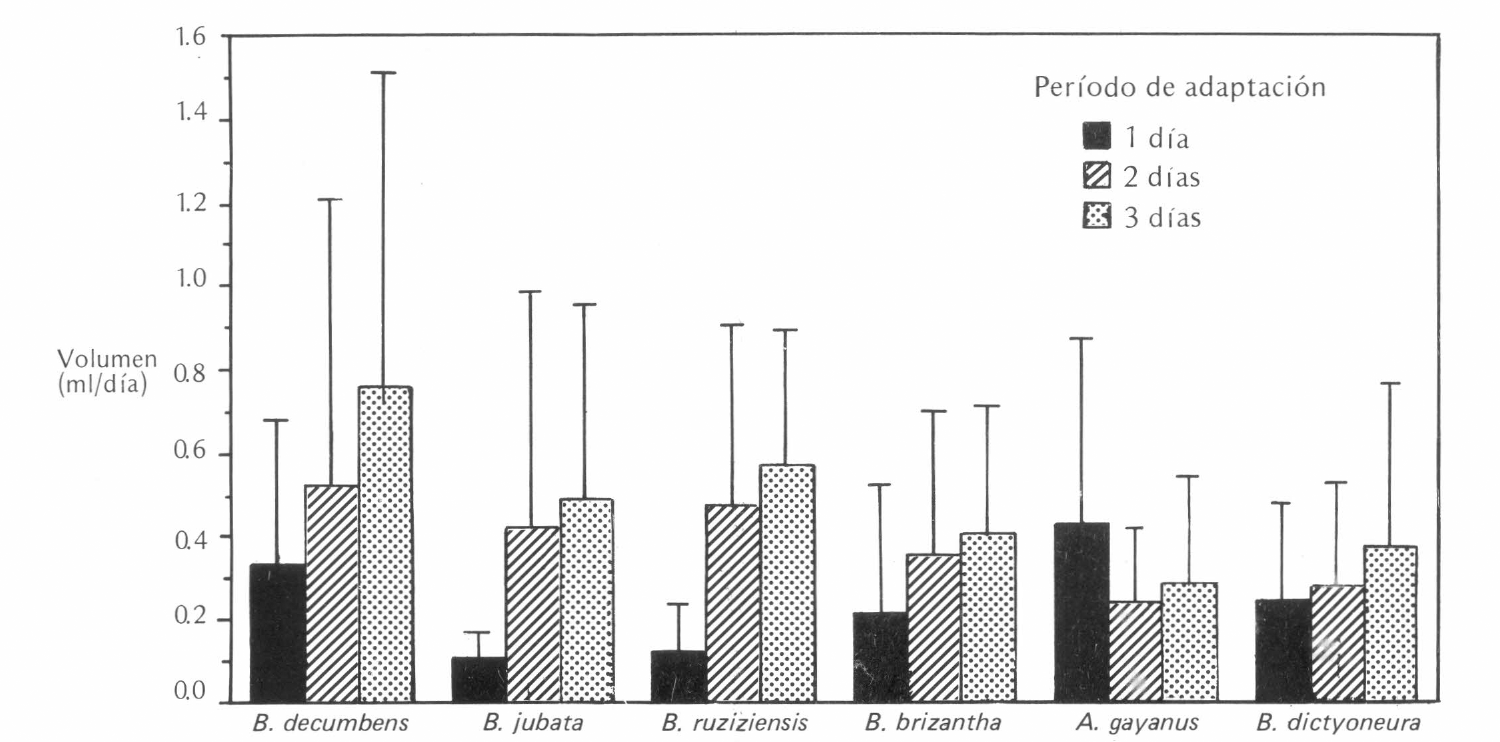
No se detectaron diferencias estadísticamente significativas en las tasas de excreción cuando las hembras se adaptaron durante 1, 2 ó 3 días en cada gramínea (Figura 3). Sin embargo, se escogió la edad de tres días para hembras a utilizar en evaluaciones futuras, debido a la tendencia a producir mayor cantidad de excreción al tercer día.
Volumen diario excretado por las hembras de Aeneolamia reducta después de adaptadas durante uno, dos y tres días en cada gramíneas.
Se encontró una relación lineal directa y significativa entre el peso del líquido y el volumen que excretan las hembras en cada una de las gramíneas (Volumen = 0,03 + 0,9 (Peso); rβ = 0,86). Esto facilitó las evaluaciones, pues. con el peso del líquido se pudo estimar confiablemente el volumen excretado. El peso de la excreción es más fácil de obtener por la diferencia entre los pesos de la bolsa con y sin líquido. En cambio, el volumen es particularmente difícil de medir cuando hay poca cantidad de líquido.
3. Tasa de excreción asociada con la resistencia de la planta.
La tasa de excreción fue significativamente mayor en las hembras alimentadas en los cultivares Basilisk (susceptible) y Llanero (tolerante) que en B. jubata CIAT 16531 (antibiótica) y el cultivar Marandú (antibiótica) (Tabla 1). Para B. ruziziensis CIAT 654 y A. gayanus cv. Carimagua 1 se confirmó con A. reducta, lo encontrado por Ferrufino y Lapointe (1989) con Z. colombiana; estas dos accesiones son buenos huéspedes para el insecto, aunque no tanto como Basilisk y Llanero. Hembras de A. reducta, alimentadas en B. decumbens cv. Basilisk, tuvieron un peso fresco (promedio ± D. E.; N= 35) de 0,02 ± 0,003 g. al inicio del período de alimentación y excretaron 1,66 ± 0,97 g. de líquido, o sea un promedio de 83,1± 51,6 veces su peso.
Volumen del líquido excretado durante 24 horas por hembras de A. reducta de tres días de edad, en seis especies de gramíneas.
Promedios seguidos por la misma letra no presentan diferencias significativas, prueba de rangos múltiples de Duncan al 5% de probabilidad.
Error Estándar
4. Sitio de alimentación
La excreción de A.reducta tuvo una densidad específica (relación peso/volumen) entre 1,05 y 1,000067 g/ml. Esta densidad es muy parecida a la del agua que se considera 1,0, lo cual evidencia que el insecto excreta básicamente agua, y sugiere que el sitio de alimentación es el xilema de la planta (Cheung y Marshall 1973).
La cromatografía de gases mostró concentraciones bajas de azúcares en la excreción de las hembras, en todas las gramíneas (Tabla 2). Resultados similares se encontraron para P. spumarius (Weigert 1964) y Z. entreriana (Valerio y Nakano 1987). También, el porcentaje de agua presente en la excreción (98,1 99,3%) es muy similar el encontrado por Cheung y Marshall (1973) en cicadas, que se alimentan del xilema.
Análisis cuantitativo de azúcares en la excreción de hembras de A reducta alimentadas en cinco especies de gramíneas.
La excreción obtenida por Valerio y Nakano (1987) con hembras de Z. entreriana alimentadas en cv. Basilisk fue aproximadamente ocho veces inferior a la obtenida en este estudio con hembras de A. reducta alimentadas en el mismo cultivar. Aunque se trata de dos especies diferentes, esta diferencia podría deberse a la heterogeneidad de las hembras empleadas. Valerio y Nakano (1987) no mencionan la edad de los adultos ni su procedencia. Así mismo, las diferencias entre individuos hacen necesario el empleo de muchas repeticiones en este tipo de experimentos. En el presente trabajo los coeficientes de variación fueron menores cuando se midió la excreción de un número mayor de hembras de edad controlada. Es probable que el período de adaptación de las hembras al alimento haya contribuido a estabilizar la tasa de excresión diaria.
Las diferencias en la tasa de excreción de los adultos de A. reducta dependieron del nivel de resistencia de la gramínea hospedante, lo cual es consistente con los resultados de Auclair (1958, 1959, 1963); Sogawa y Pathak (1970); Karim (1975); Paguia et al. (1980); Pathak et al. (1982); Cook et al. (1987); Spiller y Llewelyn (1987) y Khan y Saxena (1984, 1988) en los cuales la tasa de excreción de varias especies de insectos fue menor a medida que aumentó la resistencia en sus plantas hospedantes.
Aparentemente, los adultos de A. reducta responden a las plantas antibióticas reduciendo la ingestión de savia, si se compara con las plantas susceptibles. Se puede asumir que el insecto es capaz de percibir diferencias en la composición química de la savia de las gramíneas.
Valerio y Nakano (1987) no pudieron demostrar definitivamente que Z. entreriana se alimenta del xilema de B. decumbes cv. Basilisk, a pesar de que encontraron bajos contenidos de azúcar en la excreción. Encontraron que el insecto excretó alrededor de diez veces su peso fresco en 24 horas. Este factor fue muy bajo comparado con el hallado por Mittler (1967), citado por ellos, de cien a mil veces el peso fresco de Hordnia circellata y el de 180 a 220 veces obtenido por Horsfield (1978) para P. spumarius.
Por los resultados aquí presentados y otros similares obtenidos por Cheung y Marshall (1973) en cicadas, Weigert (1964) en P. spumarius, y Valerio y Nakano (1987) en Z. entreriana, puede concluirse que A. reducta se alimenta del xilema de su planta hospedante.
Se debe resaltar que la concentración de azúcares en la excreción de hembras provenientes de B. jubata CIAT 16531 es entre 7 y 55 veces superior a la en- Contrada en las otras accesiones (Tabla 2). El porcentaje de azúcares en la savia de esta planta debe ser mucho mayor que en las otras gramíneas estudiadas. Posiblemente por esto, los adultos de A. reducta requieren ingerir una menor cantidad de alimento para suplir sus necesidades metabólicas, lo que probablemente se refleja en la cantidad de líquido excretado.
Por los resultados aqui presentados y por los obtenidos por Ferrufino y Lapointe (1989), Lapointe et al. (1989a) y Arango et al. (1989), es evidente que operan mecanismos de antibiosis distintos en B. jubata CIAT 16531 y en B. brizantha cv. Marandú. Todavía no se puede especular sobre el tipo de factores responsables. No obstante, las posibles causas incluyen aleloquímicos como toxinas o fagorepelentes o la ausencia, en la savia, de algún nutriente o fagoestímulante.
Uno de los trabajos futuros de la Sección de Entomología del Programa de Pastos Tropicales del CIAT será la identificación de los compuestos responsables por la antibiosis y el examen de la posibilidad de incorporarlos genéticamente accesiones de Brachiaria adaptadas a suelos ácidos, como los de los Llanos orientales de Colombia.
Conclusiones
Las hembras de A. reducta presentaron una mayor tasa de excreción que los machos.
Se presentó una relación lineal directa (r2 = 0,86) entre el peso y el volumen del líquido excretado.
La tasa de excreción disminuyó con la resistencia de la planta. Los insectos alimentados en especies susceptibles y tolerantes presentaron una mayor tasa de excreción que los alimentados en especies antibióticas.
A. reductase alimenta del xilema de la planta de acuerdo con la gravedad específica de su excreción (cercana a 1) y por las concentraciones muy bajas de azúcares en la excreción.
Footnotes
Agradecimientos
Los autores desean expresar sus agradecimientos a la Dra. Marta Isabel Paéz, del Departamento de Química de la Universidad del Valle, por la elaboración de los análisis químicos y al Dr. J.R. Valerio de EMBRAPA/CNPGG, Campo Grande, Brasil, por sugerencias útiles para la elaboración de este trabajo.
References
1.
ARANGOG.; LAPOINTES.L.; SERRANOM.S.1989. Antibiosis en Brachiaria jubata a los cercópidos Zulia colombiana Lallemand y Aeneolamia reducta Lallemand. En: Congreso de la Sociedad Colombiana de Entomología, 160. Medellín, julio 25-28, 1989. Resúmenes. Medellín, SOCOLEN. p. 36.
2.
AUCLAIRJ. L.1958. Honeydew excretion in the pea aphid Acyrthosiphon pisum (Harr.). Journal of Insect Physiology (Inglaterra) v. 2, p.330-337.
3.
AUCLAIRJ. L.1959. Feeding and excre-tion by the aphid, A. pisum (Harr.), rea-red on different varieties of peas. Entomologia Experimentalis et Applicata (Holanda) v.2, p. 279-286.
4.
AUCLAIRJ. L.1963. Aphid feeding and nutrition. Annual Review of Entomology (Estados Unidos) v.5, p. 439-490.
5.
CHEUNGW.W.K.; MARSHALLA.T.1973. Water and ion regulation in ci-cadas in relation to xylem feeding. Journal of Insect Physiology (Estados Unidos) v. 19, 1801-1816.
6.
CENTRO INTERNACIONAL DE AGRICULTURA TROPICAL. 1989. Informe Anual. Programa de Pastos Tropicales 1989. Cali. CIAT. s.p.
7.
COOKA.G.; WOODHEADS.; MAGALITV.P.; HEINRICHΕ.Α.1987. Variation in feeding behaviour of Nilaparvata lugens on resistant and suscepti-ble rice varieties. Entomologia Experi-mentalis et Applicata (Holanda) v. 43, p. 227-235.
8.
DUNCAND.B.1955. A significance test for differences between ranked treat-ments in an analysis of variance. Virginia Journal of Science (Estados Unidos) v.2, p. 171-189.
9.
FERRUFINOC.A.; LAPOINTES. L.1989. Host plant resistance in Brachiaria grasses to the spittlebug Zulia colombiana. Entomologia Experimentalis et Applicata (Holanda) v.51, p. 155-162.
10.
HORSFIELDD.1978. Evidence for xylem feeding by Philaenus spumarius (L.) (Homoptera: Cercopidae). Entomologia Experimentalis et Applicate (Holanda) v. 24, p. 95-99.
11.
KARIMA.N. M.R.1975. Resistance to the brown planthopper, Nilaparvata lugens (Stal), in rice varieties. Los Baños, University of the Philippines. 131 р. (M.S. Thesis).
12.
KHANZ. R.; SAXENAR. C.1984. A tech-nique for demostrating phloem or xylem feeding by leafhoppers (Homoptera: Ci-cadellidae) and planthoppers (Homop-tera: Delphacidae) in rice plant. Journal of Economic Entomogloy, (Estados Unidos) v. 77, p. 550-552.
13.
KHANZ. R.; SAXENAR. C.1988. Probing behavior of three biotypes of Nilaparvata lugens (Homoptera: Delphacidae) on different resistant and susceptible rice varieties. Journal of Economic. Entomology (Estados Unidos) v. 81 no. 5, p. 1338-1345.
14.
LAPOINTES. L.; ARANGOG.; SOTELOG.1989a. A methodology for evaluation of host plant resistance in Brachiaria to spittlebug species (Homoptera: Cercopi-dae). En: International Grassland Congress, 160. Nice (France), 1989. Proceedings. v. 1, p. 731-732.
15.
LAPOINTES. L.; SOTELOG; ARANGOG.1989b. Improved rearing technique for spittle-bugs (Homoptera: Cercopidae). Journal of Economic Entomology (Estados Unidos) v.82 no. 6, p. 153-162.
16.
PAGUIAP.; PATHAKM.D.; HEINRICHSE. A.1980. Honeydew excretion measu-rement techniques for determining differ-ential activity of biotypes of Nilaparvata lugens on irce varieties. Journal of Eco-nomic Entomology (Estados Unidos). v. 73, p. 35-40.
17.
PATHAKM.D.; SAXENAR.C.; HEINREICHSE.Α.1982. Parafilm sachets for measurign honeydew excretion by Nilaparvata lugens on rice. Journal of Economic Entomology (Estados Unidos) v. 75, p. 194-195.
18.
SAS Institute Incorporated, 1985. SAS user guide: Stadistic. 5 ed. Cary, N.C. 956 p.
19.
SOGAWAK.; PATHAKM.D.1970. Me-chanism of brown planthopper resistance in Mudgo variety of rice (Homoptera: Delphacidae). Appl. Entomol. Zool. v.5, 154-58.
20.
SPILLERN.J.; LLEWELLYNM.1987. Honeydew production and sap ingestion by the cereal aphids Rhopalosiphum padi and Metapolophium dirhodum on seedling of resistant and susceptible wheat species. Annals of Applied Biology (Inglaterra) v. 110, p. 585-590.
21.
VALERIOJ.; NAKANOO.1987. Tasa de excrecao e atividade alimentar do adul-to da cigarrinha-das-pastagens Zulia en-treriana (Berg) (Homoptera: Cercopidae). Anais da Socieda do Entomologia do Brasil, v. 16 no. 2, p. 333-340.
22.
WEIGERTW.G.1964. The ingestion of xylem sap by meadow spittlebug Philae-nus spumarius (L.). American Midland Naturalist (Estados Unidos) v. 71, p. 422-428.