
Abstract
The mechanism of resistance to
Field studies pointed out indicated that tolerance is an important mechanism responsible for resistance in variety G40036. Feeding and ovipositional preferences were detected in the susceptible variety G40128.
Feeding preferences were not reconfirmed under greenhouse conditions. On the other hand, greenhouse studies reconfirmed ovipositional antixenosis in G40036 and ovipositional preferrence for the susceptible variety, G40128.
None of the materials tested had antibiotic efects on eggs, nymphs, and adults of
It is concluded that resistance of
Introduccion
El mejoramiento del fríjol común (
El presente trabajo se fijó como objetivo general determinar el o los mecanismos de resistencia de
Evaluar, por su tolerancia en campo, la resistencia de tres variedades de
Determinar en pruebas de no escogencia y libre escogencia, bajo invernadero, posibles preferencias de alimentación y oviposición de
Evaluar en condiciones de invernadero, los posibles efectos antibióticos de tres variedades de
Revision De Literatura
Schoonhoven (1974) reporta pérdidas, para Colombia, causadas por este insecto, de 14-23% en estaciones lluviosas y del 73-95% en estaciones secas.
La biología y hábitos del insecto fueron estudiados por Wilde et al, 1976.
La naturaleza de las variedades resistentes a insectos está clasificada en tres amplias categorías: tolerancia, no preferencia y antibiosis (Painter, 1951).
Las plantas que presentan mecanismos de defensa que no afectan directamente al insecto se consideran como plantas tolerantes. Las plantas tienen capacidad para recuperarse del daño. Este no es un mecanismo defensivo eficiente, puesto que el costo metabólico sólo se realiza si existe un daño del insecto (Kogan, 1975).
Una planta puede poseer diferentes factores que no la hacen atractiva al insecto para oviposición, alimentación o refugio. Antixenosis fue el término propuesto por Kogan y Ortman (1978), para reemplazar la cateogría de no preferencia de Painter.
Las defensas que afectan la fisiología del insecto después de la ingestión son los factores antibióticos. En la mayoría de los casos estudiados, la antibiosis parece que se debe a diferencia en los constituyentes químicos de la planta (Zúñiga, 1985a).
Con respecto al control genético, el CIAT ha evaluado hasta el momento 18.000 introducciones de
Aunque el fríjol común es el principal hospedante de
El uso potencial de
Materiales Y Metodos
Los trabajos se realizaron en la Estación Experimental del Centro Internacional de Agricultura Tropical, (CIAT), localizado en Palmira (Valle del Cauca), con temperatura promedia de 24°C y humedad relativa 67.5%.
Se evaluaron los tres posibles mecanismos de resistencia de
Para tal efecto, fueron seleccionadas las variedades G40036, G40138 y G40128, clasificadas, en estudios previos, como resistente, intermedia y susceptible, respectivamente.
Tolerancia
Los estudios de tolerancia se realizaron en dos ensayos de campo, durante los semestres 1987B у 1988A, utilizando un diseño de parcelas divididas, con cuatro replicaciones, donde las parcelas principales fueron protección (insecticida sistémico monocrotofos) y no protección y las subparcelas fueron las variedades.
La tolerancia fue medida por el efecto del ataque del insecto en el rendimiento de cada material.
Simultáneamente, se evaluaron otros parámetros a los 25; 35 y 45 días después de la siembra, para determinar posibles preferencias por alimentación y ovoposición a nivel de campo, así:
Cantidad de ninfas por hoja en 20 trifolios tomados al azar por subparcela.
Cantidad de adultos colectados mediante una aspiradora de tipo D-Vac en los dos surcos centrales de cada subparcela.
Daño visual en cada suparcela, en escala de uno a nueve, donde uno es una planta totalmente sana y nueve una planta totalmente afectada (Wilde y Schoonhoven, 1976).
Población total de insectos (ninfas y adultos) en cinco plantas seleccionadas al azar en los cuatro surcos laterales de cada subparcela. Las plantas seleccionadas fueron aisladas de otras, con 24 horas de anticipación. Al momento del muestreo, cada planta se cubrió rápidamente con una bolsa plástica de 90 x 90 cm. y se procedió a cortar el tallo a nivel del suelo. Las bolsas fueron llevadas al laboratorio para efectuar el respectivo conteo.
Cantidad de huevos en 20 peciolos tomados al azar por subparcela a los 45 D.D.S. para el primer ensayo (1987B) y a los 12, 25, 35 y 45 D.D.S. para el segundo ensayo (1988A). Estos peciolos fueron hervidos en agua durante 20 minutos y, luego, colocados en lactofenol (preparado como lo describe Carlson y Hibbs, 1962), durante 15 días, para aclarar los tejidos y, al cabo de los cuales, con ayuda de un estereoscopio, se contó la cantidad de huevos.
Cantidad de tricomas por centímetro cuadrado en diez folíolos centrales tomados al azar en cada subparcela a los 25 D.D.S. en el primer ensayo (1987B).
A la cosecha, se evaluaron los siguientes componentes de rendimiento en 10,80 mβ (parcela efectiva de cada subparcela):
Vainas por planta en 20 plantas tomadas al azar.
Granos por vaina en 100 vainas.
Peso de 100 semillas.
Porcentaje de humedad del grano.
Rendimiento de la parcela efectiva.
Para el segundo ensayo (1988A) sólo se midió el rendimiento de la parcela efectiva (10,80 mβ).
Antixenosis
La preferencia para alimentación y oviposición se determinó en condiciones de invernadero (77.2 ± 9% H.R. y 24.2 ± 5°C), mediante pruebas de libre escogencia (combinaciones binarias y combinación de todas las variedades) y no escogencia, a dos edades de la planta: 10 y 20 días, utilizando un diseño completamente al azar con 4 repeticiones.
Los ensayos se realizaron en jaulas de 2.5 x 0.65 x 0.60 m. y cada jaula dividida en seis compartimientos mediante tabiques verticales removibles. En cada compartimento se colocaron seis plantas infestadas con 15 adultos por planta, durante un período variable de exposición, según el ensayo.
Para evaluar la preferencia por alimentación, se tomaron plantas de 10 días de edad y se hicieron evaluaciones a las 72 horas después de infestación, con recuentos de adultos por variedad a las 8 A.M., 12 М. у 5 P.M. En cada oportunidad, se bajaban, según el ensayo, las divisiones de los compartimentos con el fin de aislar los adultos de cada material. Terminado el conteo, las divisiones se levantaban y la población se disturbaba para permitir su redistribución hasta el próximo conteo.
La preferencia para oviposición se midió a los cinco días después de infestar, mediante recuentos de huevos en las dos hojas cotiledonales y en la primera hoja verdadera de cada planta, usando el mismo método ya descrito para conteos oviposiconales a nivel de campo.
Con plantas de 20 días de edad se siguió el mismo proceso de infestación, pero sólo se evaluó preferencia para oviposición.
Antibiosis
Esta prueba se realizó en invernadero (77.2 ± 9% H.R. y 24.2 ± 5°C).
Para medir el efecto antibiótico de las variedades en la eclosión de huevos, se montaron dos ensayos: En uno se contó la cantidad de huevos y en el otro, la cantidad de ninfas emergidas. Para cada una de las pruebas se utilizaron seis plantas de 12 días de edad, infestadas con 15 adultos por planta y sometidas a cinco días de exposición.
Para determinar el efecto antibiótico de las variedades sobre las ninfas, se establecieron crías sobre cada una de ellas y las plantas de cada material fueron infestadas con 1.500 adultos, durante cinco días de exposición y se dejaron emerger las ninfas. A partir de la emergencia de ellas, diariamente se retiraron 30, para medir su tamaño y duración hasta el momento de la emergencia de los adultos.
El efecto antibiótico de las variedades sobre los adultos fue medido bajo los parámetros de fecundidad y longevidad de 40 parejas de insectos recién emergidos y provenientes de crías de la respectiva variedad. Los adultos fueron sexados y confinados por parejas en jaulas pinza, las cuales están conformadas por dos cilindros de tres centímetros de diámetro y uno de altura, sostenidos por una pinza de cabello y cubiertos con tela de nylon.
Para obtener la curva de oviposición de cada hembra, diariamente, se removió de la hoja el tejido comprendido por la jaula pinza, el cual fue aclarado con lactofenol para contar los huevos colocados por hembra y por día. La jaula con los adultos se trasladó cada día a un nuevo tejido. La longevidad fue medida por el tiempo de duración de los adultos.
Resultados Y Discusion
Los datos de los diferentes ensayos fueron sometidos a análisis de varianza y, en los casos en que las F fueron significativas, a separación de medias, por medio de la prueba de rangos múltiples de Duncan.
Tolerancia
En el primer ensayo de campo (1987B), no se encontraron diferencias significativas entre variedades en cuanto a rendimiento, lo cual sugiere un potencial de rendimiento similar en ausencia de infestación (Tabla 1).
Estadísticamente no se detectaron diferencias respecto a las otras variedades dado el alto coeficiente de variación causado por problemas de tipo agronómico en el ensayo, tales como exceso de humedad y pudrición de vainas, condiciones a las cuales las variedades de
Entre los dos ambientes se encontraron diferencias significativas en rendimiento, siendo mayor en promedio para el ambiente de protección (Tabla 1).
Los niveles de tolerancia se ven reflejados en los porcentajes de pérdida y en los índices de susceptibilidad, calculados por la fórmula:

Donde:
RNP
RP
RNPi: Rendimiento sin protección de la variedad.
RPi: Rendimiento bajo protección de la variedad.
Con respecto a la variedad resistente G40036, la variedad intermedia G40138 fue 3.6 veces más susceptible, mientras que la susceptible G40128 fue 3.2 veces más susceptible que la resistente.
En el segundo ensayo de campo (1988A), se mantuvieron las tendencias encontradas en el primero, aunque, bajo condiciones de protección, se encontraron diferencias significativas entre variedades. La variedad susceptible G40128, de igual forma que en el semestre anterior, presentó el mayor rendimiento bajo estas condiciones (Tabla 2).
Efecto del ataque de E. kraemeri en el rendimiento de P. acutifolius bajo infestación natural (1987B). Promedios de cuatro repeticiones.
Las medias, dentro de cada columna, seguidas de la misma letra no son significativamente diferentes al nivel del 5% (DUNCAN).
Las medias, dentro de la misma línea, seguidas por la misma letra no son significativamente diferentes al nivel del 5% (DUNCAN).
En respuesta al daño causado por el insecto, la variedad G40128 clasificada como susceptible fue la más afectada, presentando un porcentaje de pérdida del 40%. En cuanto al índice de susceptibilidad, se observa una marcada diferencia entre la variedad resistente G40036 y las demás (Tabla 2).
Efecto del ataque de E. kraemeri en el rendimiento de P. acutifolius bajo infestación natural (1988A). Promedios de cuatro repeticiones.
Las medias, dentro de cada columna, seguidas de la misma letra no son significativamente diferentes al nivel del 5% (DUNCAN).
Las medias, dentro de la misma línea, seguidas por la misma letra no son significativamente diferentes al nivel del 5% (DUNCAN).
Los componentes de rendimiento, (vainas llenas, semillas por vaina y peso de 100 semillas) no mostraron diferencias significativas entre variedades.
Los ensayos de tolerancia y las diferencias en rendimiento, expresados en términos de porcentaje de pérdida e índice de susceptibilidad, sugieren que el mecanismo de tolerancia es importante para explicar la resistencia varietal de
Al contar la cantidad de tricomas por centímetro cuadrado, se encontró que la variedad resistente, G40036, fue la que menor densidad presentó, mostrando resultados contradictorios a los reportados por otros autores, donde la variedad resistente presenta una mayor densidad de tricomas (Pillemer y Tingey, 1978; Zúñiga, 1985B). Teniendo en cuenta lo anterior, se concluyó que este mecanismo de defensa no forma parte de la resistencia varietal de
Los recuentos de poblaciones, hechos en los ensayos anteriormente descritos, permitieron encontrar diferencias significativas en poblaciones de insectos entre las variedades.
En el primer ensayo de campo se encontró que, independientemente de la edad de la planta, la variedad resistente G40036 mostró significativamente menor cantidad de adultos. La significancia se mantuvo en el segundo ensayo a partir de los 35 días de edad (Figura 1).
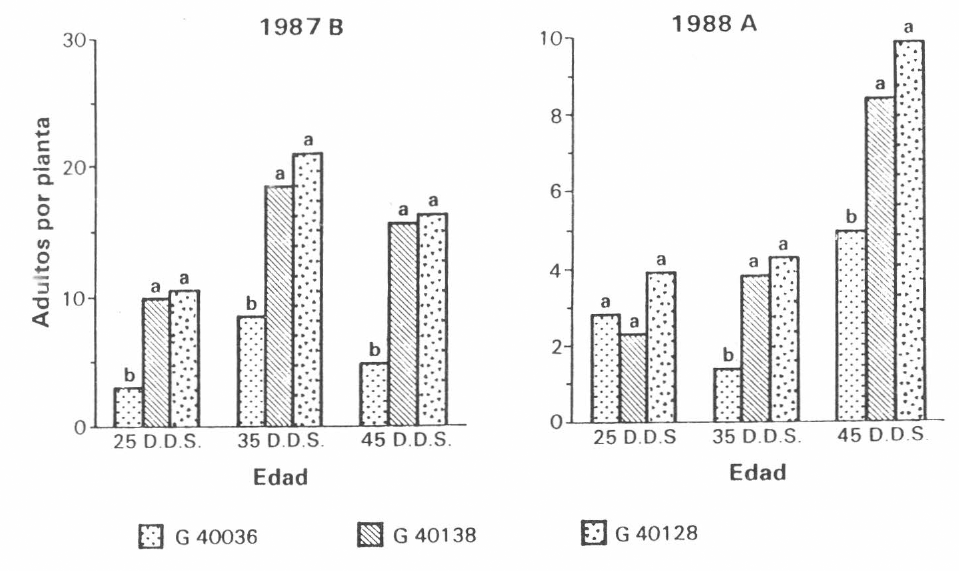
Adultos de
En cuanto a adultos por metro lineal, ninfas por planta y ninfas por hoja, durante los dos ensayos, se mantuvo la tendencia de la variedad resistente G40036 a mostrar menores niveles de población.
Los anteriores resultados sugieren la presencia, en
Las calificaciones visuales de daño reflejaron consistentemente, a través de los dos semestres, la mayor resistencia de G40036, el nivel intermedio de G40138 y la susceptibilidad de G40128. Esta variable, de un alto valor subjetivo, generalmente no refleja diferencias entre variedades.
Se encontraron correlaciones positivas entre los diferentes métodos de contar poblaciones, lo cual sugiere que cualquiera de ellos es confiable para medir y detectar diferencias entre variedades.
Dados los altos niveles de población en el primer ensayo y teniendo en cuenta las marcadas diferencias entre variedades, se determinó hacer un muestreo de oviposición a los 45 D.D.S., el cual mostró preferencia oviposicional hacia la variedad susceptible, G40128 (Figura 2).
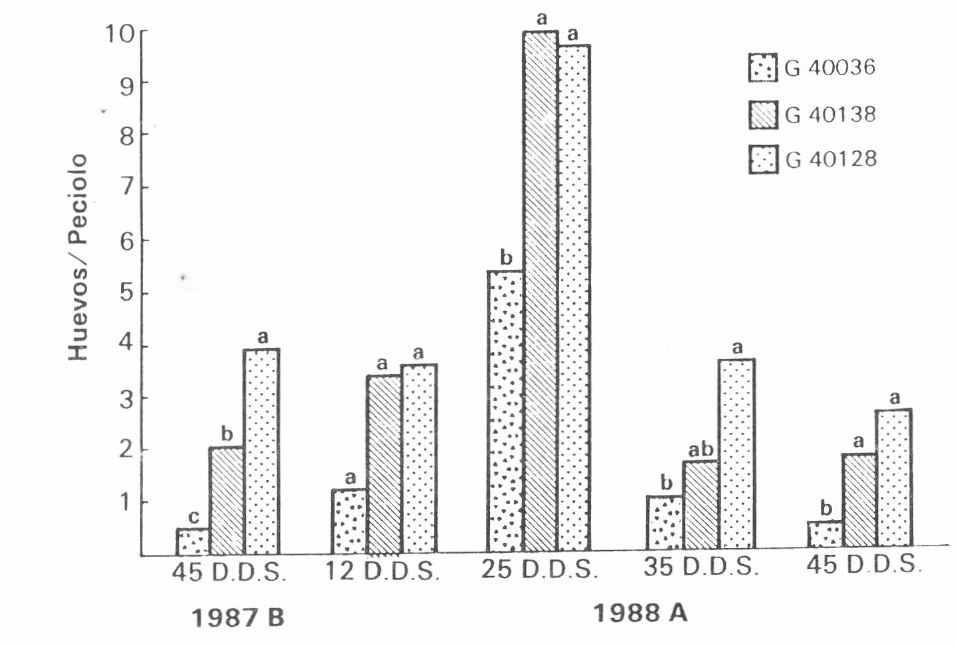
Oviposición de
Al hacer el muestreo de oviposición en el segundo ensayo, no hubo diferencias significativas entre variedades a los 12 D.D.S., pero, a los 25, 35 y 45 D.D.S., las diferencias fueron marcadas, pues la variedad resistente G40036, fue significativamente diferente de la intermedia, G40138 y de la susceptible, G40128 (Figura 2). A través de las diferentes edades, la oviposición fue siempre menor en la variedad resistente.
No se encontró interacción entre variedad y edad, lo cual indica que la preferencia oviposicional se mantuvo a medida que la edad de la planta aumentó. Así se confirmó la presencia del mecanismo de antixenosis por oviposición.
Antixenosis
La preferencia por alimentación encontrada en los ensayos de campo no fue ratificada por las pruebas de invernadero.
En ninguno de los ensayos de invernadero se encontraron diferencias entre variedades en la cantidad de huevos para la edad de 10 días. La variedad resistente G40036 fue la que mostró la menor cantidad de huevos con respecto a las otras variedades (Figura 3).
En los ensayos con plantas de 20 días de edad, se encontraron diferencias significativas entre variedades para la cantidad de huevos puestos en cada planta, siendo la variedad resistente G40036 la menos preferida para ovipositar (Figura 3).
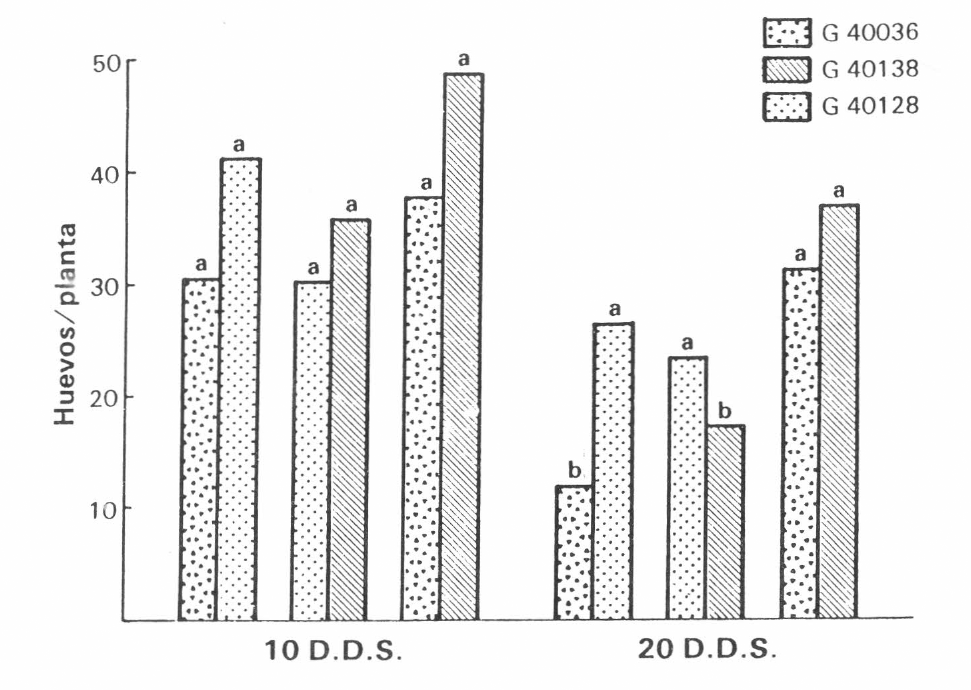
Oviposición de
Las variedades presentaron un comportamiento estable, sin importar, en sus dos edades, si se encontraban solas o asociadas. Así, se confirmaron los niveles de antixenosis para oviposición hallados a nivel de campo (Figura 2).
Antibiosis
El efecto antibiótico de
En este análisis no se encontraron, en las tres variedades, diferencias significativas entre la cantidad de ninfas emergidas y la cantidad de huevos colocados. Al calcular el porcentaje de eclosión, se encontró que, para todas las variedades, fue superior al 90%, descartándose un efecto antibiótico de las variedades sobre la eclosión de huevos (Tabla 3).
Eclosión de huevos de E. Kraemeri en tres variedades de P. acutifolius. Promedios de cuatro repeticiones.
NS: No significativo.
Cuando las tres variedades fueron probadas para detectar su efecto antibiótico en duración sobre las ninfas de
Duración de los instares ninfales de E. kraemeri en tres variedades de P. acutifolius en prueba de antibiosis.
No se encontraron diferencias significativas en la longevidad de hembras y machos (Tabla 5), lo cual sugiere que no hubo efecto antibiótico de las variedades en la longevidad del adulto. La interacción variedad por sexo no fue significativa, lo cual indica que hembras y machos se comportaron de igual manera en las tres variedades.
En la supervivencia de adultos no se encontró diferencia entre variedades, por cuanto las curvas de supervivencia fueron semejantes en forma a través del tiempo, lo cual indica que la mortalidad no es causada por efecto varietal (Figura 4).
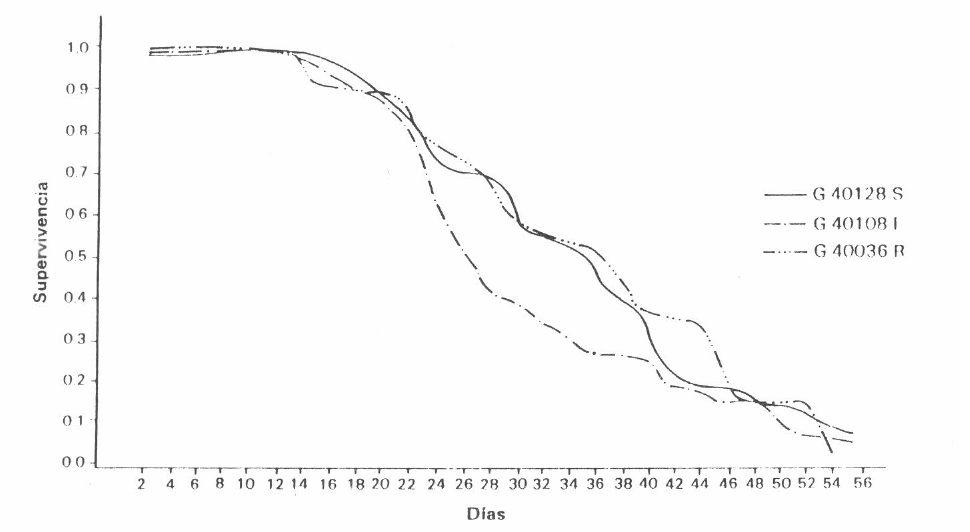
Supervivencia de adultos de
En los ensayos de fecundidad no se encontraron diferencias significativas en cuanto a los días de oviposición, lo cual descarta el efecto de las variedades en el período de oviposición (Tabla 5).
Fecundidad y longevidad de adultos de E. kraemeri en tres variedades de P. acutifolius en pruebas de antibiosis. Promedio de 40 individuos por variedad.
Medias seguidas por la misma letra no son significativamente diferentes al nivel del 5% (Duncan).
La cantidad de huevos por hembra de
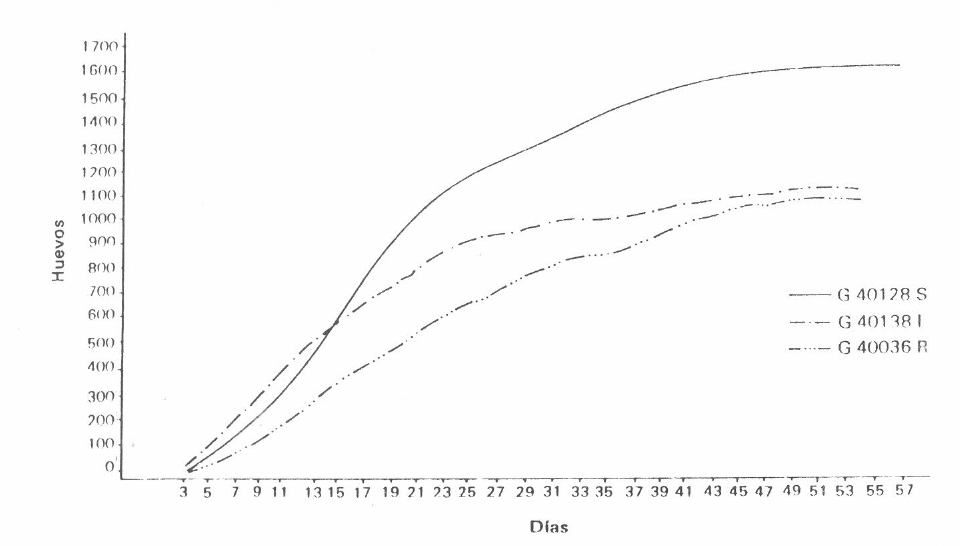
Oviposición acumulada de
Recopilando la información anterior y teniendo en cuenta que no se encontró efecto antibiótico de las variedades en la eclosión de huevos, se reconfirma que la variedad resistente G40036 posee carácter antixenótica para la oviposición y la ausencia de factores antibióticos como mecanismo responsable de la resistencia,
Conclusiones
Bajo las condiciones en que se realizaron los diferentes ensayos de esta investigación, se pueden obtener las siguientes conclusiones:
Que la resistencia de
Que se encontró un nivel de tolerancia en campo, reflejado en un menor índice de susceptibilidad y un menor porcentaje de pérdidas para la variedad resistente G40036.
Que se encontró preferencia por alimentación a nivel de campo, siendo la variedad resistente G40036 la que presentó los menores niveles de población. Dicha preferencia no fue confirmada en ninguna de las pruebas de invernadero.
Que, en el campo, se encontró preferencia por oviposición en la variedad susceptible G40128. Esta preferencia se mantuvo a través de la edad de la planta. Esta preferencia por oviposición, encontrada en el campo, fue ratificada en todos los ensayos de invernadero. La variedad resistente G40036 fue la menos preferida para ovipositar.
Que no se encontró efecto antibiótico de las variedades de