Foliar mildew complex on African Oil Palm (association of Leptinotarsa gibbicarina (Froeschner) Hemiptera: Tingidae with the plant pathogen Pestalotiopsis spp.) is an endemic problem in the Central and Norther regions of Colombia. This abnormality is responsible for reductions up to 60% in yields. The entomopathogenic S. insectorum was found controling up to 98% the lace wing bug L. hevea in rubber plantations of Brazilian Amazonas. This fungy was introduced into Colombia by Indupalma, San Alberto (Cesar), to test against L. gibbicarina; comparing the growth of this fungy in four media (Sabourand, maltosa, agar plus 1% yeast extract SDA; potato, dextrosa, agar PDA; agar plus panvit (mineral salts); semisynthetic salt enrich medium and the comercial medium 5467, specific for entomopathogens), under a completely randomized factorial desing with four replicates in which two strains were sujected to 20, 27 and 30°C. It was found that S. insectorum grew well in all the tested media at 27 and 30°C (65% R.H.), being the optimal temperature 27°С.
Testing pathogenicity under laboratory conditions using five conidial concentrations (10, 105, 106, 107, 108) on 100 adults of L. gibbicarina per treatment, under a completely randomized desing with four replications, it was found a possitive effect at all concentrations in controling the insects, when they were compared to the control (10 ml of water) the porcentages of mortality were: 1.3, 73.5, 73.4 and 72.2% showing not significant differences between differents concentration.
To test pathogenicity of the two strains of fungi under fields conducing a randomized desing was used with the same concentrations and replication as before, but using 100 insects per treatment which were settled up in a palm leaf inside a screeny cage (1,2 m x 40 cm diameter). The differences in porcentage of mortality between the treatments (concentrations) and the control were highly significants for both strains.
For practical proposes, it advisibled to use 107 to 108 conidias/ml of fungy. The results obtained showed the great possibility of using S. insectorum to reach a good control of L. gibbicarina in African Oil Palm. However, more research should be done in other agroecological areas and perhaps in Tingidae of other crops.
Introduccion
La palma africana de aceite (Elaeis guineensis Jacq.) es la especie oleaginosa que mayor cantidad de aceite produce por unidad de superficie. Muchos insectos se han habituado a ella, convirtiéndose en un peligro potencial para la misma y uno de los de mayor cuidado es la chinche de encaje (Leptopharsa gibbicarina. (Froeschner), que, asociada con el complejo fitopatogénico, conocido como Pestalotipsis, que causa el añublo foliar, enfermedad endémica en las zonas central y norte de Colombia.
En Manaos, Amazonas (Brasil), se encontró en caucho, un congénere de la chinche, parasitado en un 98% de su población de campo, por el hongo entomopatógeno Sporothrix insectorum, Hy E. Dicho hongo se importó a Colombia para probar su patogenicidad y adaptación sobre L. gibbicarina, estudiándose su comportamiento, tanto en condiciones de laboratorio como en el campo.
Los resultados de esta investigación ofrecen posibilidades para el manejo integrado de la chinche, problema que, hasta el momento, es prioritario en la palma africana de aceite y es considerado de mucha importancia en Colombia.
Revision De Literatura
La chinche de encaje Leptopharsa gibbicarina F. es el principal diseminador de Pestalotiopsis sp. en palma africana de aceite. Genty y López, en 1973, lo registraron, por primera vez, asociado con el ataque de defoliadores, principalmente limacocidos, causando el 70% de las heridas por las cuales penetran los hongos componentes del añublo foliar, que son considerados patógenos secundarios (Genty, 1975).
L. gibbicarina tiene como hospedante natural en la palma silvestre del Magdalena Medio, llamada Bactris marajá, que no presenta problemas de Pestalotiopsis sp., mientras que, en palma africana, la enfermedad, en tiempo seco, está asociada con la presencia del Tíngidae, pues, en esta época, su población se eleva, especialmente a finales de este período y comienzos del tiempo lluvioso (Genty et al., 1975).
Cuando, por las lluvias, se disminuye la población de L. gibbicarina se activa el ataque de Pestalotiopsis sp., pues los hongos del complejo penetran a través de las heridas dejadas por la chinche sobre las hojas de palma africana en el período seco. Estas heridas son causadas por picaduras y deposición de los huevos. El incremento anormal de la población de L. gibbicarina en la plantación de INDUPALMA S.A., hace que este fenómeno se considere de importancia económica. (García, 1976). En 1986, Celestino Filho y Magalhães encontraron, en las plantaciones de caucho del Centro Nacional de Pesquisa Agropecuaria en Seringueira y Dende (CNPSD) EMBRAPA, Manaos (Brasil), el hongo Sporothrix insectorum parasitando Leptopharsa heveae. Cuando el inóculo es abundante, se observan en las hojas de las plantas de caucho insectos muertos cubiertos por micelio y esporas de color blanco, característico del entomopatógeno (Junqueria et al., 1987).
S. insectorum se aisló de adultos de L. heveae parasitados en el campo y se cultivó masivamente en el laboratorio del EMBRAPA (Brasil) y se importó a Colombia, con el fin de probar su efecto patogénico sobre la chinche de encaje en palma africana de aceite.
El entomopatógeno S. insectorum es un hongo imperfecto que se reproduce asexualmente por esporas o conidias pequeñas, hialinas, insertas en los extremos de conidióforos libres y delgados, originados a partir de hifas septadas (Barnett y Hunter, 1972). El hongo pertenece al grupo de los Hyphomycetos, caracterizados por formar hifas vegetativas de reproducción asexual.
Este hongo, inicialmente, se registró en la literatura como Beauveria sp. y, posteriormente, Hoog y Evans lo reclasificaron como S. insectorum (Junqueria y otros, 1988). Este entomopatógeno ataca Hemípteros tíngidos en Manaos (Brasil) y, en condiciones de laboratorio, se observa parasitando insectos benéficos.1
MORAES, L. Comunicación Personal. EMBRAPA, Programa de Entomología. Manaus, Amazonas (Brasil), 1988.
Por exámenes microscópicos en adultos muertos de L. gibbicarina y L. heveae, parasitados por S. insertorum, se comprueba la fase inicial de salida del micelio del hongo, principalmente de las suturas del pronoto y la zona elitral a nivel de escutelo (Ordóñez, 1988).
Los hongos Hyphomycetos penetran la cutícula del insecto por actividad enzimática y llegan a la hemolinfa, donde se reproducen y causan la muerte del insecto e inician una fase de salida hacia el exterior del cuerpo, para comenzar la producción de conidióforos y la esporulación (Rodríguez, 1984).
Materiales Y Metodos
La investigación se llevó a cabo en la plantación de Industrial Agraria "LA PALMA" (INDUPALMA S.A.), localizada en el Municipio de San Alberto, Departamento del Cesar (Colombia). La zona presenta una temperatura promedia de 28°C, una insolación anual de 2.041 horas sol y una pluviometría de 2.524 mm anuales y una humedad relativa del 75% y una estación seca de tres meses, como máximo y está situada a 150 m.s.n.m. Los suelos son areno-limosos con un pH de 4.5 - 6.5 y el material sembrado es IRHO, procedente de Costa de Marfil.
Se compararon dos aislamientos de S. insectorum; uno reaislado a partir de la primera cepa importada por Genty en 1986 y el otro aislado de L. heveae parasitada por el hongo proveniente de plantaciones de caucho del EMBRAPA (Brasil). En este caso, la muestra se mantuvo en cámara húmeda, luego, se introdujo por dos minutos en alcohol etílico al 25% y por un minuto en agua destilada, antes de proceder al aislamiento del patógeno. Inicialmente, las dos cepas usadas se mantuvieron en medio 5467 (Merck)2, en condiciones de oscuridad continua y a una temperatura de 25°C durante una semana, para estimular el desarrollo de las conidias. Después de este lapso, el hongo se reaisló y se purificó.
Medio Merck para hongos entomopatógenos número 5467.
Posteriormente, las dos cepas de S. insectorum se evaluaron a tres temperaturas constantes (20°C; 27°С у 32°C), en los medios de cultivo Sabouraud-maltosa-agar más extracto de levadura al 1% (smae); Saboraud-dextrosa-agar (sda); papa-dextrosa-agar más panvit3 (este último elemento lo constituyen una mezcla de aminoácidos, minerales y vitaminas); medio semisintético enriquecido con sales (sales) y medio para hongos entomopatógenos 5467 Merck. Previo a la siembra en los diferentes medios, se prepararon colonias de cada cepa de S. insectorum bien esporuladas, a partir de las cuales se sacaron porciones de 2 a 3 mm de diámetro para evaluar crecimiento radial en mm. Para cada medio de cultivo, se realizó un diseño completamente al azar con un arreglo factorial 2 x 3 (dos cepas del hongo y tres temperaturas: 20°C; 27°C у 32°C).
Manual DIFCO, 1977.
El crecimiento se evaluó midiendo el radio de la colonia a las 24 horas y cinco evaluaciones más, realizadas cada dos días hasta el día doce después de la siembra.
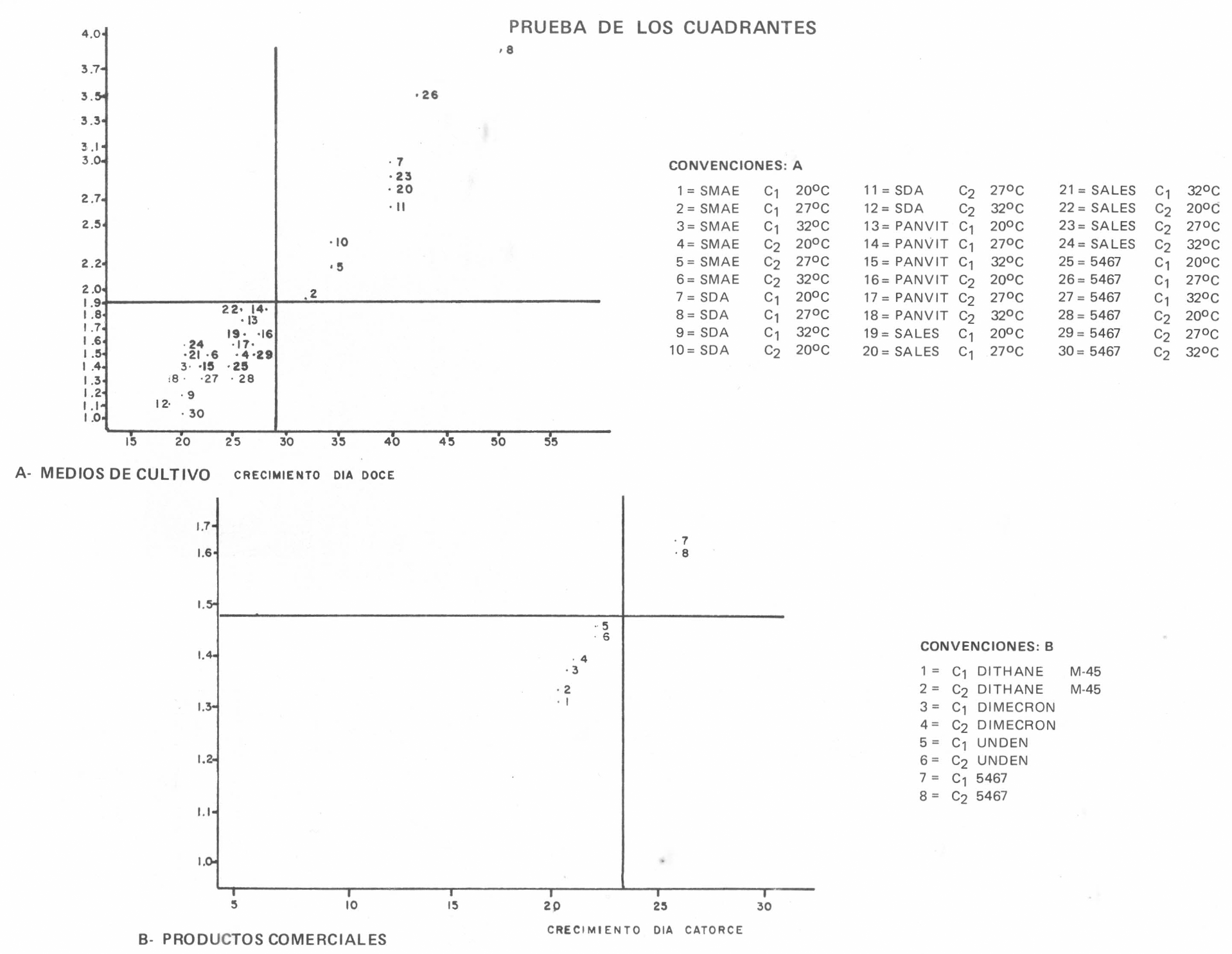
Para seleccionar los mejores tratamientos de acuerdo con el crecimiento, se utilizó como criterio la relación gráfica entre la velocidad de crecimiento por día (y) (tomada como el coeficiente de regresión en los ajustados) y el crecimiento observado a los doce días de. la siembra (x). Al finalizar el ensayo, los tratamientos considerados como los de mayor desarrollo radial con la tasa de crecimiento más alta se compararon usando la prueba "t" para homogeneidad de coeficientes de regresión (tasas de crecimiento diario) y se llevaron a la prueba de los cuadrantes. Aquellos tratamientos urbicados en el cuadrante I, cuyos coeficientes de regresión no eran homogéneos, se consideraban estadísticamente diferentes (Steell y Torrie, 1985).
En el laboratoio, se efectuaron pruebas de patogenicidad, utilizando bandejas esmaltadas con folíolos de palma en los cuales se depositaron veinticinco adultos de L. gibbicarina y las bandejas se asperjaron con 10 ml de la suspensión del hongo a concentraciones de 106, 107, 108, conidias/ml y se usó un testigo tratado, sólo, con agua destilada esterilizada.
El inóculo se obtuvo de cultivos mantenidos, durante siete días a 25°C, en Erlemeyers en 5467 (Merck). A cada Erlemeyer con la colonia esporulada se le adicionaron 100 ml de agua destilada esterilizada con cinco gotas de Tween 20, para facilitar el desprendimiento de las conidias. Para efectuar el conteo de las diluciones, se usó un hemocitómetro tipo Neubauer.
Estas pruebas se desarrollaron utilizando un diseño completamente al azar con cuatro repeticiones. Las evaluaciones se hicieron a las 24, 48, 72 y 96 horas después del tratamiento. Las variables medidas fueron porcentaje de mortalidad y cantidad de insectos afectados por el hongo. Las condiciones ambientales promedias del laboratorio fueron 65% de H.R. y 27°C.
Para los ensayos de patogenicidad en campo, se utilizó un diseño completamente al azar con cinco repeticiones, distribuido en veinte líneas con un área de contenía 463 palmeras. En las palmeras de cada tratamiento, se escogieron las hojas del nivel diecisiete, en las cuales se colocaron jaulas de tull de 1.20 m de largo por 40 cm de diámetro sostenidas por una base de alambre galvanizado número diez y cerradas con cremallera, diseñado por Reyes y Jiménez (1975), para Euprosterna eleasa. Dyar (Lepidoptera: Limacodidae). Las hojas se contaminaron artificialmente con cien adultos de L. gibbicarina por jaula. Previo a los tratamientos se efectuaron conteos de la población y se completaron a cien. Las aspersiones se hicieron usando 30 ml de suspensión de S. insectorum en concentraciones de 105, 106, 107 у 108, conidias/ml y el testigo fue tratado con agua destilada y esterilizada. Los ensayos se efectuaron a comienzos de la época lluviosa, bajo condiciones ambientales de 28°C y 75% H.R. Las evaluaciones se hicieron a las 24, 48, 72, 96 y 120 horas después de la aplicación y las variables medidas fueron las mismas que para las pruebas de patogenicidad en laboratorio. De los insectos muertos por micosis, se reaisló el hongo.
La multiplicación masiva se hizo a partir de cultivos de S. insectorum mantenidos en el medio 5467 Merck en Erlemeyers de 250 ml. Cuando los cultivos se desarrollaban en la superficie del medio y cubrían el diámetro del Erlemeyer, se les añadían, 200 ml de agua destilada y esterilizada, agitando el contenido. Con esta solución micelial, se sembró el medio de multiplicación masiva, según el método usado por Junqueira (1987) en Manaos (Brasil) que se preparó con 1 Kg. de salvado de trigo, 15-20 Kg. de azúcar, 15 ml. de Panvit y10 g. de cloranfenicol. Esta mezcla se secó en un horno a 27°C y, posteriormente, se repartió en frascos de vidrio de boca ancha previamente esterilizados durante 30 minutos a 121°C; luego, en cada frasco, con 1 kg de la mezcla, se depositaron 30 ml de la solución de conidias de S. insectorum. También, se utilizaron otros sustratos, como arroz de tercera y cascarilla de arroz, solos o mezclados con salvado de trigo humedicido en agua azucarada (10 gr/lt), para incrementar la producción de conidias.
La influencia de algunos productos químicos comerciales más usados en palma africana sobre el crecimiento radial de S. insectorum, también se evaluó, al incorporarlos al medio 5467 (Merck), en proporción de 1/10 de sus dosis comerciales (Propoxur: 0.25 gr/hg de I.A.; Mancozeb: 0.002 cc/lt de I.A.; Fosfamidón: 0.7 cc/lt de I.A.) y, posteriormente, sembrar el hongo. El conjunto del material se incubó a 25°C en oscuridad continua durante quince días. El ensayo se realizó bajo un diseño estadístico completamente al azar, con un arreglo factorial 2 x 4 (dos cepas del hongo, cuatro concentraciones, incluyendo el testigo). Los registros del radio de la colonia se llevaron durante los días 3, 8, 12 y 14 después de la siembra. Los modelos de crecimiento del hongo se ajustaron utilizando el método de mínimos cuadrados.
Resultados Y Discusion
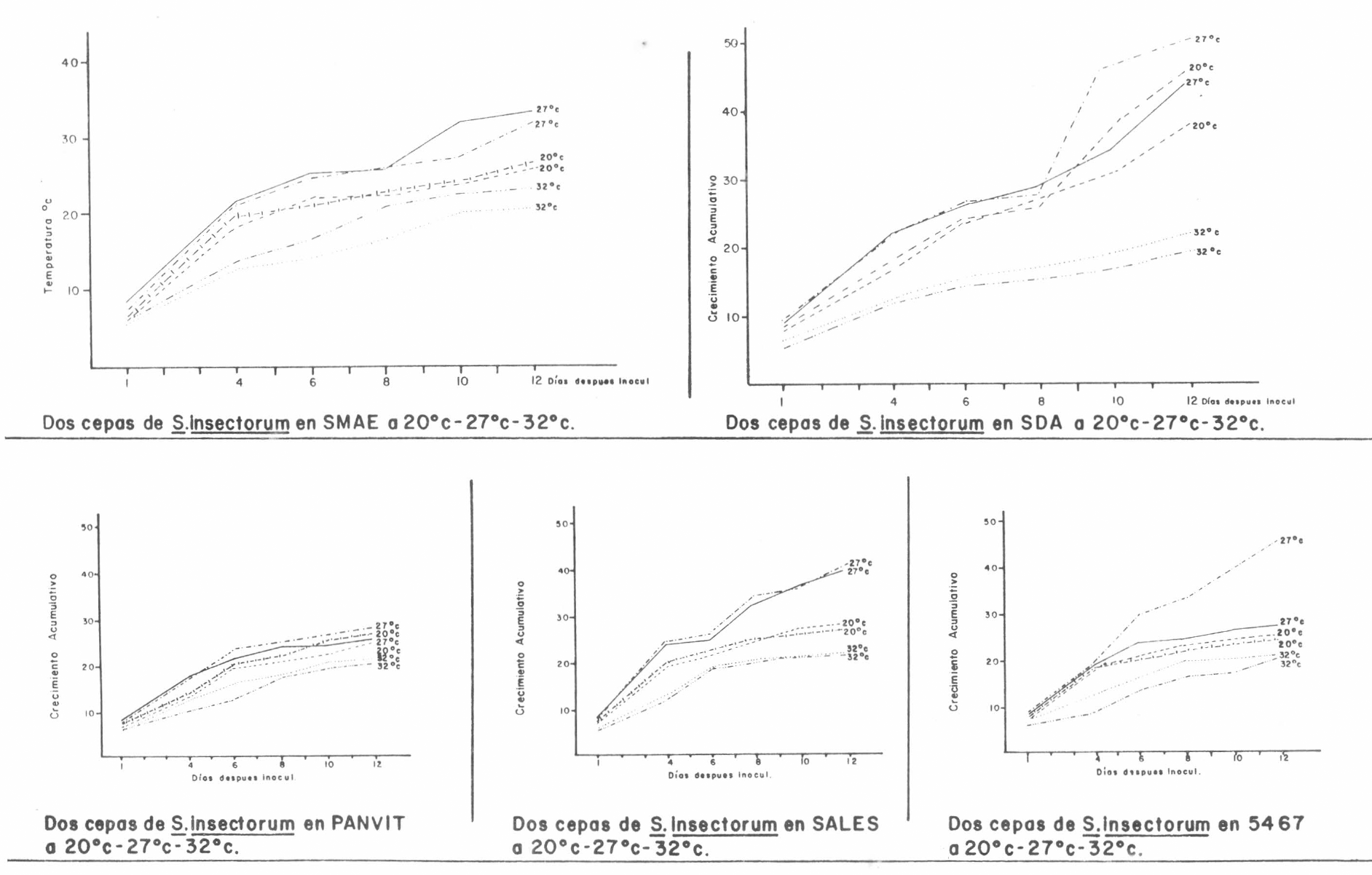
Los resultados del crecimiento radial de las colonias de S. insectorum a tres temperaturas se presentan en la tabla No. 1, tomados a 1, 4, 6, 8, 10 y 12 días después de la siembra, con su respectiva tendencia de crecimiento (Figura No. 1). Posteriormente, se tomó el crecimiento radial al décimo segundo día (x) con la velocidad de crecimiento por día (y), para obtener una relación gráfica que condujo a la prueba "t" (Tabla No. 2) y los coeficientes de regresión se homogenizaron, para establecer modelos de crecimiento a partir de los cuales se hicieron comparaciones biológicas (Tabla No. 3). La velocidad de crecimiento diario (bi), con el crecimiento al décimo segundo día, ubicó los tratamientos en la prueba de los cuadrantes (Figura No. 2A), la cual permitió concluir que el hongo presenta su mejor crecimiento radial a temperaturas entre 27°C y 20°C y que Panvit es el medio de cultivo menos favorable para el desarrollo radial de S. insectorum. El hongo disminuyó su velocidad de crecimiento a temperaturas de 32°C.
Crecimiento radial promedio acumulativo (mm) de dos cepas de S.insectorum a tres temperaturas en cinco medios de cultivo.
Distribución de los tratamientos de acuerdo al crecimiento doce días después de la Siembra y la tasa de crecimiento/día.
Crecimiento acumulativo en mm. de dos cepas de S. insectorum a diferentes temperaturas en cinco medios de cultivo.
Temperatura °C
DIAS DESPUES DE LA INOCULACION
1
4
6
8
10
12
SMAE
CEPA 1
20°C
6.5
19.6
21.4
23.1
24.2
27.0
27°C
7.9
21.3
25.0
26.4
27.8
32.0
32°C
5.3
12.4
14.3
16.9
20.1
21.0
CEPA 2
20°C
6.3
18.6
22.1
23.0
24.0
26.0
27°C
8.3
21.4
25.8
26.3
32.2
34.0
32°C
5.6
13.9
16.9
21.0
22.4
23.0
SDA
CEPA 1
20°C
7.2
16.8
22.0
23.4
34.0
42.0
27°C
8.5
18.9
24.1
25.0
45.0
51.3
32°C
5.1
11.3
13.8
15.9
17.4
20.0
CEPA 2
20°C
6.9
15.9
21.1
24.2
28.0
34.0
27°C
7.9
19.4
23.4
26.0
31.0
40.0
32°C
4.3
10.5
12.9
13.5
15.2
18.0
PANVIT
CEPA 1
20°C
6.7
13.7
19.3
21.3
25.0
26.2
27°C
7.0
16.7
23.2
24.7
26.1
27.8
32°C
5.0
11.3
15.5
17.2
20.3
21.0
CEPA 2
20°C
6.3
12.8
18.7
20.6
22.0
24.3
27°C
7.4
16.8
21.2
23.7
24.2
25.0
32°C
5.8
10.1
12.3
17.0
18.9
20.0
SALES
CEPA 1
20°C
6.9
19.6
22.1
25.0
26.1
27.0
27°C
7.5
24.2
26.0
34.0
35.5
40.1
32°C
5.3
12.9
18.7
20.3
21.0
21.9
CEPA 2
20°C
6.5
18.7
21.3
24.6
27.0
27.8
27°C
7.9
23.3
24.8
32.1
36.4
40.0
32°C
5.0
11.8
18.6
19.4
21.0
21.0
5467
CEPA 1
20°C
7.8
18.6
20.0
22.4
23.9
24.2
27°C
8.9
19.6
29.6
33.6
44.0
45.8
32°C
6.2
12.1
16.3
19.8
20.1
21.3
CEPA 2
20°C
6.9
17.8
21.0
23.0
24.3
25.1
27°C
8.7
19.3
23.8
24.6
26.0
27.8
32°C
5.2
8.3
13.4
16.5
17.1
20.1
Modelos de crecimiento para dos cepas de S. insectorum en cinco medios.
Modelos de crecimiento expresados en significado biológico para S. insectorum.
Comparaciones
V/r. de Tc (+)
Significados biológicos
SMAE C1 a 27°C vs.
0,50 n.s.
Las dos cepas se comportan de manera similar a 27°C en SMAE.
SMAE C1 a 27°C vs. SDA C1 a 27°C
2.93*
La cepa uno reduce su velocidad de crecimiento de manera significativa, cuando se cambia de SMAE a SDA.
SMAE C1 a 27°C vs. SDA C2 a 27°C
1.63 n.s.
A 27°C, las dos cepas se comportan de manera similar en su tasa de crecimiento, ya sea que se siembren en SMAE O SDA.
SMAE C1 a 27°C vs. SALES C1 a 27°C
1.59 n.s.
Las dos cepas de S. insectorum no presentan diferencias significativas en su tasa y velocidad de crecimiento, cuando se ven sometidas a temperatura de 27°C en SMAE y SALES.
SMAE C1 a 27°C vs. 5467 C1 a 27°C
3.53**
La cepa uno, cuando se cambia de SMAE a 5467, incrementa sustancialmente su desarrollo radial.
SMAE C2 a 27°C vs. SDA C1 a 27°C
2.65**
Para una temperatura de 27°C, el crecimiento de las dos cepas de Sporothrix insectorum está asociado directamente con el medio en el cual se siembren. Esto nos indica que el hongo presenta requerimientos con respecto a la nutrición en el medio de cultivo.
SMAE C2 a 27°C vs. SALES C1 a 27°C
SMAE C2 a 27°C vs. 5467 C1 a 27°C
3.19*
SDA C1 a 20°C vs. SDA C1 a 27°C
1.40 n.s.
Las dos cepas presentan un comportamiento similar en su velocidad de crecimiento al ser sembradas en SDA bajo temperaturas de 20°C у 27°C.
SDA C1 a 20° C v.s. SDA C2 a 20°C
SDA C1 a 20°C vs. 5467 C1 a 27°C
4.28*
Un cambio de medio (SDA a 5467) y un aumento de la temperatura (20°C a 27°C) reduce significativamente la velocidad y la tasa de crecimiento de la cepa uno.
SDA C1 a 27°C vs. SDA C2 a 20°C
2.75*
Para un cambio de temperatura (20°C a 27°C), el crecimiento de las cepas uno y dos se hace significativo, sembrándolas en el medio de cultivo SDA.
SDA C1 a 27°C vs. SDA C2 a 27°C
2.07 n.s.
Las cepas uno y dos de Sporothrix insectorum, sembradas en los medios de cultivo SDA, SALES y 5467, se comportan de manera similar en su velocidad y tasa de crecimiento, cuando se las somete a temperaturas de 20°C y 27° С.
SALES C1 a 27°C vs. SALES C2 a 27°C
1.08 n.s.
SDA C1 a 27°C vs. 5467 C1 a 27°C
0.63 n.s.
Expuestas a una temperatura de 27°C, las dos cepas de Sporothris insectorum, sembradas en los medios de cultivo SALES y 5467, muestran velocidad y tasa de crecimiento muy similares. La ausencia de diferencias significativas en este punto de comparación, nos indica que el hongo se desarrolla normalmente a 27°C, si se siembra en medios de cultivo con la nutrición adecuada.
SDA C2 a 20°C vs. SDA C2 a 27°C
0.38 n.s.
SDA C2 a 20°C vs. SALES C2 a 27°C
0.73 n.s.
SALES C1 a 27°C vs. SALES C2 a 27°C
0,020 n.s.
SALES C1 a 27°C vs. 5467 C1 a 27°C
1.55 n.s.
(+) El valor de Tc se comparó con los valores de Tt (4 gl): a niveles de significancia del 5% (2.306) * y del 1% (3.355) **.
n.s. Diferencia no significativa entre las tasas de crecimiento del hongo bajo las tres temperaturas comparadas.
Las pruebas de patogenicidad en laboratorio indicaron que aplicaciones de 106, 107 y 108 conidias/ml de S. insectorum causaron mortalidades entre 71.1% у 73.5% respectivamente, en adultos de L. gibbicarina (Tabla No. 4). La respuesta favorable de S. insectorum sobre L. gibbicarina, en concentraciones de 106, 107 y 108 conidias/ml de solución micelial, se debe a que los factores aleatorios fueron totalmente controlados en el laboratorio y en las bandejas y, por ello, no se observan diferencias significativas entre las mortalidades a estas concentraciones.
Mortalidad de L. gibbicarina en folíolos de palma africana con dos cepas de S. insectorum en laboratorio.
Los tratamientos de la misma letra no son significativamente diferentes (P: 0.05) de acuerdo con la nueva prueba de intervalos múltiples de DUNCAN.
Las pruebas de patogenicidad en campo indicaron que aplicaciones de 108 y 107 conidias/ml de S. insectorum causaron mortalidades hasta del 73% en adultos de L. gibbicarina (Tabla No. 5), observándose diferencias significativas con respecto al testigo. Aplicaciones entre 106 y 105 conidias/ml de S. insectorum causan mortalidades casi cercanas al 50%.
Mortalidad de L. gibbicarina en folíolos de palma africana con dos cepas de S. insectorum bajo campo.
Los tratamientos seguidos de la misma letra no son significativamente diferentes (P: 0.05) de acuerdo con la nueva prueba de intervalos múltiples de DUNCAN.
A pesar de que el hongo no se evaluó sobre estados ninfales de L. gibbicarina, las observaciones de campo indican que, también, afecta este estado y que, usando concentraciones más altas del hongo, se podrían lograr porcentajes de mortalidad superiores al 73% (Figura No. 3).
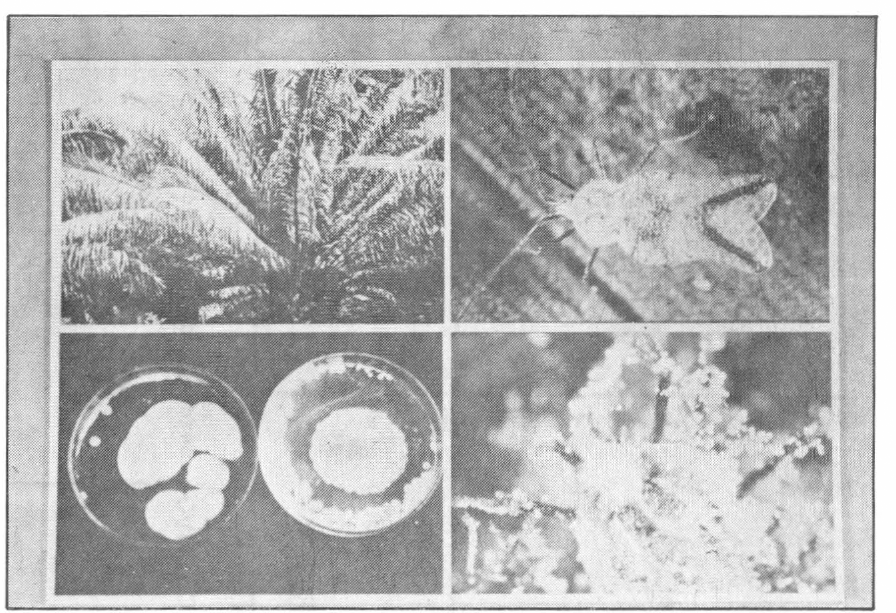
Secuencia del Complejo Pestalotiopsis sp.-Leptopharsa gibbicarina y parasitismo por Sporothrix insectorum. Sup. Izqda. Elaeis guineensis, j. atacado por Pestalotiopsis sp. Foto: Philippe Genty. Sup. Derch. Adulto de Leptopharsa gibbicarina vector indirecto del Añublo Foliar. Foto: Philippe Genty. Inf. Izqda. Hongo entomopatógeno Sporothrix insectorum. Foto: Manolin Avila. Inf. Derch. Adulto de Leptopharsa gibbicarina parasitado por Sporothrix insectorum Foto: Philippe Genty. Collage Foto: Luis Fdo. García A.
De todos los sustratos evaluados para la multiplicación masiva, el salvado de trigo fue el que presentó menor costo y el que propició mayor producción de esporas. S. insectorum tardó doce días en invadir totalmente el salvado de trigo a temperaturas de 24°С у 27°С. Durante este período, el hongo creció en todo el sustrato, formando una masa micelial blanca. Para la obtención de esporas, en aplicaciones de campo, esta masa micelial fue retirada de los frascos y colocada en agua en proporción de 1 kg de sustrato, cubierto por la masa fungosa, para quince litros de agua. Esta mezcla se colocó, posteriormente, en un cedazo. La concentración final de la misma quedó por encima de 107 (diez millones) conidias/ml. cuando se detectó la presencia de la chinche, en días de escasa lluvia se hicieron aplicaciones de la mezcla, evitando utilizar aspersoras usadas en algún producto químico comercial en ocasión anterior.
Una práctica favorable fue eliminar las plantas de Bactris maraja de las zonas cercanas a las parcelas tratadas, ya que es considerada huésped de la chinche.
El efecto de S. insectorum sobre la chinche de encaje, se verificó hasta la segunda generación de los tíngidos (30 - 50 días después de la aspersión), pues se observó la presencia de ninfas y adultos muertos fijados en el envés de las hojas.
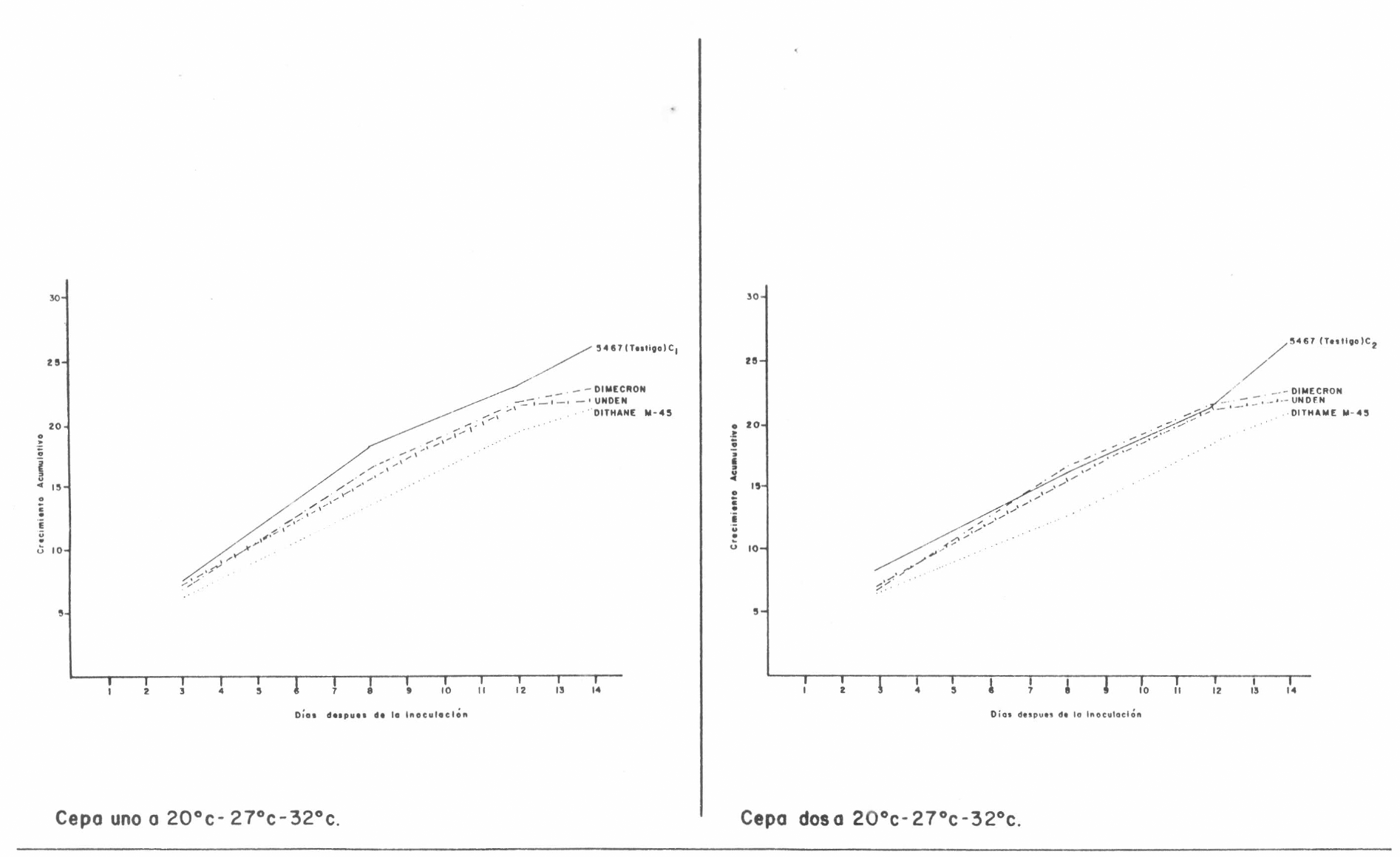
Los resultados de la influencia de los productos comerciales sobre el crecimiento radial de las colonias de S. insectorum a tres temperaturas se presentan en la Tabla No. 6, tomados a los 3, 8, 12 y 14 días después de la siembra, con su respectiva tendencia de crecimiento (Figura No. 4). Finalmente, se tomó el crecimiento radial al décimo cuarto día (x) con la velocidad de crecimiento por día (y); sin embargo, los coeficientes de regresión indicaron que las comparaciones biológicas no eran significativas. Aplicando sobre los tratamientos la prueba de los cuadrantes, se observó que, únicamente, los testigos se encontraron ubicados en el cuadrante I (Figura No. 2B), lo cual permitió concluir que, en condiciones de laboratorio, MANCOZEB, FOSFAMIDON y PROPOXOR inhiben el crecimiento radial de S. insectorum en condiciones de laboratorio.
Crecimiento acumulativo en m.m. de S. insectorum sembrado en 5467 con productos químicos a 1/10 de sus dosis comerciales.
Crecimiento acumulativo en mm. de S. insectorum con productos químicos a 1/10.
Producto Comercial
Días después de la inoculación
3
8
12
14
CEPA 1
DITHANE M-45
6.7
13.8
19.40
21.30
DIMECRON
7.3
16.3
22.00
23.00
UNDEN
7.1
15.9
21.80
22.00
5467 TESTIGO
7.9
18.4
23.60
26.40
CEPA 2
DITHANE M-45
6.3
12.9
18.6
21.00
DIMECRON
6.9
16.8
21.9
22.80
UNDEN
7.0
15.8
21.6
22.10
5467 TESTIGO
8.3
16.4
21.76
26.60
Debido a que este ensayo no se evaluó en el campo, se carece de punto de comparación que permita concluir si el comportamiento observado en laboratorio se mantiene en condiciones ambientales.
Conclusiones Y Recomendaciones
Las dos cepas de S. insectorum presentan buen crecimiento radial (entre 24.2 mm y 51.3 mm al décimo segundo día), a 27°C у 20°C en Sabouraud-dextrosa-agar, Saboraud-maltosa-agar más 1% de extracto de lavadura; medio enriquecido con Sales y Medio para entomopatógenos 5467 Merck, exceptuando para Papa-dextrosaagar más Panvit (entre 24.3 y 27.8 mm al décimo segundo día). En todos los medios, el hongo disminuye significativamente su tasa de crecimiento a temperaturas superiores a 27°C.
La velocidad y tasa de crecimiento del hongo, para las dos cepas, está en relación directa con los nutrientes del medio.
La cepa de S. insectorum proveniente de L. heveae mostró mejor desarrollo y acción patogénica que la cepa existente en la colección de INDUPALMA S.A. (9 meses), aislada del mismo insecto en Brasil, por lo cual se asume que, en el medio, perdió virulencia.
El desarrollo uniforme del hongo en un medio de cultivo permite disponer del material del inóculo y programar adecuadamente los tratamientos en el campo.
La presencia de algunos productos comerciales (MACOZEB, PHOSPHAMIDON, UNDEN) inhibe la velocidad y tasa de crecimiento del hongo en relación con el testigo.
El mejor medio de multiplicación masiva es el salvado de trigo, el cual es el más económico y más fácil de preparar.
Bajo condiciones de laboratorio, a las 96 horas del tratamiento, S. insectorum en L. gibbicarina causa mortalidades por micosis del 73,5 у 73,4 у 72,2% a concentraciones de 108, 107 y 106 conidias/ml del hongo, respectivamente, en comparación con el testigo que es de 1.3%. En pruebas de campo, la mortalidad por micosis es de 72,9; 71,3; 49,1 у 45,3% con dosis de 108 y 107, 106, 105, esporas/ml a las 120 horas, respectivamente, en el testigo, se obtuvo 25%.
En condiciones de campo y de laboratorio, el hongo afecta tanto los estados adultos como los ninfales de L. gibbicarina.
Teniendo en cuenta el porcentaje de mortalidad obtenido en el campo con dosis de 108 y 107, se considera que S. insectorum es un agente microbial con gran potencial de uso para la palma africana de aceite en Colombia, pues estos resultados constituyen el primer registro y justifican incrementar los estudios en este campo.
A dosis de 106 se obtiene un 49.1% de control, lo cual se considera de utilidad, si se tiene en cuenta que en el control integrado de la chinche debe intervenir otros auxiliares.
Footnotes
Agradecimientos
Los autores expresan sus más sinceros agradecimientos al Dr. Nilton Junqueria de EMBRAPA (Brazil), por facilitar la cepa y la información sobre S. Insectorum.
A las doctoras Alba Rodríguez, del ICA (Tibaitatá) y Marina Sánchez de Praguer, de la Universidad Nacional, Facultad de Agronomía (Palmira), por su trabajo de codirección, colaboración y constante apoyo.
Al Dr. Germán Valenzuela, de BAYER S.A. (Colombia) por sus valiosas sugerencias en la realización de este estudio.
Al Dr. Edgar Restrepo y todo el personal de Palmas Oleaginosas BUCARELIA S.A. (Colombia.
Al Dr. Hernando Pabón y todo el personal de PALMERAS DE LA COSTA S.A. (Colombia), por su colaboración en la transcripción de los resultados.
A mi amigo Hernando Patiño Cruz (q.e.p.d.) por haberme introducido al maravilloso mundo de los Artrópodos.
A SOCOLEN por su constante interés en los resultados de la investigación.
Al Sr. Luis Fernando García, por su trabajo de fotografía y a la señorita Esperanza Sánchez, por su colaboración en el trabajo de mecanografía.
References
1.
BARNETTH.L.; HUNTERB.B.1972. Illustred genera of imperfect fungi. Burges, Public. Co., Minneapolis, 214p.
2.
CELESTINO FILHOP.; MAGALHAESF.E.L.1986. Ocurrencia de fungo Sporothrix insectorum (HOOG y EVANS) parasitando a mosca-de renda (Leptopharsa heveae, DRAKE y POOR) en seringal de cultivo. EMBRAPA - CNPSD., Pesquisa en andamento, 42. Manaues, A.M., Brasil.
3.
DIFCO1977. Difco manual of dehydrated culture media and reagents for microbiological and clinical laboratory procedures. 9a. ed., Difco Laboratories Inc., Detroit, Michigan, 350p.
4.
GARCIAR.1976. Nueva plaga de la palma africana Elaeis guineensis Jacq. en Colombia, un Hemíptero Tingidae Gargaphia sp. y asociación con Pestalotiopsis sp. y otros hongos foliares. 1–34p.
5.
GENTYP.1975. Problemas entomológicos de Elaeis guineensis Jacq. en América Latina. 12p. (Mimeografiado).
6.
GENTYP. et al. 1975. Daños y control del complejo Leptopharsa-Pestalotiopsis en palma africana. Oléagineux, Francia, Vol. 30No. 3, 199–204p.
7.
GENTYP. et al. 1975. Daños de Pestalotiopsis consecutivos a unos ataques de Gargahia sp. en Colombia. Oleagineux, FranciaVol. 30No. 5, 119–204p.
8.
JUNQUERIAN. et al. 1987. Isolamento e cultivo do fungo Sporotothrix insectorum (HOOG y EVANS) a ser utilizado para o controle da mosca-de-renda da seringueira. Comunicado Técnico, EMBRAPA- CNPSD. Número 56. Manaus, A.M., Brasil, 1–4p.
9.
JUNQUERIAN., et al. 1988. Controle biologico da mosca-de-renda (Leptopharsa heveae) en seringal de cultivo no estado do Amazonas. En: Memories Primer Simposio Nacional de Controle Biologico de Pragas e Vetores. SICONBIOL, Rio de Janeiro., R.J. Noviembre 1988, 159 p.
10.
ORDOÑEZA.I.1988. Evaluación de la patogenicidad del hongo Sporothrix insectorum (HOOG y EVANS) sobre la chinche de encaje Leptopharsa gibbicarina (FROESCHNER) en palma africana. Tesis de Grado, Universidad Nacional de Colombia. Facultad de Ciencias Agropecuarias. Palmira, Septiembre 1988. 85p.
11.
RODRIGUEZD.A.1984. Hongos entomopatógenos. En: Seminario sobre Patología de insectos. Sociedad Colombiana de Entomología. SOCOLEN. Seccional Medellín. Myo 1984. 157p.
12.
STEELR.G.D.; TORRESJ.A.1985. Bioestadística. Principios y procedimientos. Segunda Edición. Mc. Graw-Hill, Inc.250–251p.