
Abstract
The parasitoids E. diversicornis and A. coccois have been found abundantly in Meta (Carimagua) and Valle del Cauca (CIAT-Palmira), respectively, attacking P. herreni and P. gossypii in cassava cultivar.
In order to compare the behaviour of the two hymenoptera, the following situations were determined: developmental stage of P. herreni preferred by the aforementioned parasitoids, encapsulation by the mealybug, mortality of P. herreni due to parasitism under laboratory conditions (25°C and 70% RH), effect of temperature (20°, 25° and 30°C) on the development of both natural enemies to estimate the physiological time or accumulative temperature, given in days-degree (OD), necessary to complete the development and minimum threshold of development.
The result showed that the 3rd. nynphal instar and the adult female of the mealybug were the stages preferred by the parasitoids. Encapsulation by P. herreni was verified in E. diversicornis but not in A. coccois. P. gossypii did not produce encapsulation in either species of parasitoids. E. diversicornis caused mortality in the first instar of P. herreni, but A. coccois did not cause this kind of effect on the mealybug.
Average period from oviposition to adult emergence was 35.7, 20.2 and 18.1 days for E. diversicornis and 39.5, 18.5 and 19.7 days for A. coccois at 20°, 25° and 30°C, respectively. A minimum threshold of 230°D for E. diversicornis and 175°D for A. coccois was found.
Introduccion
Uno de los insectos plagas que ocasionan daño económico en la planta de yuca es el piojo harinoso,
Ante dicho problema, una de las alternativas, para el manejo de estas plagas, es el uso de enemigos naturales, en programas de control biológico dirigido. Con este fin a partir de 1983B, se inició el reconocimiento de los principales reguladores naturales de la plaga, encontrándose como más importantes los parasitoides
La presente investigación tuvo como objetivo, realizar una comparación de los dos parasitoides de los piojos harinosos, en cuanto a: estado de
Revision De Literatura
Una buena cantidad de enemigos naturales del piojo harinoso son Hymenopteros de la familia Encyrtidae. Observaciones realizadas en la Estación Experimental de Carimagua (Meta), con temperatura promedia anual de 28°C y una humedad relativa de 80%, indicaron la existencia de importantes poblaciones del parasitoide E. diversicornis (Bellotti et al., citado por Reyes J.A. 1983) y en el Centro Internacional de Agricultura Tropical, CIAT-Palmira, se han observado poblaciones de A. coccois (CIAT, 1983).
En Africa, se registran varios parasitoides de piojo harinoso de la yuca (
La familia Encyrtidae presenta un importante cantidad de benéficos, que son, principalmente, parasitoides de plagas del orden Homoptera. Se pueden citar los siguientes:
Algunos huéspedes presentan una reacción de defensa hacia el parasitoides que es la “encapsulación” que se define como una defensa del huésped, al encapsular o impedir el desarrollo de estados inmaduros o posturas de los benéficos (Van Driesche, información pers.). Esta característica del huésped se ha registrado en 14 órdenes de insectos, especialmente en la familia Tachinidae (Diptera) y en Hymenoptera de las familias Ichneumonidae, Braconidae, Encyrtidae, Eulophidae, Pteromalidae y Eucolidae (Askew, 1971).
La forma de la producción de cápsula (encapsulación) por la especie huésped contra la parasita es generalmente invariable. En la mayoría de los casos, diferentes especies de parasitoide generan una misma respuesta en el mismo huésped. Por el contrario, la misma especie parasita en diferentes huéspedes produce una reacción totalmente diferente. (Askew, 1971).
Se ha observado encapsulación en
La encapsulación de huevos de los parasitoides de la Encytida (
Estudios realizados en el parasitoide
La noción de que el desarrollo y crecimiento de muchos organismos depende de la temperatura, especialmente los organismos Poikilotérmicos, data desde la mitad del siglo XVIII y, aún, es un concepto útil en la agricultura moderna (Zalom et al 1983).
Un organismo requiere un tiempo largo para su incremento y desarrollo a bajas temperaturas. Así, al incrementarse este factor abiótico, dicho tiempo decrece progresivamente hasta llegar a ser demasiado alto y, entonces, lo afecta negativamente. Según Zalom et al (1983), la tasa de desarrollo es simplemente el porcentaje del mismo por un día a una temperatura específica.
La relación entre la temperatura y el tiempo de desarrollo (días promedio) da como resultado una curva de la forma de “j” invertida. Si la recíproca de tiempo de desarrollo se toma como la tasa de desarrollo (1/tiempo de desarrollo) versus temperatura, resulta una curva sigmoidea en forma de “s” (Wagner, et al 1984).
Teniendo en cuenta las gráficas de la relación de tasa de desarrollo y temperatura, podemos realizar el cálculo de una constante para un determinado insecto, que es umbral mínimo de desarrollo, que es la temperatura (°C) por debajo de la cual el desarrollo se detiene, sin causarle la muerte a dicho insecto. En la práctica, este umbral mínimo es obtenido por la proyección del segmento lineal de la curva hasta la intercesión del eje de las temperaturas (Zalom, et al 1984).
Una de las constantes del insecto que se calcula es el tiempo fisiológico, que se define en grados-día (OD). Este estudio, que es usado ampliamente, en los trabajos de diversos autores (Caudulle 1955, Reibich 1902, Sanderson y Pearg 1913, Arnold 1960, Baskerville y Ewin 1969, Abrami 1972, Savacheriam et al 1977) es fácil de calcular y de aplicar y, con frecuencia, produce valores aproximadamente correctos (Eckenrode y Chapman 1972, Aliniazee 1976, Bett y McEwn 1981, Obrycki y Tauber 1981). Sin embargo, sólo, es válido para temperaturas medias (Wagner L.T. et al 1984).
Los modelos de población y desarrollo que incorporan umbrales y tasa de desarrollo basados en grados-día, en un evento preciso, pueden ayudar al agricultor y asesorarlo en decisiones sobre el control de plagas. En estos modelos, el ciclo de vida es medido en tiempo fisiológico (grados-día) en lugar de tiempo calendario (días). Usando los grados-día para predecir el desarrollo de un organismo, hace posible minimizar conflictos entre cultivos y operaciones, tales como, irrigaciones y aplicaciones de plaguicidas (Zalom et al 1983).
Como los organismos poikilotérmicos, usualmente, viven en la naturaleza a temperaturas fluctuantes, resulta complicado realizar experimentos con dichas temperaturas y, además, los resultados de los experimentos son difíciles de interpretar (Andrewartha, 1973).
Andrewartha y Birch (1954) realizaron una revisión de literatura existente sobre temperaturas fluctuantes y llegaron a la conclusión de que los principios relacionados con temperaturas constantes eran aplicables, en sentido general, a temperaturas fluctuantes (Andrewartha, 1973).
Materiales Y Metodos
El presente trabajo se realizó en el Centro Internacional de Agricultura Tropical CIAT, ubicado en el Municipio de Palmira a 1060 msnm y una temperatura promedio anual de 24°C. Los experimentos fueron realizados bajo condiciones de laboratorio, en cámaras de temperatura y humedad relativa controladas.
Estados de P. herreni preferido por cada parasitoide.
Se tuvieron en cuenta para este experimento los siguientes estados de desarrollo del piojo: Primer instar ninfal (no presenta dimorfismo sexual), segundo instar hembra, segundo instar macho, tercer instar hembra, cocon o capullo ninfal (tercer y cuarto instar macho) y el adulto hembra.
Es una hoja de yuca, preferiblemente de cinco lóbulos, se colocaron, distribuidos al azar, dos individuos de cada estado del piojo en cada lóbulo. Estos individuos eran expuestos, durante una hora continua, a un individuo hembra del parasitoide para evaluar. Durante este tiempo, se hicieron observaciones: cada ninfa atacada u ovipositada se removió y se cambió por otra del mismo estado. Cuando se observó la penetración de su ovipositor en el cuerpo del piojo, se registró la ovoposición.
Como repetición, se tomó cada individuo parasitoide y se observó por igual tiempo.
Respuesta a la encapsulación.
Se evaluaron dos estados de desarrollo de los huéspedes del piojo harinoso: Segundo y tercer instar y dos temperaturas constantes, 25°C y 30°C.
Plantas infestadas con piojos, en el estado escogido, eran cubiertas con bolsas de tela (organza o tul) y, después de cubiertas, se les introdujo 50 individuos del parasitoide hembra que se iba a evaluar, dejándolos por espacio de tres días y, finalizado este lapso, se extrajeron los parasitoides y la planta se volvió a tapar durante 10 días, después de los cuales, se efectuó una poda total de la planta para realizar el conteo de las ninfas que presentaban un punto o mancha negra en su cuerpo, lo mismo que las momias formadas. Las ninfas con la mancha negra se disectaron para determinar la presencia de la encapsulación que impide el desarrollo de todo estado inmaduro del parasitoide o su ausencia y, por lo tanto, si existían estados vivos e inmaduros del parasitoide. Por cada variable, se efectuaron cinco replicaciones.
Mortalidad de P. herreni por el daño que ocasiona la acción del parasitismo.
Se evaluaron los diferentes estados del piojo
En una hoja de yuca se pusieron 6 individuos de cada especie de piojo, se les adicionó un individuo hembra del parasitoide a evaluar y se realizaron observaciones durante una hora continua. Se registraron los siguientes eventos:
A- Localización del hospedero por el parasitoide; B- Intento de oviposición y C- Oviposición propiamente dicha, la cual se registraba al penetrar el ovipositor en el cuerpo del piojo. Se realizó el ensayo por separado para cada parasitoide.
Velocidad, umbral mínimo de temperatura y tiempo fisiológico de desarrollo
Teniendo en cuenta que el desarrollo de un organismo es dependiente de la temperatura, se estimó, para cada estado biológico y para cada especie, la velocidad de desarrollo a la temperatura evaluada. Esta se calculó, multiplicando por 100 el inverso del tiempo de desarrollo (días), es decir, 1/t.d. x 100.
Al expresarse gráficamente la velocidad de desarrollo en el eje “y” y la temperatura, en el eje “x”, se obtuvo una curva de tipo signoidea (S), cuya parte central es posible aproximar mediante una línea recta, la cual, al prolongarse, corta el eje “x” en el punto que corresponde al umbral mínimo de temperatura. Conociendo el valor de umbral mínimo, se puede realizar el cálculo de valor de la constante térmica o tiempo fisiológico (K), utilizando la siguiente forma:
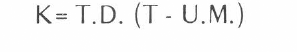
Donde: T.D. es el tiempo de desarrollo (días) en la temperatura (T), una de las temperaturas utilizadas del umbral mínimo (U.M.). En nuestro caso, para el cálculo del umbral mínimo, se utilizaron dos temperaturas, 20°C y 25°C. Se descartó 30°C, porque a esta temperatura, la tasa de desarrollo se encuentra en la fase que tiende a disminuir.
Especie de piojo harinoso por cada especie de parasitoide
Se tuvieron en cuenta los estados adultos de los piojos harinosos, P. herrani y
Resultados Y Discusion
Estado de P. herreni preferido por cada parasitoide.
Se presenta preferencia de cada parasitoide al estado de desarrollo de
En relación con
Teniendo en cuenta el estado preferido del piojo, estos resultados son importantes para el éxito del establecimiento de colonias, pues se le debe suministrar el estado preferido, pues, así, se asegura un alto parasitismo y se obtiene una población estable de la colonia.
Preferencia de los parasitoides por diferentes estados de
Datos obtenidos en una hora de exposición de parasitoide al hospedero.
Encapsulación de
Prueba de Duncan NS=0.05
Respuesta a la encapsulación.
En relación con la temperatura, la encapsulación se incrementa, cuando ésta disminuye, ya que a 30°C es de 3,6% y a 25°C, de 10,9%.
La encapsulación es fácilmente reconocible a través del cuerpo del piojo, por un punto negro brillante, que tiene una consistencia gelatinosa y no tiene un determinado sitio en el cuerpo del insecto y la cantidad de ellos no es uniforme en el mismo cuerpo.
Estos datos son de mucha importancia, ya que sabiendo que estos parasitoides tienen un menor riesgo de encapsulación en los estados avanzados de desarrollo y teniendo en cuenta la preferencia de los mismos, se deduce que estos factores están sincronizados para asegurar alta supervivencia de su próxima generación, a los cuales se agrega el factor ambiental o abiótico de la temperatura de 30°C, asegurándose una menor posibilidad de encapsulación.
En cuanto a A. coccois, este parásito no fue encapsulado por
Mortalidad de P. herreni por el daño que causa la acción de parasitismo.
En este aspecto se observa una diferencia entre las dos especies de parasitoides (Tabla 3). En el caso de E. diversicornis, se nota un efecto sobre el primer instar del hospedero, con un porcentaje de mortalidad de 13.2%, presentando diferencia significativa con los demás estados evaluados, lo mismo que entre el primer instar atacando y testigo (sin ataque), siendo lo contrario en los demás estados. A. coccois no presentó este efecto sobre P. herreni, comparado con el testigo.
Mortalidad de P. herreni por la acción de dos parasitoides.
Prueba de Duncan NS= 0.05
Velocidad de desarrollo, umbral mínimo de temperatura y tiempo fisiológico de desarrollo de los parasitoides.
Con la curva de desarrollo relacionada con la temperatura se pudo calcular el umbral mínimo de desarrollo (°C) debajo del cual el insecto detiene su metabolismo, lo cual se realizó para cada uno de los parasitoides, con los siguientes resultados: el umbral mínimo de desarrollo de
El tiempo fisiológico o constante térmica fue para
Los datos obtenidos (en días promedio) dan una idea de cuál es medio climático más apropiado para cada parasitoide para su liberación y la razón del por qué los dos parasitoides se encuentran en diferentes zonas ecológicas.
Efecto de la temperatura en el desarrollo de dos parasitoides de
Especie de piojo harinoso preferido por cada especie de parasitoide.
En este experimento se pudo comprobar la especificidad de los parasitoides, al ofrecerles ambas presas simultáneamente y al analízar, individualmente, los siguientes eventos: localización, intento de oviposición y la oviposición propiamente dicha (Tabla 5).
Esta preferencia es tan clara que, en el intento de oviposición.
Conclusiones
Bajo las condiciones en que se realizó este trabajo, se pudo llegar a las siguientes conclusiones:
Que ambos parasitoides tienen una alta preferencia hacia los estados más desarrollados del piojo harinoso, como son el tercer estado ninfal y el adulto hembra.
Que A. coccois no fue encapsulado por sus hospederos.
Que E. diversicornis es encapsulado por P. herreni en mayor porcentaje a 25°C en el segundo instar.
Que Ph. gossypii no encapsuló a ninguna de las dos especies de parasitoides.
Que E. diversicornis produce una mayor mortalidad en el primer instar, con un 13.2% y que A. coccois no produce mortalidad a los estados del piojo P. herreni.
Que, para E. diversicornis, el desarrollo de los parasitoides a 20°, 25° y 30°C fue de 35.2, 20.2 y 18.1 días promedio, respectivamente y, para A. coccois, de 39.5, 18.5 y 19.7 días promedio.
Que el umbral mínimo de temperatura fue de 13.5°C y 15.6°C y el tiempo fisiológico de desarrollo, 230.9°D y 175°D para E. diversicornis y A. coccois, respectivamente.
Que E. diversicornis tiene una clara preferencia por P. herreni y A. coccois, por P. gossypii.