
Abstract
The response of adult female of
Introduccion
Después de la segunda guerra mundial, creció la importancia de los ácaros fitófagos como plagas en plantas cultivadas de importancia comercial. Los miembros de la familia Tetranychidae son los de mayor incidencia y los que causan mayores daños. Esta explosión de grandes poblaciones açarinas ha sido asignada a diferentes causas, de las cuales, tal vez, las más importantes son el desarrollo de resistencia aproductos acaricidas y la eliminación de enemigos naturales debida al uso intensivo de plaguicidas, por ésto cada día se aumenta el interés en la aplicación de otros métodos de control de ácaros fitófagos y, en especial, el relacionado con el uso de enemigos naturales, lo cual se justifica si se tienen en cuenta el alto costo de los productos agroquímicos utilizados en la eliminación de las plagas de los cultivos.
Artrópodos depredadores son los controladores más importantes de ácaros fitófagos y, dentro de ellos, los ácaros de la familia Phytoseiidae son los enemigos naturales más efectivos, debido a la corta duración de su ciclo de vida y a las pocas necesidades de alimento para su sobrevivencia.
Muchos factores afectan la interacción depredador-presa, destacándose, como más importantes, la densidad del depredador y de la presa, las características del medio, de la presa y del depredador. Dos de estas variables, densidad del depredador y densidad de la presa, son características inherentes de cada situación depredador-presa, ya que originan los denominados componentes básicos de depredación, mientras que las tres restantes generan los componentes subsidiarios.
El presente trabajo constituye una de las partes de la evaluación funcional y numérica de los ácaros Phytoseiidae
Revision De Literatura
Un gran número de autores ha reconocido el valor de los ácaros depredadores de la familia Phytoseiidae en el control de ácaros fitófagos en general (Huffaker et al., 1970; Mc Murtry et al., 1970) y de los Tetranychidae, especialmente
Desafortunadamente, las características depredadoras de dichos organismos no han sido suficientemente estudiadas y ésto dificulta la identificación de los factores que afectan la interacción depredador-presa y la efectividad real de los Phytoseiidae como enemigos naturales.
La información para determinar la efectividad real de un depredador se ha basado, principalmente, en su habilidad para atacar la presa, la duración de su ciclo de vida con relación al de la presa y la tolerancia ambiental. A pesar de la obvia importancia de estas características, se reconoce ampliamente que, también, la capacidad de búsqueda es de vital importancia; por determinar la habilidad del depredador para localizar su presa y mantener su número en diferentes densidades de presa (Huffaker et al., 1970; Varley et al., 1973). No obstante lo anterior, muy pocos estudios han tenido en cuenta esta característica de depredación (Fleschner (1950) y Eveleigh and Chant (1982, b y c)).
Teóricamente, se ha demostrado que tres respuestas básicas de los depredadores tienen un efecto importante sobre la interacción depredador-presa: (1) Respuesta del depredador a la densidad de la presa, (2) Respuesta del depredador a la densidad del depredador y (3) Respuesta del depredador a la distribución de la presa (Hassell et al., 1976). Sin embargo, con la excepción de estudios teóricos, estas respuestas han sido tratadas como componentes separados de los procesos de depredación (Eveleigh y Chant, 1981 a).
Las respuestas de los depredadores a la densidad de la presa han sido divididas en dos tipos: (1) Respuesta funcional, definida por Holling (1961) y Hassell et al. (1976) como el cambio de la rata de ataque por depredador cuando cambia la densidad de la presa y (2) Respuesta numérica, que es el cambio en la densidad del depredador por cambios en la densidad de la presa. Holling describe tres tipos básicos de curvas para la respuesta funcional: Tipo I, caracterizada por un incremento lineal hasta alcanzar un pico a medida que se incrementa la densidad de la presa; Tipo II, con aceleración negativa hasta un pico máximo; y Tipo III en forma de “S” hasta alcanzar también un pico.
Las respuestas funcionales de los Phytoseiidae parecen ser variables, tanto intra, como interespecíficas y algunos estudios sugieren que, probablemente, sean multiformes (Mori, 1967; Sandness y Mc. Murtry, 1970). Varios factores internos o externos a los depredadores afectan la forma de la respuesta funcional, tales como: hambre, edad del depredador, aprendizaje, confusión, tamaño del depredador, posibilidades de presa alterna, posibilidades de dieta complementaria, tamaño, defensa y distribución de la presa. Por todo lo anterior, no es lógico esperar un solo tipo de curva como representativa del gran rango de respuestas del depredador a la densidad de la presa (Eveleigh and Chant, 1981a).
Los primeros pasos, en el sentido de integrar los diferentes componentes de depredación en la interacción depredador-presa de Phytoseiidae-Tetranychidae, han sido dados por las publicaciones de Eveleigh y Chant (1981 a, b; 1982 a,b,c), cuyos estudios han analizado un volumen bastante grande de 'tales componentes, a saber: respuestas funcional, numérica y total en diferentes estadios del depredador, emigración de depredadores y comportamientos de búsqueda bajo diferentes condiciones.
Materiales Y Metodos
Los patrones de distribución de
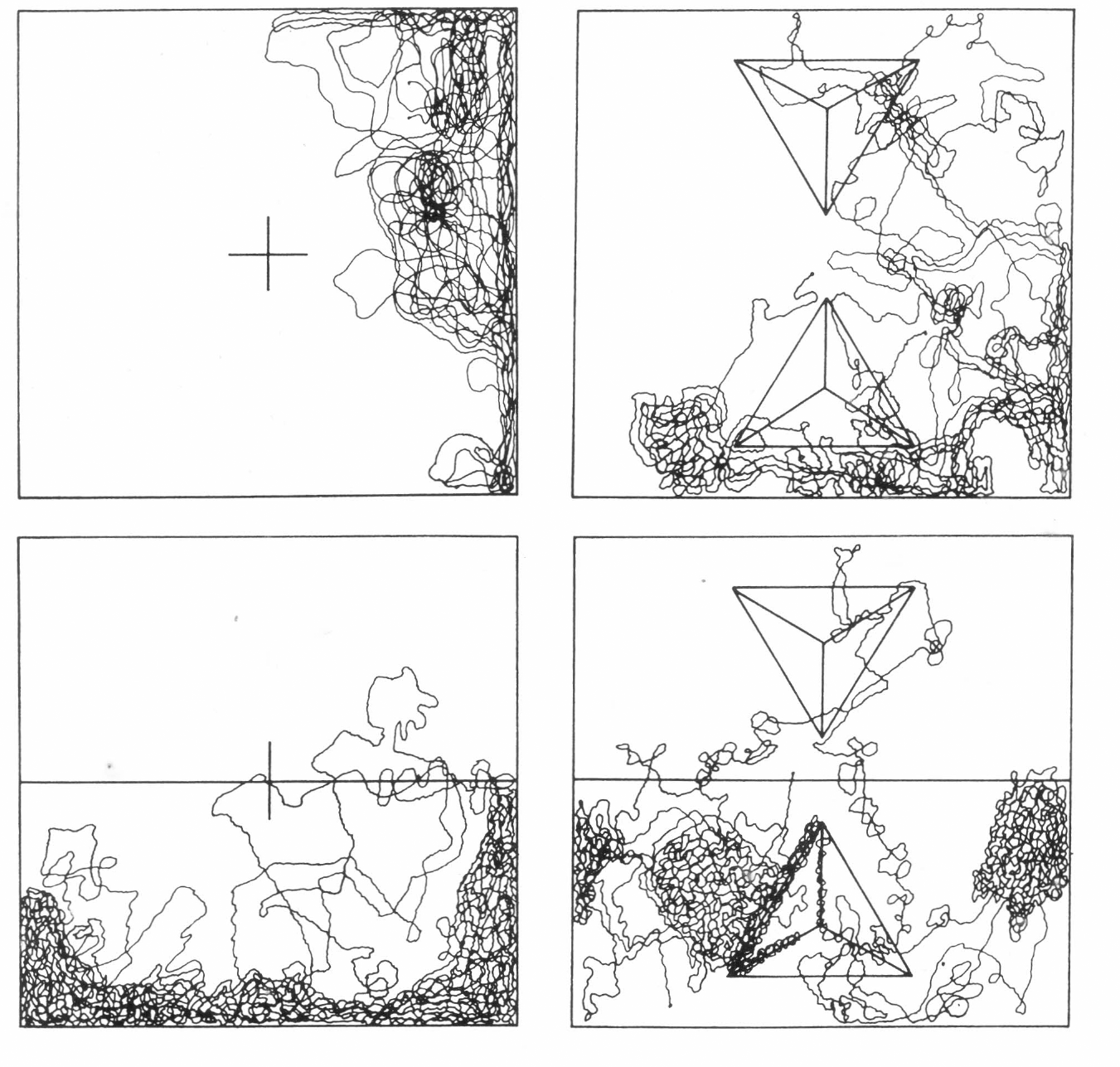
Todas las evaluaciones de los universos experimentales se llevaron a cabo en un cuarto oscuro con temperatura de 22°C y humedad relativa del 75%. La superficie de experimentación fue un cuadrado de papel milimetreado de 20 cm de lado; la iluminación se suministró con una lámpara de estereoscopio y sus variaciones (iluminación de solo la mitad del cuadrado) se lograron interponiendo un cartón negro mate entre la fuente y la superficie. La introducción de dos pirámides triangulares de cartón de 7 cm de lado y forradas con papel milimetrado sirvió para dar las variaciones topográficas.
Los organismos se resitringieron al cuadrado colocando una barrera de vaselina neutra en los bordes.
Los patrones de distribución de
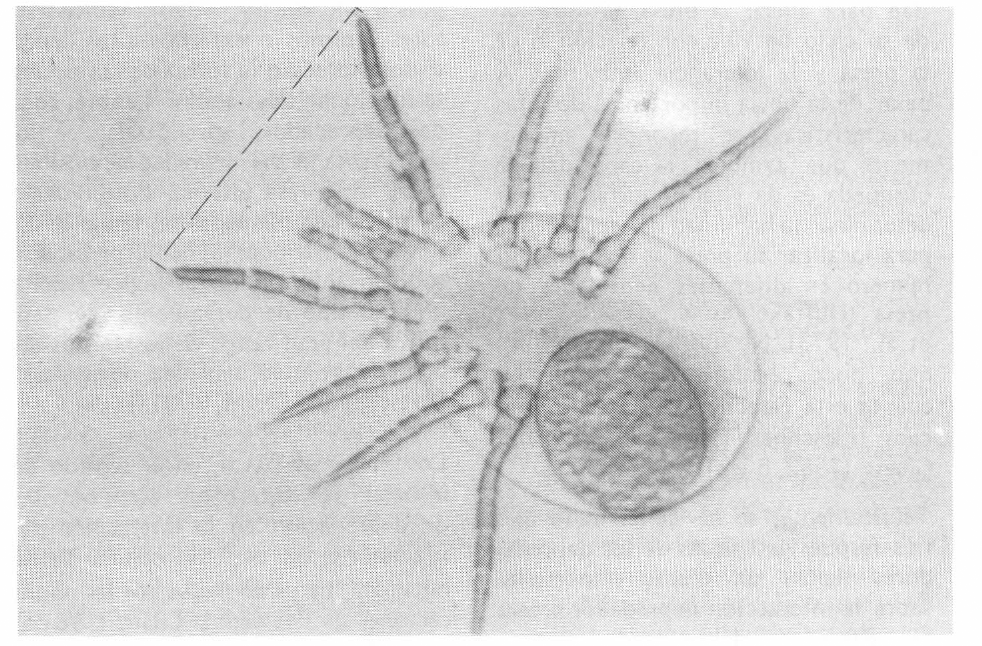
Los patrones de búsqueda para cada especie de depredador y cada universo se realizaron liberando una hembra grávida sobre la superficie y dibujando su desplazamiento durante una hora. Sobre los dibujos patrones así obtenidos y mediante la elaboración de calcos consecutivos hasta lograr desenredarlos completamente en líneas sencillas, se determinaron midiendo sobre una hoja de papel milimetrado las velocidades de búsqueda. Cada uno de los ejemplares evaluados y otros más fueron montados dorsalmente en solución Hoyer's, para su observación al microscopio y determinar el campo de percepción o distancia entre las uñas del par de patas I (Figura 2), con las cuales el depredador va barriendo la superficie por donde se desplaza en busca de su presa.

El área total de búsqueda de cada depredador para cada universo experi-.metal fue determinada por el producto entre la velocidad de búsqueda y el campo de percepción.
Para la determinación de la respuesta funcional de los depredadores a la densidad de la presa, se utilizaron discos de hoja para
Los discos de hoja para
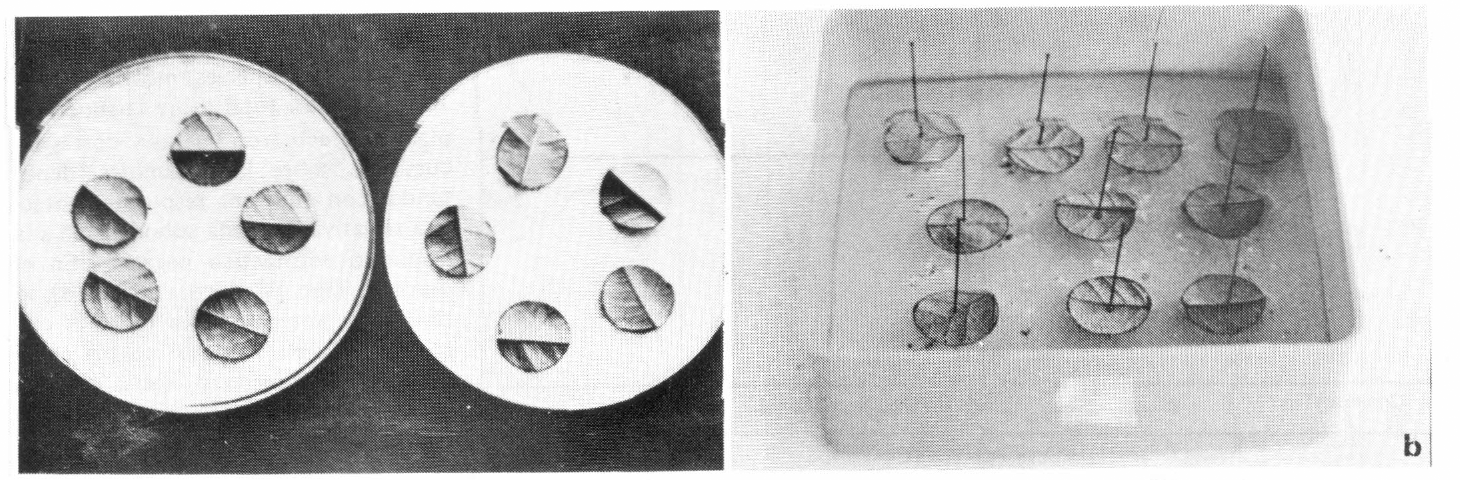
Para
Las evaluaciones de respuesta funcional se realizaron dentro de una cámara bioclimática con condiciones controladas de temperatura a 23± 2°C y humedad relativa de 85± 5%, diseñada y construida por los autores para tal fin (Figura 4). En un armazón de madera de 1,65 m de largo, por 0,8 m de alto y 0,8 m de fondo, se cubrieron las caras superior, inferior, anterior y posterior con plástico para invernadero y las caras laterales restantes fueron protegidas con lienzo, con el objeto de permitir la estabilización del microambiente y evitar la condensación de agua en el interior.
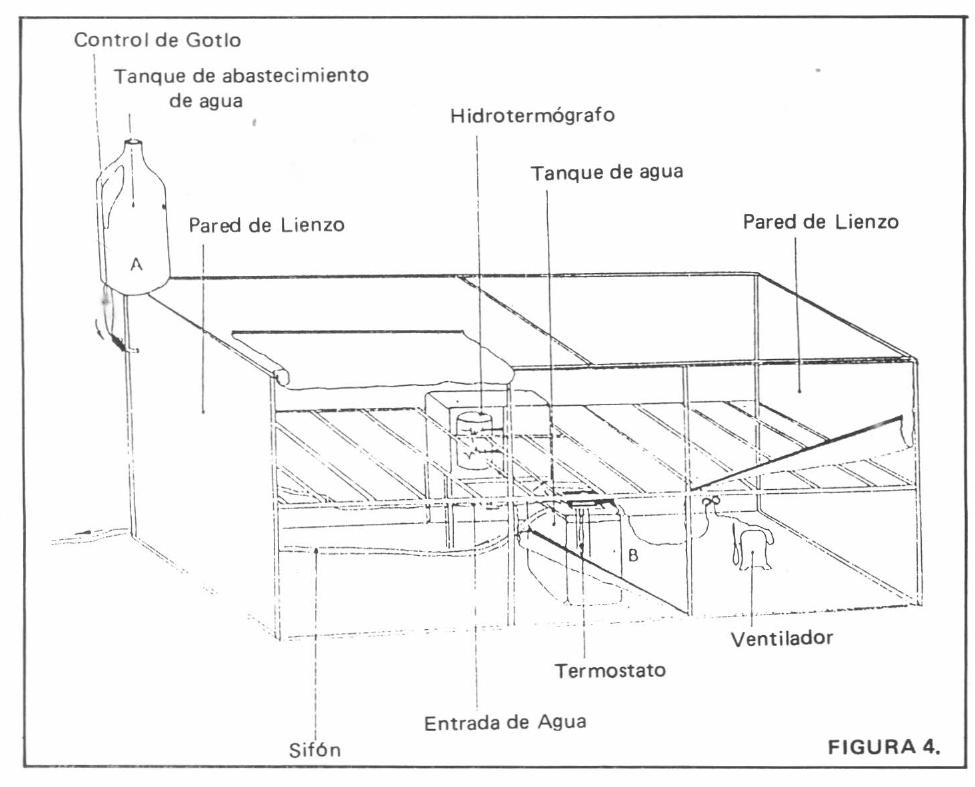
El sistema de regulación de temperatura y humedad relativa consistió en un recipiente plástico exterior (elemento A), colocado sobre la cámara y dotado de un equipo de venoclisis (microgoteo para dosificar la entrada de agua a un segundo recipiente plástico (elemento B) colocado dentro y adaptado con un sistema de sifón para drenar el exceso de líquido hacia el exterior. Un termostato para acuario, con regulador, marca Metaframe y modelo 22 Economatic, colocado dentro del último recipiente, mantuvo la temperatura y evaporación del agua constantes y, a la vez, la temperatura y humedad relativa dentro de la cámara. Con el fin de evitar estratificación de condiciones ambientales, se hizo circular constantemente el aire, mediante el uso de una hélice plástica, tipo ventilador, adaptada a un motor eléctrico de 4,5 voltios.
La cámara diseñada permite cierta amplitud en la escogencia de los valores de temperatura y humedad relativa, mediante la variación de la velocidad de goteo, la temperatura del termostato, la velocidad de la hélice ventiladora y la superficie de evaporación y el volumen de agua del elemento B.
Resultados Y Discusion
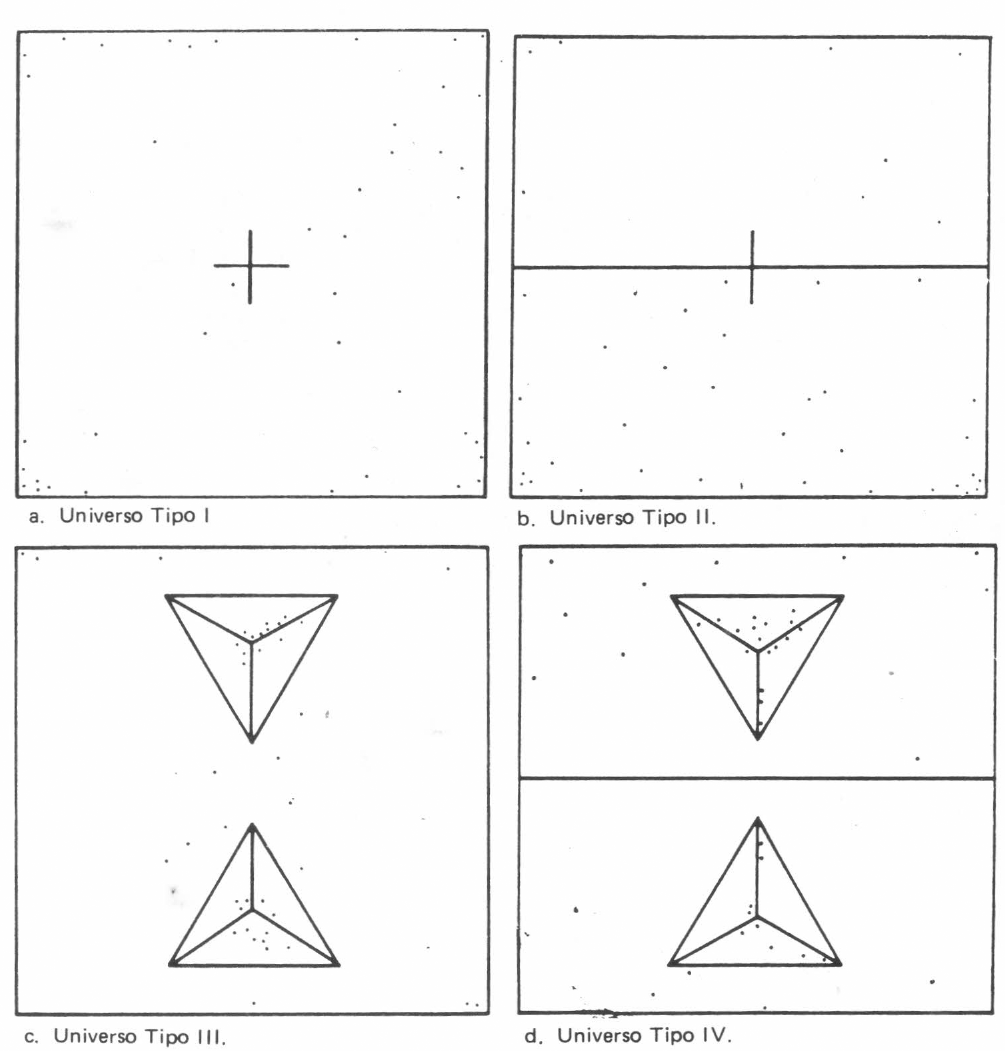
Patrones de Búsqueda de N. chilenensis.
En el universo Tipo III (Figura 5c), la respuesta a ascender, o respuesta geotáctica negativa, fue superior a la respuesta de escape la luz o fototactismo, ya que la mayoría de los especímenes se ubicaron sobre las pirámides y hacia los ápices y los individuos que permanecieron sobre la superficie plana respondieron de la misma manera que si estuvieran en el universo Tipo I. En el universo Tipo IV (Figura 5d), se observó más claridad, ya que la respuesta geotáctica negativa de las hembras de
En el universo Tipo III (Figuras 6c y 7c), las dos especies mostraron una búsqueda casi total sobre la superficie plana y efectuaron ligeras y cortas incursiones sobre las pirámides, denotando, con ello, una respuesta geotáctica negativa pequeña subordinada a la respuesta fototáctica negativa. En el universo Tipo IV (Figuras 6d y 7d), lo observado anteriormente fue más claro, pues la búsqueda se centró en la parte no iluminada y, sobre ésta, la superficie plana fue más transitada que la pirámide.
Superponiendo los patrones de distribución de
Areas de búsqueda de los depredadores
Las diferencias observadas fueron:

El campo de percepción, también, fue mayor para
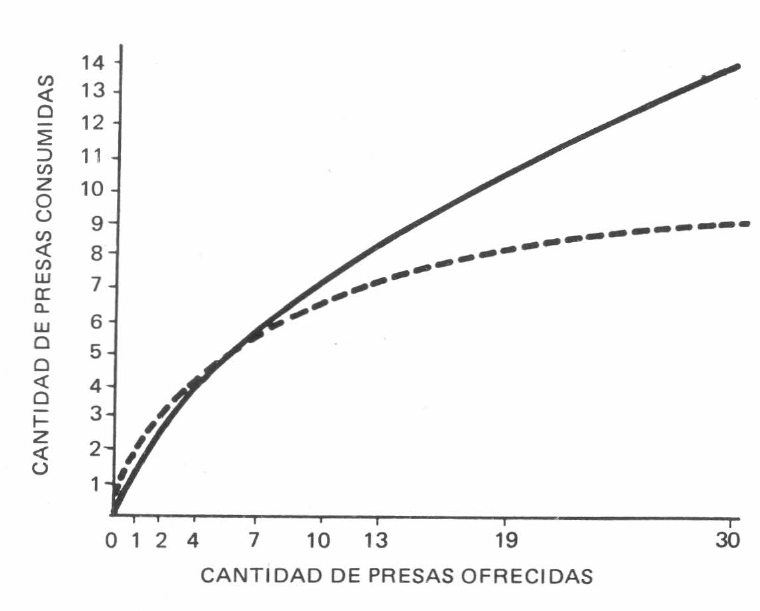
Comparación de respuestas funcionales de los ácaros Phitoseiidae (Amblyseius herbicolus (CHANT) y Neoseiulus chilenensis (DOSSE) devorando huevos de T. urticae (KOCH) sobre rosa.
Las hembras de
Conclusiones
En las condiciones del presente experimento, se concluye que:
Hembras adultas recién formadas de
Para
No obstante la diferencia de prioridad de respuestas de los depredadores en relación a las de la presa, las zonas coinciden suficientemente con las zonas preferidas por
En todos los universos experimentales
Las dos especies de depredadores, en densidades intermedias de 7,10 y 13 huevos, presentaron respuestas funcionales bastante parecidas.
Por presentar respuesta funcional a densidades bajas e intermedias de presa y no tener necesidades de dieta complementaria,
Es necesario determinar los umbrales de significancia económica para los daños causados por