
Abstract

Introduccion
Entre los insectos más interesantes se encuentran los himenópteros que agru- pan formas tan conocidas como las hormigas, las avispas y las abejas. Estas últimas constituyen la superfamilia Apoidea (Roth, 1973), cuyo número de especies pasa de 20.000 (Michener, 1979) agrupadas en 10 familias (Mi- chener, 1969, 1974; Batra, 1984). Las abejas se encuentran distribuidas en todo el mundo y las limitaciones para su distribución se dan sólo por la dis- ponibilidad de flora (Michener, 1979). Aunque es común considerar a todas las abejas como insectos sociales, re- sulta que más del 80% de éstas son solitarias (Batra, 1984).
Las familias Colletidae, Oxacidae, An- drenidae, Ctenoplectridae, Melittidae y Fidelidae comprenden especies de dis- tribución restringida, en general solita- rias y, excepto Fidelidae, de lengua corta, que los determina como visitan- tes de flores abiertas y planas, primi- tivas dentro de las Angiospermas. La familia Halictidae está ampliamente distribuida y comprende formas que alcanzan el grado social más primitivo (Michener, 1969); la familia Megachi- lidae, que incluye las abejas cortadoras de hojas, es de amplia distribución y comprende formas solitarias; la fami- lia Anthophoridae es cosmopolita y abarca un extenso número de especies solitarias, semisociales y parásitas (Linsley, 1958). La familia Apidae comprende a los abejorros principal- mente de las regiones templadas, a los euglosinos (neotropicales), a las abejas sin aguijón (pantropicales) y a la abeja melifera (diseminada en todo el mun- do por el hombre); esta familia com- prende formas prosociales (abejorros y euglosinos) y formas altamente socia- les (Michener, 1974). Las familias Fi- delidae, Megachilidae, Anthophoridae y Apidae poseen lengua larga y por lo tanto están adaptadas para tulizar flo- res de forma tubular, más evoluciona- das dentro de las Angiospermas (Eick- wort y Ginsberg, 1980; Kevan y Baker, 1983).
Los himenópteros superiores (Subor- den Clistogastra) se derivaron proba- blemente de un grupo parásito de plan- tas, parecido a las actuales moscas- sierra (Chalastogastra); este grupo pa- rásito, que provenía de himenóperos primitivos comedores de plantas, cons- tituyó la vía que condujo a los Clisto- gastra (Aculeata) que comprenden formas libres, parasitoideas y colecto- ras de polen (Lanham, 1951; Miche- ner, 1974). Las abejas, a su vez, se de- rivaron de un grupo de avispas esfe- coideas, considerándose a éstas como parafiléticas con respecto a las abejas (Lomholdt, 1982). Desde muy tempra- no, la dependencia de las abejas de néctar y polen suministrados por flores de plantas superiores, inició una com- pleja evolución dependiente entre abe- jas y Angiospermas (Baker y Hurd, 1968; Kevan y Baker, 1983).
La existencia de diversos grados de sociabilidad entre las abejas hacen de éstas un grupo ideal para estudios de biología social comparada, de incalcu- lables beneficios para la biología y ge- nética del comportamiento (Michener, 1969). Los diferentes niveles sociales en las abejas se relacionan en la Tabla 1.
Niveles de sociabilidad entre las abejas (Michener, 1969).
Las abejas carpinteras comprenden es- pecies de las Tribus Xylocopini y Cera- tinini, Subfamilia Xylocopinae, Fami- lia Anthophoridae. Las abejas del gé- nero Xylocopa se caracterizan por ni- dificar preferentemente en la madera. El género Proxylocopa, limitado a las zonas áridas del suroccidente de la región paleártica, nidifica en la tierra, considerándose el más primitivo de la Triby Xylocopini, y el género Lestis está limitado al oriente australiano.
El género Xylocopa es el más represen- tativo de la tribu Xylocopini; se en- cuentra en todo el mundo, principal- mente en los trópicos; en el Viejo Mundo se han descrito especies agrupa- das en 32 subgéneros y en el Nuevo Mundo especies de 17 subgéneros (Hurd y Moure, 1963). Las abejas car- pinteras construyen nidos dentro de la madera, haciendo galerías en diferen- tes direcciones a partir de una entrada común; para tratar la madera utilizan secreciones de sus glándulas mandibu- lares. En las galerías construyen celdas que aprovisionan de alimento; una vez colocado un huevo, sellan la celda con un tabique de aserrín y saliva (Batra, 1984). Entre estas abejas parece haber cuasisociabilidad (Tabla 1) teniendo en cuenta datos sobre xylocopas del Viejo Mundo (Michener, 1972) y resultados de este trabajo.
Según Hurd y Moure (1963), el factor más importante en la evolución de este grupo ha sido la disponibilidad de sus- trato para nidificar, más que la dispo- nibilidad de fuentes florales. Las abejas carpinteras son generalmente polilécti- cas (Linsley, 1958, 1961), polinizado- res principales de algunas plantas como maracuyá y té (Hurd y Moure, 1963), y plagas, al dañar flores no adaptadas para estos insectos y al utilizar madera de construcciones para nidificar (Lins- ley, 1958; Hurd y Moure, 1963; Kevan y Baker, 1983).
El parasitismo ejercido por avispas de la familia Sapygydae (Hurd y Moure, 1961) y por coleópteros meloideos (L.C. Pardo. Comunicación personal) puede afectar el crecimiento de sus po- blaciones (Watmough, 1983).
Materiales Y Metodos
El trabajo de campo se llevó a cabo en la Reserva Natural Nacional de "La Macarena", la cual se encuentra en el departamento del Meta; está limitada por la serranía del mismo nombre y por los ríos Güejar y Ariari al nor- oriente y Guayabero al sur. Su riqueza en flora y fauna se evidenció desde las primeras expediciones (Philipson, Don- caster e Idrobo, 1951); riqueza que, en parte, se explica por los pasados cam- bios climáticos (Vuilleumier, 1971).
Los trabajos de reconocimiento de la apifauna y observación de nidos se efectuaron en la región del río Güejar, próxima a la desembocadura del río en el Ariari, en la vereda "Caño Alfa", municipio de Puerto Rico. El paisaje característico está formado por suaves elevaciones y extensas llanuras de vege- tación abierta de escasa cobertura, no inundable en ninguna época del año; esta zona recibe el nombre local de "banqueta". La vegetación ribereña (selva de inundación) se conoce como "bajo" (Vargas, Rivera y Mendoza, en imprenta). El trabajo se efectuó en Enero de 1985, época seca de la región, con escasa humedad y altas tempera- turas. Los nidos se buscaron en el "bajo" y en la "banqueta".
En la descripción de cada nido se tiene en cuenta: 1. Localización: en qué tipo de zona se encuentra el nido y cómo se halla dispuesto. 2. Caracterís- ticas generales: tamaño y forma del nido, materiales con que está construi- do y estado en el momento de la ob- servación. 3. Estructura interna del nido. 4. Población: huevos, larvas, pu- pas, preadultos, adultos prereproducti- vos, adultos, parásitos. 5. Comporta- miento y nivel social.
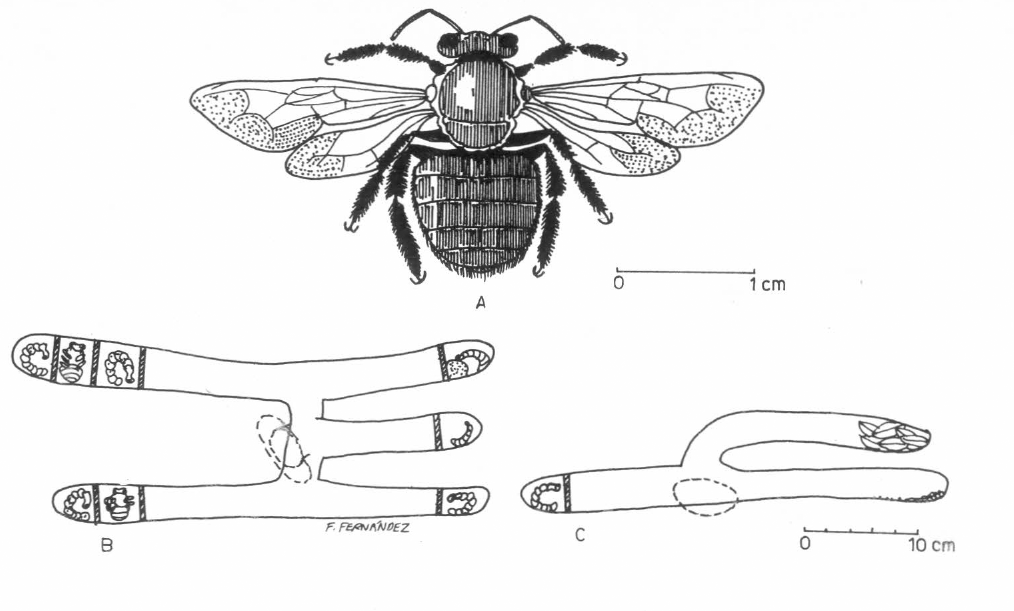
A. Hembra de
Resultados
En total se encontraron seis nidos de tres especies de
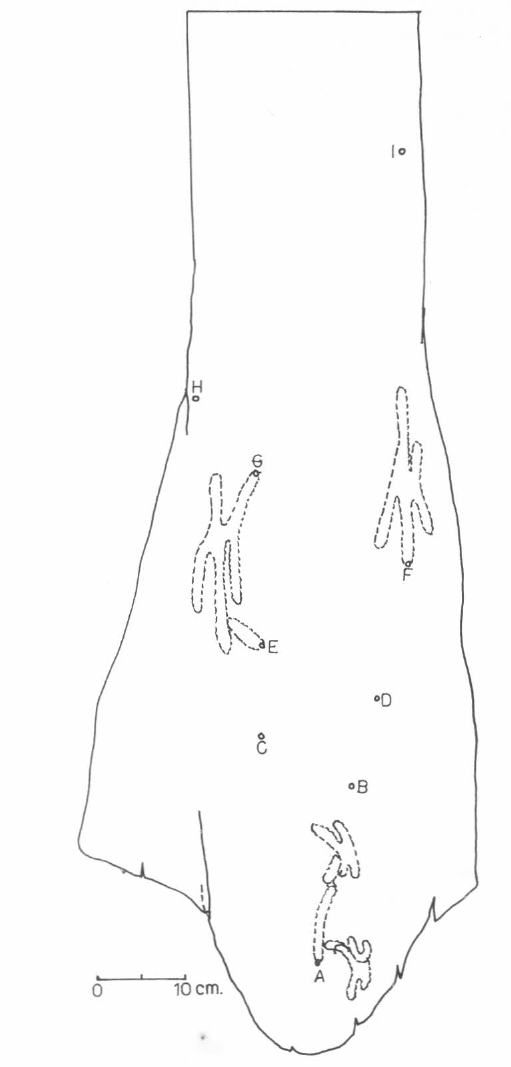
Ubicación de los cuatro nidos de
En el nido E (Fig. 3) correspondiente a la especie
Los nidos F, G y A corresponden a
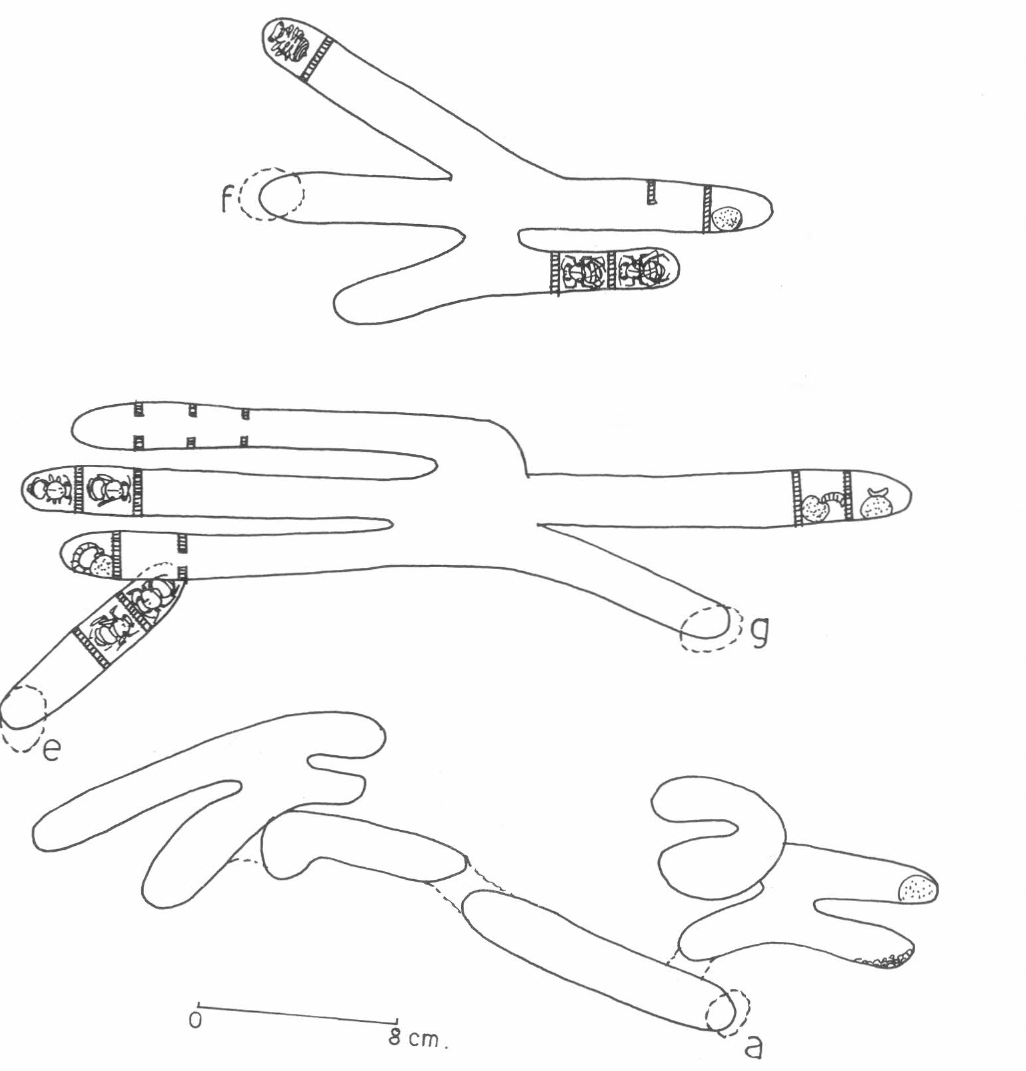
Esquema de los nidos encontrados en un árbol muerto en la "banqueta". E corresponde al nido de
El nido G (Fig. 3) contenía cinco gale- rías y cinco celdas selladas. Una con polen y un huevo, otra con polen y una larva alimentándose activamente, una tercerca celda con polen y una larva desarrollada devorada hasta la mitad por una larva no identificada; las dos últimas celdas contenían pupas. Se encontraron celdas abandonadas, 2 machos y 3 hembras adultas y 2 hem- bras recién emergidas.
El nido A (Fig. 3) contenía 11 galerías, una masa de polen en una de ellas y tierra en otra. Se capturaron dos hembras y un macho adulto.
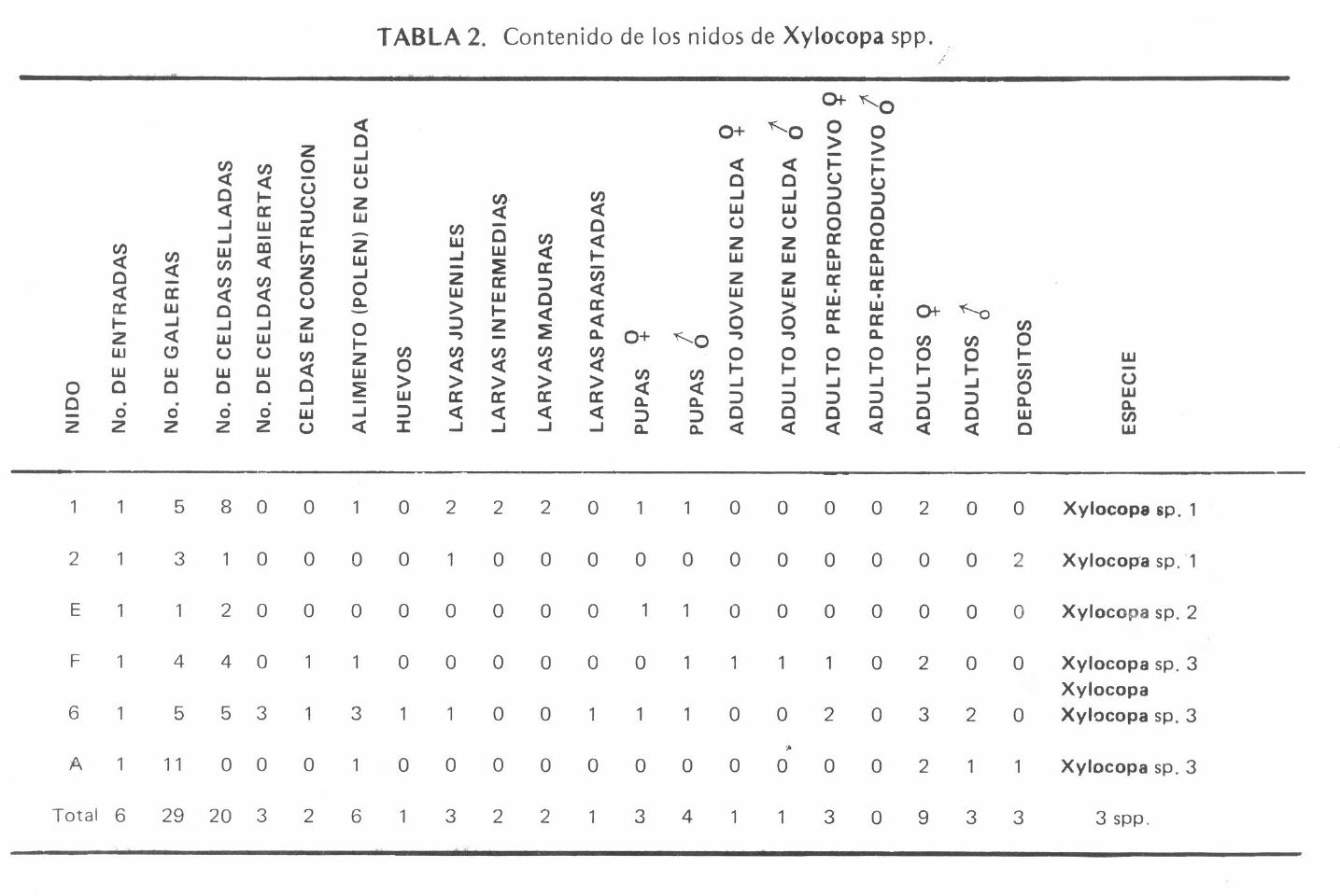
La Tabla 2 muestra el contenido de todos los nidos de
Discusion
Las abejas constituyen un vasto grupo con especies solitarias, parasociales y eusociales (Tabla 1). En general, las formas más primitivas son solitarias y las más recientes tienen algún grado de sociabilidad.
En la Introducción se señaló la impor- tancia de las flores para las abejas: el néctar y el polen que éstas poseen son esenciales para la nutrición de estos insectos (Michener, 1974). Las abejas están dotadas de adaptaciones morfo- fisiológicas únicas para la explotación de las flores (Thorp, 1979); por ello la evolución del grupo está íntimamente unida a la de las plantas con flores (Baker y Hurd, 1968) llegando a ex- tremos sorprendentes de dependencia insecto-planta como puede observarse en el trópico (Soderstrom y Calderon, 1971).
Contenido de los nidos de
En las abejas carpinteras, sin embargo, como ya se señaló, el factor importan- te en su evolución es la disponibilidad de sustrato para nidificar; el curso de la evolución de este grupo está y esta- rá asociado a aquellos eventos florísti- cos que afecten el sustrato para nidificar (Hurd y Moure, 1963). En América sólo el subgénero
Acerca del comportamiento nidifica- torio de las abejas carpinteras los datos son insuficientes para hacer aprecia- ciones generales. Según Hurd y Moure (1963), el subgénero
Puesto que existe, según Lindley (1958), cierta tendencia de la fauna del neártico a invadir el neotrópico, se espera encontrar en Colombia un puente entre las faunas del norte y del sur, una interesante variedad de formas y hábitos, teniendo en cuenta la di- versidad de ambientes que ofrece este país; sin embargo, la pobreza de estu- dios al respecto en Colombia impide en el momento apreciar la magnitud de tal variedad.
Las abejas carpinteras excavan en la madera valiéndose de secreciones de sus glándulas mandibulares y de sus poderosas mandíbulas; sin embargo, poco se sabe sobre la composición química de estas secreciones. Lo cierto es que estas abejas son capaces de abrirse paso aún en maderas duras. Estas abejas construyen varias galerías que parece seguir un patrón definido. Partiendo del fondo de cada galería, construyen las celdas, cámaras donde coloca una masa de polen y néctar mezclados, un huevo, un líquido en todo el piso de la celda que lo prote- gerá de microorganismos y finalmente, un tabique de aserrín y saliva mezcla- dos (Batra, 1984). En este estudio, el único huevo encontrado en el nido G midió 15 x 2 mm, siendo el huevo más grande entre las abejas y quizás entre todos los insectos (Michener, 1974). El nido F (Fig. 3) poseía una celda con alimento pero sin huevo; puesto que estaba sellada, puede tra- tarse de mortalidad a nivel del huevo cuya causa puede ser baja fertilidad de la hembra (Watmough, 1983). Larvas jóvenes, intermedias y maduras fueron observadas en varios nidos alimentán- dose activamente del polen, actividad que continuaron aún fuera de sus nidos.
En el nido G se encontró una larva de
En las pupas de
La presencia de adultos prereproducti- vos (desarrollados totalmente excepto las alas) y de machos adultos que no abandonan el nido pronto, encontra- dos en los nidos, toca un aspecto muy importante con respecto al comporta- miento de estas abejas. En primer lugar, la presencia en un nido de for- mas que van desde el huevo hasta el adulto, puede implicar la utilización del mismo nido por generaciones y la presencia de varias generaciones en un año (multivoltinismo) en oposición a una sola generación por año de espe- cies de
Al observar los niveles de sociabilidad entre las abejas (Tabla 1) se puede deducir que las abejas de los nidos des- critos se ajustan al nivel cuasisocial: no poseen castas ni división de labor, las colonias pueden ser o no matrifiliales, puede haber trabajo cooperativo en celdas, las hembras son estructural- mente similares y hay alimentación progresiva (regurgitación) a los adultos prereproductivos y machos que demo- ran en abandonar el nido, ya que la cría se alimenta siempre masivamente, es decir, se lo dota de alimento sufi- ciente para todo su desarrollo hasta pupa.
Es posible que en cada nido descrito se trate de más de un nido, donde las hembras simplemente utilicen una mis- ma entrada; sin embargo, en el nido G dos hembras y un macho adultos de los capturados se encontraron en una misma galería y en el nido E se encon- traron dos adultos en una única gale- ría. Estudios más detallados sobre el comportamiento de estas abejas pue- den despejar las incertidumbres que existen al respecto.
Conclusiones
La proximidad entre nidos de la misma especie y de especies diferentes, indica que no hay marcada territorialidad en la competencia por el sustrato.
El sustrato para nidificar fue siempre madera muerta. Cada nido poseía una sola entrada y el númro de galerías fue variable. Se encontraron todos los estados de desarrollo.
Se encontró parasitismo, posiblemente por una avispa sapigida.
Las abejas carpinteras de los nidos des- critos no son estrictamente solitarias; deducciones hechas sobre lo encontra- do las coloca como parasociales, posi- blemente en el nivel cuasisocial.
Aunque muchos nidos observados de
El conocimiento de los hábitos nidifi- catorios de estas abejas puede despejar incógnitas sobre la evolución y expan- sión del grupo en el neotrópico, espe- cialmente en Colombia, puente entre las faunas del norte y el sur de Améri- ca. Igualmente, puede crear bases racionales para el diseño de nidos arti- ficiales para programas de polinización dirigida.