
Abstract
The serine/threonine kinase PKB/Akt is a key mediator of survival and resistance to cancer therapy. Pharmacologic inhibition of Akt and its biologic sequelae may significantly impact the treatment of cancer. The use of molecular imaging technologies has contributed significantly to drug discovery research with an emphasis on drug efficacy, the mechanism of action, and target validation studies. We constructed a genetically engineered hybrid bioluminescent Akt reporter (BAR) molecule that reports on Akt serine/threonine kinase activity. Based on the fact that Akt is recruited to the plasma membrane on activation, we here describe a modified version of this reporter molecule (myristoylated and palmitoylated bioluminescent Akt reporter [MyrPalm-BAR]), which is membrane bound and whose bioluminescence activity can be used to monitor Akt activity at the cell membrane. Using changes in Akt activation status with small molecule inhibitors of Akt, we demonstrated that the membrane-targeted Akt reporter was more sensitive and quantitative. In addition, inhibition of upstream signaling kinases such as epidermal growth factor receptor and phosphatidylinositol 3-kinase activity resulted in changes in Akt activity that were quantitatively monitored by bioluminescence imaging. Based on these results, we propose that the membrane-associated Akt reporter may be better suited for identification of novel compounds that modulate the Akt pathway by high-throughput screening.
GENETIC ALTERATIONS that result in changes within signaling pathways have been implicated in tumor initiation, tumor growth, and resistance to cancer therapy. In particular, amplification and/or overexpression of receptor tyrosine kinases (RTKs), of which epidermal growth factor receptor (EGFR) and HER2 are prototypic examples, have been implicated as initiating events in a variety of human malignancies. 1 These kinases and their downstream signaling kinases therefore make a very attractive target for therapeutic interventions. Indeed, recent clinical success of trastuzumab in HER2-overexpressing breast cancer2,3 and erlotinib in EGFR-overexpressing cancers, 4 as well as imatinib mesylate (Gleevec) for chronic myeloid leukemia, 5 provides the impetus for targeted inhibition of kinases as a viable therapeutic paradigm. The serine/threonine kinase PKB/Akt is a key mediator of survival and resistance to therapy. Akt functions as a signaling hub wherein many upstream signaling pathways, such as those activated in response to RTK activation, eventually converge. The integration of these intracellular signals at the level of Akt and its kinase activity regulates the phosphorylation of several downstream effectors, such as nuclear factor κB, mTOR, Forkhead, Bad, GSK-3, and MDM-2.6–12 These phosphorylation events in turn mediate the effects of Akt on cell growth, proliferation, protection from proapoptotic stimuli, and stimulation of neoangiogenesis. 13 The activation of Akt is regulated by both translocation to the plasma membrane and phosphorylation at Thr308 and Ser473. Constitutive phosphorylation of Akt at these sites is frequently observed in a wide range of solid tumors and hematologic malignancies. Ongoing efforts have focused on therapeutically targeting Akt and its biologic sequelae, either at the level of Akt itself or at the levels of its upstream regulators and downstream effectors. 14 Because Akt is also important for proliferative and antiapoptotic signaling pathways critical for normal cells, particular emphasis is placed on fine-tuning the targeting of individual components of this pathway to maximize the therapeutic index of anticancer strategies. Although there have been major developments in our understanding of the biology of Akt and its role in human malignancies, the development of molecular imaging technologies to monitor and quantify Akt activity is still in its infancy.
Noninvasive technologies such as magnetic resonance imaging, magnetic resonance spectroscopy, positron emission tomography, and optical imaging will contribute significantly to drug discovery research with an emphasis on drug efficacy, the mechanism of action, and target validation studies in animal disease models.15–24 Molecular imaging techniques bridge the gap between preclinical and clinical research for the development of successful drug candidates that have target specificity, optimal safety, pharmacokinetics/pharmacodynamics, and efficacy.25–28 The use of molecular imaging end points instead of time-consuming dissection and histology significantly decreases the workload involved in tissue analysis and thereby speeds up the evaluation of drug candidates. As imaging methods are noninvasive, they allow for longitudinal studies in a single animal. This increases the statistical relevance of a study, allows for more clinically relevant study designs, and decreases the number of animals required. These tools also provide much earlier surrogate markers of therapeutic success and thus speed up the process of drug discovery and clinical evaluation.
In previous studies, we described a reporter molecule whose expression provides the ability to noninvasively image Akt activity using bioluminescence imaging (BLI). 29 This bioluminescent Akt reporter (BAR) was designed to be expressed within the cytosolic compartment. Based on the reasoning that Akt, when activated, is recruited to the plasma membrane, here we describe the development of a membrane-targeted bioluminescent Akt reporter (myristoylated and palmitoylated bioluminescent Akt reporter [MyrPalm-BAR]). Fusion of 10 amino-terminal residues of Lyn, which enables recruitment of Lyn to the plasma membrane by virtue of myristoylation and palmitoylation, 30 to the amino-terminus BAR resulted in membrane association of the reporter. Using various inhibitors of the Akt pathway, we demonstrate that membrane-targeting BAR is a more sensitive reporter of Akt activity. MyrPalm-BAR would therefore be better suited for studies wherein Akt activity was being evaluated in a biologic context as well as studies for the identification of novel compounds targeting the Akt pathway.
Methods
Gene Construction and DNA Plasmid
MyrPalm-BAR reporter was cloned in the mammalian expression vector pEF. 31 MyrPalm-BAR was generated by adding the 5’ 30 bp (animo acids M to D) of Lyn kinase to the 5’ end of the BAR complementary deoxyribonucleic acid (DNA). The pEF-BAR plasmid has been described previously. 29
Cell Culture and Transfection
D54 (human glioma) cells was maintained in RPMI (Gibco, Carlsbad, CA) supplemented with 10% fetal bovine serum (Gibco). To generate stable cell lines, the MyrPalm-BAR plasmid was transfected into D54 cells using Fugene (Roche Diagnostics, Indianapolis, IN), and resulting stable clones were selected using 200 µg/mL G418 (Invitrogen, Carlsbad, CA).
Antibodies and Chemicals
Rabbit polyclonal antibodies to Akt, phospho-Akt (phosphorylated on Ser473), and phospho-EGFR (Y845) were purchased from Cell Signaling Technology (Danvers, MA), and rabbit polyclonal antibody to EGFR (SC-03) was purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Perifosine was purchased from Cayman Chemical (Ann Arbor, MI). LY294002 was obtained from Cell Signaling Technology (Danvers, MA). Luciferin was obtained from Biosynth (Staad, Switzerland). Erlotinib was kindly provided by Genentech (San Francisco, CA). 2-Bromopalmitic acid (2-BP) was purchased from (Sigma, St. Louis, MO).
Immunostaining
Cells grown on a coverslip were fixed with 3.5% paraformaldehyde in phosphate-buffered saline for 15 minutes at room temperature. Chilled methanol was then added, and cells were incubated at −20°C for 10 minutes. Coverslips were then blocked using 10% donkey serum and incubated with 1:50 antiluciferase antibodies for 1 hour at 37°C in a humidified environment. After washing, coverslips were incubated with donkey antigoat antibody coupled to Cy3 (Jackson ImmunoResearch, West Grove, PA) and were analyzed using a Nikon fluorescence microscope using filters with an excitation wavelength of 533 nm.
Bioluminescence Imaging
Live-cell luminescent imaging was achieved by adding
Western Blotting Analysis
Cell lysates were prepared using NP40 lysis buffer (1% NP40, 150 mM NaCl, 25 mM Tris pH 8.0) supplemented with protease inhibitors (Calbiochem, San Diego, CA) and phosphatase inhibitors (Sigma). Proteins were resolved by SDS/PAGE and analyzed by Western blotting using appropriate antibodies. Bound antibody was detected using horseradish peroxidase–conjugated secondary antibodies and enhanced chemiluminescence (Amersham Pharmacia, Uppsala, Sweden).
Data Analysis
Percent changes in signal intensity were calculated using pretreatment values as baseline and plotted as mean ± SEM for each group. Statistical comparisons were made using the unpaired Student t-test, with a value of p < .05 being the cutoff for significance.
Results
Construction of Membrane-Targeting Akt Reporter
Akt/PKB is recruited to the plasma membrane following production of phosphatidylinositol (3,4,5) trisphosphate and phosphatidylinositol (3,4) bisphosphate. At the cell membrane, Akt is activated on phosphorylation. 13 To investigate if membrane recruitment of the BAR would enhance the sensitivity of the reporter, we constructed a membrane-targeted bioluminescent Akt reporter (MyrPalm-BAR) by fusing 10 amino-terminal residues of the Lyn kinase to BAR (Figure 1A). This sequence contains signals for myristoylation and palmitoylation, which have been previously demonstrated to effectively target a nonmembrane protein to the plasma membrane. 30 The functional basis of the reporter, which is similar to that previously described, is shown schematically in Figure 1B. In the presence of Akt kinase activity, phosphorylation of Akt consensus substrate sequence within the reporter would result in its interaction with the FHA2 domain, thus stearically preventing reconstitution of the luciferase reporter molecule. In the absence of Akt kinase activity, loss of this stearic constraint would allow reconstitution of the luciferase reporter molecule, whose activity can be detected noninvasively by BLI.
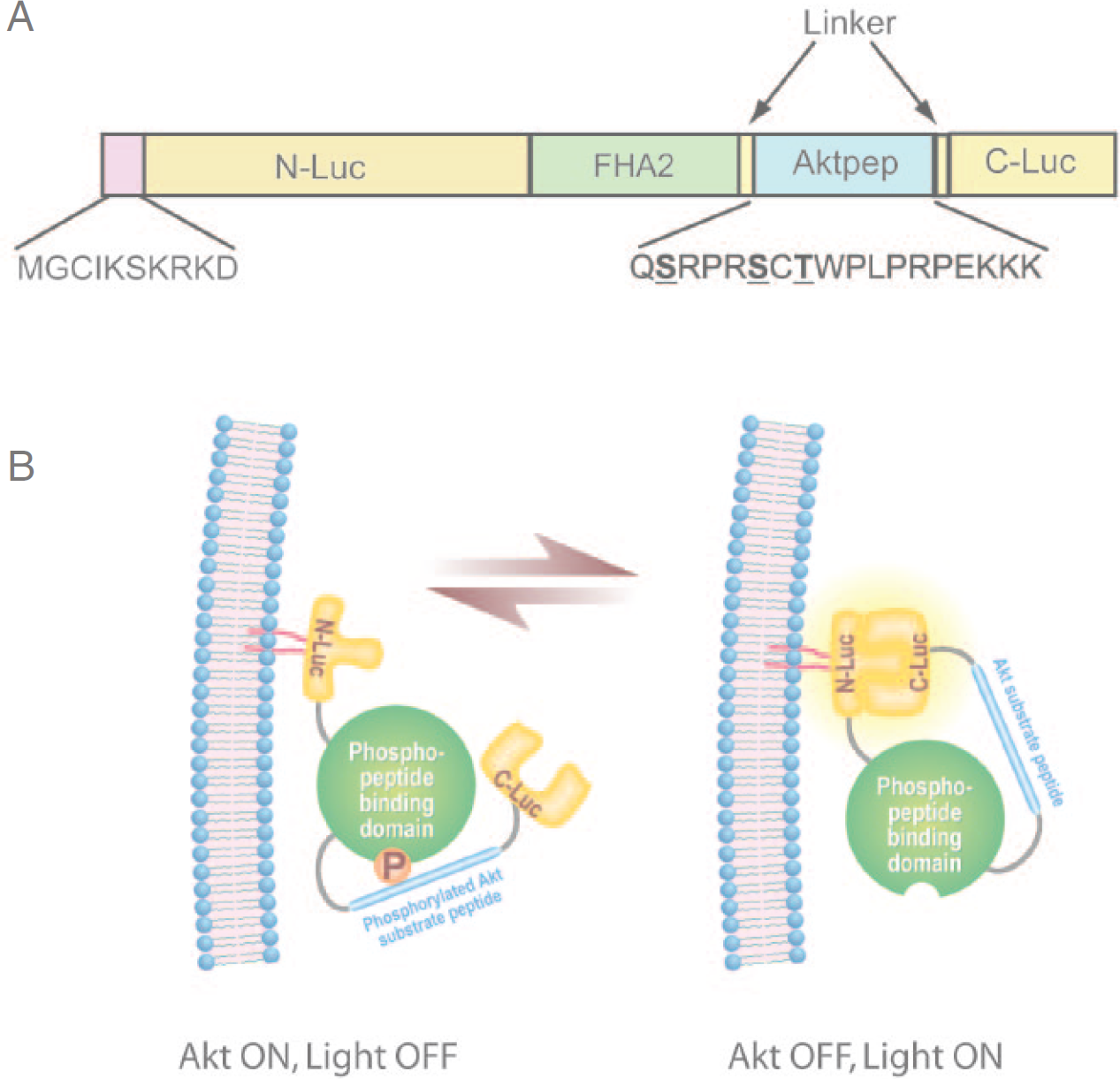
MyrPalm-BAR. A, The domain structure representation of MyrPalm-BAR. MyrPalm-BAR was generated by adding 10 amino acids from the N terminus of Lyn kinase to the BAR complementary DNA. N-Luc and C-Luc are the animo- and carboxyl-terminal domains of firefly luciferase that were fused to the appropriate ends of the reporter. 29 The Aktpep domain constitutes a consensus Akt substrate sequence. 33 On either side of the substrate sequence, flexible linker sequence was included (GGSGG). At the amino-terminal of the Aktpep domain, the yeast FHA2 phospho-Ser/Thr binding domain (residues 420 to 582) 30 was included. B, The proposed mechanism of action for the MyrPalm-BAR reporter involves Akt-dependent phosphorylation of the Aktpep domain, which results in its interaction with the FHA2 domain. In this form (Akt-ON), the reporter has minimal bioluminescence activity. In the absence of Akt activity, association of the N-Luc and C-Luc domains restores bioluminescence activity owing to the loss of interaction between Aktpep and FHA2.
MyrPalm-BAR Is Membrane Located
The expression plasmid for this hybrid reporter, pEF-MyrPalm-BAR, was stably transfected to D54 human glioma cells, and resulting clones (D54-MyrPalm-BAR) were analyzed by Western blot. D54-MyrPalm-BAR and D54-BAR yielded an 84 kDa antiluciferase reactive polypeptide. Mock transfected D54 cells failed to show a luciferase-specific polypeptide (Figure 2A).
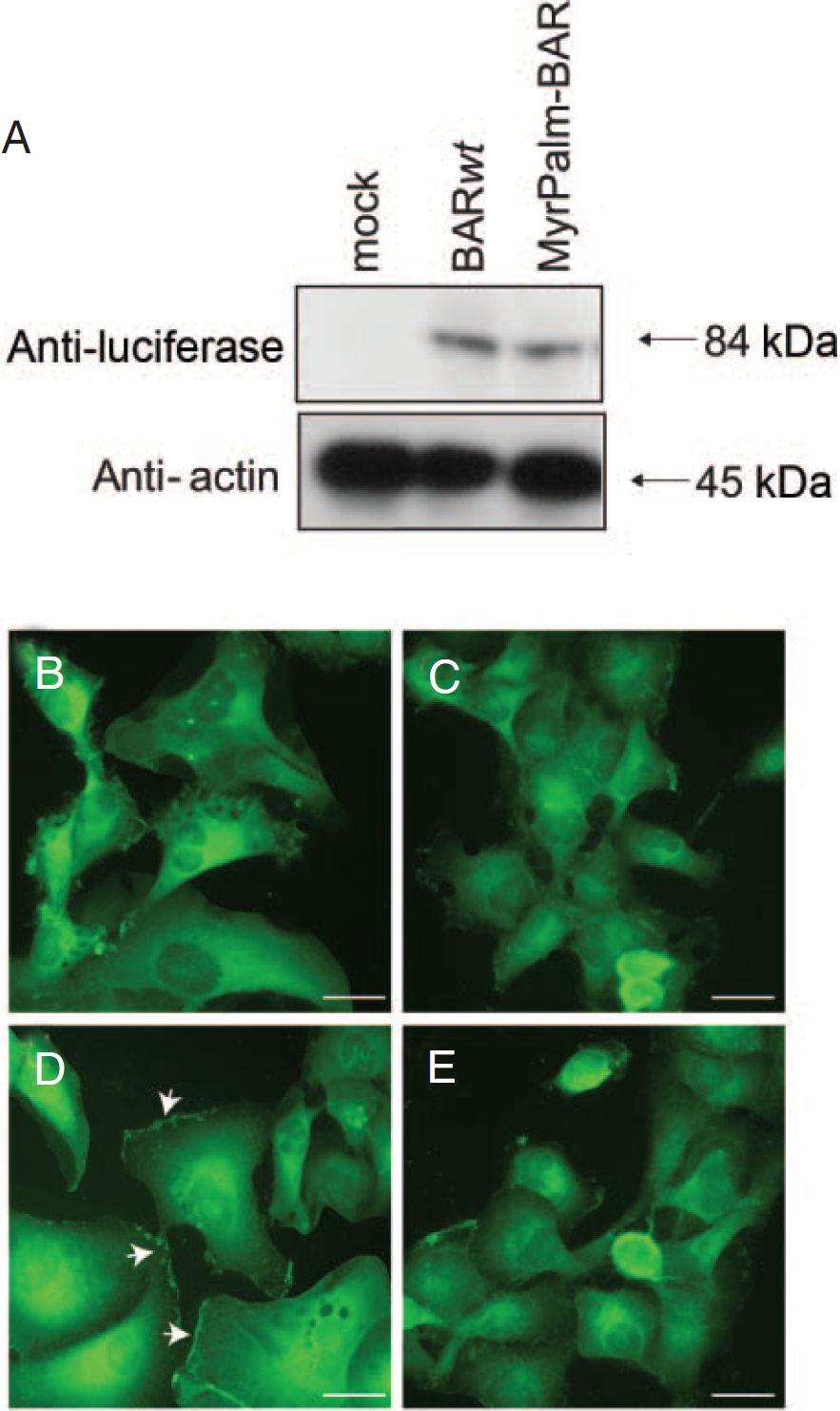
Subcellular localization of MyrPalm-BAR. A, D54 cells expressing MyrPalm-BAR and control D54 cells expressing BAR were analyzed by Western blotting using antibodies specific for luciferase or actin as a loading control. D54-BAR cells treated with control vehicle (0.1% dimethyl sulfoxide [DMSO] in the medium) (B) or 10 µM 2-bromopalmitate overnight (C) and D54-MyrPalm-BAR cells treated with control vehicle (0.1% DMSO in the medium) (D) or 10 µM 2-bromopalmitate overnight (E) were fixed, permeabilized, and immunostained with luciferase antibody and monitored under a fluorescence microscope.
To confirm the membrane targeting of the reporter, we examined the subcellular distribution of MyrPalm-BAR in D54 cells and compared it with BAR localization by immunofluorescence microscopy. The distinct membrane staining with luciferase antibody was seen only in cells expressing MyrPalm-BAR, whereas BAR was predominantly localized in the cytoplasm (Figure 2, B and D). To address the requirement of palmitoylation for targeting to plasma membrane of MyrPalm-BAR, D54-MyrPalm-BAR cells were treated with the palmitoylation inhibitor 2-BP. 32 Such treatment of D54-MyrPalm-BAR cells resulted in the loss of membrane fluorescence signal (Figure 2E). This effect was specific for palmitoylated BAR because the distribution of BAR that lacks a palmitoylatable N-terminal sequence remained the same in the absence and presence of 2-BP treatment (Figure 2C).
MyrPalm-BAR Is More Sensitive to Akt Activity
To compare the sensitivity of MyrPalm-BAR and BAR to Akt inhibition, we imaged D54-MyrPalm-BAR and D54-BAR cells after treatment with an Akt inhibitor, perifosine. Treatment of D54-BAR cells resulted in a twofold increase in bioluminescence activity, whereas treatment of D54-MyrPalm-BAR cells resulted in a fivefold increase in bioluminescence activity (Figure 3, A and B). Western blotting analysis confirmed that perifosine treatment resulted in a similar decrease in phospho-Akt levels in both cell lines (Figure 3C). We further treated D54-MyrPalm-BAR with LY294002, a phosphatidylinositol 3-kinase (PI-3K) inhibitor, which resulted in a ninefold increase in bioluminescence activity. However, treatment of D54-BAR cells resulted in a sixfold increase in bioluminescence activity (Figure 3, D and E). These changes in bioluminescence activity correlated with phospho-Akt as determined by Western blotting analysis (Figure 3F).

Imaging of Akt activity. A, D54 cells expressing MyrPalm-BAR and D54 cells expressing BAR were treated with 40 µM perifosine or control vehicle (0.1% phosphate-buffered saline in the medium) for 1 hour. The changes in bioluminescence activity compared with the pretreatment value were plotted as fold induction. Data were derived from a minimum of five experiments. B, Images of representative cells of A are shown before and after treatment. C, Cells from one of the experiments of A were used for Western blotting analysis with antibodies specific for phospho-Akt or total Akt. D, D54-MyrPalm-BAR cells and D54-BAR cells were treated with 50 µM LY294002 or control vehicle (0.1% dimethyl sulfoxide in the medium) for 1 hour, and the bioluminescence activity before and after treatment was compared and the changes were plotted as fold induction. Data were derived from a minimum of five experiments. E, Images of representative cells from D are shown before and after treatment. F, Western blotting analysis of samples from D using antibodies specific for phospho-Akt or total Akt.
Monitor EGFR Activity by MyrPalm-BAR
We further investigated the ability of the MyrPalm-BAR to detect changes in Akt status on inhibition of upstream signal from growth factor receptors. D54-MyrPalm-BAR and D54-BAR cells were treated with the EGFR inhibitor erlotinib, and the increase in the BLI of reporters was monitored. D54-BAR cells showed a 5-fold induction of BLI activity, whereas D54-MyrPalm-BAR cells displayed an 11-fold increase in BLI activity within 1 hour of erlotinib treatment (Figure 4A). Figure 4C demonstrates that this discrepancy in BLI was not due to differences in the sensitivity of the two cell lines to erlotinib since changes in phospho-Akt levels and phospho-EGFR levels were comparable. To verify that increased sensitivity of MyrPalm-BAR is due to its membrane targeting ability, we pretreated both D54-MyrPalm-BAR and D54-BAR cells with the palmitoylation inhibitor 2-BP and then monitored the induction of BLI of reporter activated by erlotinib. The pretreatment D54-MyrPalm-BAR cells with 2-BP resulted in a lower-fold induction, whereas D54-BAR cells had a similar induction in bioluminescence activity in the presence or absence of 2-BP.

Molecular imaging of receptor tyrosine kinase. A, D54 cells expressing MyrPalm-BAR and D54 cells expressing BAR were preincubated overnight with or without 10 µM 2-BP, treated with 10 µM erlotinib or control vehicle (0.1% dimethyl sulfoxide in the medium), and imaged after 1 hour. The changes in bioluminescence activity compared with the pretreatment value were plotted as fold induction. B, Images of representative cells from A are shown before and after treatment. C, Western blotting analysis of samples from A using antibodies specific for phospho–epidermal growth factor receptor (EGFR), total EGFR, phospho-Akt, or total Akt.
Discussion
In our previous studies, we developed a split firefly luciferase-based reporter (BAR) wherein Akt activity can be detected by BLI. We showed that the inhibition of Akt activity using various kinase inhibitors resulted in an increase in bioluminescence activity in a time- and dose-dependent manner, which indicated that BAR provides a surrogate for Akt activity in terms of quantity and dynamics. In the present study, we modified BAR by fusing the 10 amino-terminal residues of Lyn kinase. This sequence contains signals for myristorylation and palmitoylation, effectively targeting BAR to the cell membrane, where the activated form of Akt is recruited.
Using an Akt inhibitor and a PI-3K inhibitor, the sensitivity of BAR and MyrPalm-BAR reporter was studied by BLI, as well as by traditional Western blotting analysis (see Figure 3). The MyrPalm-BAR reporter showed a higher-fold induction in bioluminescence than did the BAR reporter. These results suggested that MyrPalm-BAR is more sensitive than BAR. Furthermore, we evaluated the erlotinib-mediated inhibition of EGFR using both reporters. When compared with BAR, MyrPalm-BAR generated a higher increase in bioluminescence, and pretreatment of a palmitoylation inhibitor, 2-BP, resulted in a lower activation of MyrPalm-BAR. Taken together, these results suggested that the membrane targeting ability of MyrPalm-BAR confers increased sensitivity in reporting Akt activity. Targeting of kinase reporters to appropriate subcellular compartments has been successfully used previously to enhance their sensitivity. 30
Molecular imaging of Akt in cells and live animals will greatly facilitate the process of target validation, dose and schedule optimization, and rapid identification of novel compounds from a library of inhibitors using cell-based high-throughput screening. We suggest that since the signal to noise ratio of MyrPalm-BAR is higher than BAR alone, the membrane-targeted Akt reporter may be better suited for high-throughput screening to identify Akt inhibitors.