
Abstract
The multifunctional growth factor scatter factor/hepatocyte growth factor and its tyrosine kinase receptor, c-MET, have been implicated in the genesis and malignant progression of numerous human malignancies, including hepatocellular carcinomas. The incidence of hepatocellular carcinomas in the United States has increased noticeably over the past two decades and is listed as the fifth major cancer in men worldwide. In this study, we used a choline-deficient
The multifunctional growth factor scatter factor–hepatocyte growth factor (HGF) and its tyrosine kinase receptor, c-MET, have been implicated in the genesis and malignant progression of multiple human malignancies. c-MET is crucially involved in invasive cell growth and motility during embryogenesis and has been detected in invasive tumors, including hepatocarcinomas, gliomas, meningiomas, colorectal cancer, uveal melanomas, thyroid carcinomas, breast carcinomas, gastric carcinomas, and sarcomas.1–11
Imaging science implies the formation and display of the spatial distributions of material properties, and in biomedical applications, these properties provide uniquely valuable information about tissue composition, morphology, and function, as well as quantitative information of many underlying biologic processes. With the use of magnetic resonance imaging (MRI), these processes can be studied in vivo and noninvasively, allowing serial studies in experimental animals and humans to be conducted. MRI, the images of which are constructed on the water content of the body, has considerably improved our ability to detect and localize lesions. MRI procedures have helped somewhat with pathologic diagnosis and lesion characterization to a certain extent; however, there is often a lack of molecule-specific information.
Recently, new and more powerful MRI technologies for specific molecular targeting or tagging have been used for in vivo visualization of cell surface antigens and/or receptors. This method relies on the specific labeling of extracellular cell surface receptors or antigens with a targeted contrast agent. The contrast agent MRI probe can be targeted to a specific receptor or antigen by a monoclonal antibody that binds with high affinity to the receptor or antigen. For instance, this approach has been successfully used to image with MRI the neovasculature in angiogenic tumors with gadolinium (Gd)-labeled polymerized liposomes targeted against the αvβ3 integrin expressed on neovascular endothelium.12,13 Konda and colleagues used a polyamidoamine folate-dendrimer conjugated to folic acid and Gd– diethylenetriaminepentaacetic acid (DTPA) to specifically target the high-affinity folate receptor, overexpressed in more than 80% of ovarian tumors, in vitro in mouse erythroleukemia cells and in vivo in ovarian tumor xenografts, as another approach to amplify the amount of Gd reaching the tumor site. 14 Artemov and colleagues used avidin-Gd-DTPA complexes targeted for tumor cells prelabeled with a biotinylated anti-Her-2/neu monoclonal antibody to obtain in vivo T1-weighted MRIs of Her2/neuexpressing tumors inoculated subcutaneously in severe combined immunodeficiency mice. 15 Vera and colleagues used a Gd complex of polydiethylenetriamine pentaacetic acid polyneogalactosyl polylysine (Gd-DTPA858-gal2284-PL2136) to obtain contrast-enhanced MRIs of rat liver– implanted mammary adenocarcinomas that had a decreased intensity (tumors devoid of asialoglycoprotein [ASG] receptors) when compared with the increased intensity of normal hepatic tissue. 16 The ASG receptor is the hepatocellular prototype of a cell surface lectin that reflects the differentiated state of the liver cell, and its expression has been found to be significantly reduced during malignant dedifferentiation.17,18
Alternatively, particulate contrast agents based on magnetite, monocrystalline iron oxide nanoparticles, ultrasmall superparamagnetic particles of iron oxide (USPIO), or superparamagnetic particle of iron oxide (SPIO) microspheres generate significant susceptibility changes and therefore provide strong T2 and T2* contrast enhancement.19–21 Artemov and colleagues used streptavidin-conjugated SPIO targeted to the tyrosine kinase HER2/neu receptor (found in ≈25% of human breast cancers) to visualize three established human breast cancer cell lines in vitro with varying expression levels of HER2/neu. 22 Also, USPIOs targeted to ASG receptors on rat hepatocytes by coating the iron oxide particles with arabinogalactan were previously used by Reimer and colleagues to obtain MRIs of ASG-positive nonmalignant rat liver tumors. 17
According to the 2003 World Health Organization report, liver cancer was the fifth highest cause of mortality in men owing to malignant neoplasms in 2002, accounting for 7.2% incidence and 11% mortality of all cancers worldwide, 23 signifying it as one of the most common cancers. In less developed countries around the world, over 457,000 individuals develop liver cancer, whereas in more developed countries, the number is decreased to 106,000. 23 The incidence of hepatocellular carcinoma (HCC) can be as high as 20% of the population in certain endemic regions of Africa and Asia. 24 Etiologic factors that lead to the development of HCC in humans may include chronic infections of hepatitis B (two-thirds of liver cancer cases in developing countries) and C viruses, exposure to carcinogens (eg, mycotoxins, such as aflatoxins [eg, aflatoxin-B1 (AFB1)] and cyanobacterial toxins), cirrhosis (major clinical HCC risk factor; 70 to 90% of HCCs develop in patients with macronodular cirrhosis), chronic alcoholism, and smoking. 23 In combination with hepatitis B viral infections, exposure to AFB1 is thought to be largely responsible for the high incidence of HCC in southeast China and southern Africa. 25 The incidence of HCC in the United States has increased noticeably over the past two decades and has become progressively associated with younger age groups, mainly owing to hepatitis C viral infections. 23 Human HCCs make up 90% of all primary hepatic malignancies. 24 Although the etiology of HCC varies around the world,25–27 in general, the prognosis associated with HCC is very poor. 26
Nearly all types of primary liver tumors known to occur in humans can be generated by either exposure to chemicals 27 or dietary changes in laboratory animals. 28 Carcinogenesis may occur as a result of exogenous stimuli exposure, such as aflatoxin,25,28–30 as well as by genetic 31 and epigenetic influences from endogenous factors, such as diet. 32 Continuous feeding of a diet deficient in choline is well documented to cause HCCs in rats within ≈12 to 18 months in the absence of any known exogenous carcinogens via the following mechanisms: repeated hepatocyte death and subsequent regeneration, oxidative stress, altered signal transduction, and deoxyribonucleic acid (DNA) hypomethylation. 32
In this study, we used magnetic resonance molecularly targeted imaging in a choline-deficient rat hepatocarcinogenesis model to visualize increased in vivo expression of the c-MET antigen in neoplastic nodules with the use of an SPIO-based compound tagged to a rabbit antirat antibody specific for c-MET.
Materials and Methods
The use of rats and the procedures conducted in the presented research are in accordance with the US Public Health Service Policy on Human Care and Use of Laboratory Animals, and the required approval was obtained from the Oklahoma Medical Research Foundation Institutional Animal Care and Use Committee (IACUC).
Choline-Deficient, L-Amino Acid–Defined Hepatocarcinogenesis Model
Six-week-old male Fischer rats (n = 3) were continuously fed a semisynthetic, choline-deficient, L-amino acid–defined (CDAA) diet containing practically no choline and low methionine, (Dyets, Bethlehem, PA) for up to 56 weeks.32,33 The control diet was a choline-sufficient,
Syntheses of c-MET-Specific MRI Contrast Agents
To recognize the c-MET antigen, a mouse polyclonal anti-c-MET antibody (Met [B-2], Santa Cruz Biotechnology Inc., Santa Cruz, CA) was used. To enable attachment of streptavidin–SPIO conjugates to the antibody, anti-c-MET antibodies were biotinylated as described by Artemov and colleagues. 34 Anti-c-MET antibodies were prepared in phosphate-buffered saline (PBS) (5 mg/mL), and an EZ-Link Sulfo-NHS-LC Biotinylation kit (Pierce, Rockford, IL) was used to attach N-hydroxysulfo-succinimide-biotin groups to primary amines of the antibody with a spacer arm of 22.4 Å. 34 The conjugated antibody was separated from lower molecular weight compounds with a dextran desalting column (Pierce). Dynabeads M-280 strepatividin (uniform, superparamagnetic, polystyrene beads [2.8 μm diameter] with a monolayer of streptavidin covalently attached to the hydrophobic bead surface layer) (Dynal Biotech ASA, Oslo, Norway) were used to provide specific binding to biotinylated antibodies. 34 The strong magnetic moment of SPIO molecules is the basis for magnetic cell separation, and in this technique, the SPIO microbeads are directed to cell surface receptors either directly using SPIO-conjugated specific antibodies or indirectly by attachment of SPIO to the cell surface markers prelabeled with antibody. 34
In Vitro Studies on Cells
Initial binding specificities of our molecular targeting agents to cell culture cells were done as follows. H4-II-E-C3 rat hepatoma and clone 9 rat hepatocyte cells were grown in six-well tissue culture plates. The labeling of cells was performed by the following steps. Cells (107) were fixed with 2% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.4 for 15 minutes at room temperature, washed three times (5 minutes each) in PBS buffer cells, and blocked with 1% gelatin bovine serum for 30 minutes. After aspiration of the blocking solution, the cells were incubated with primary biotinylated antibody (anti-c-MET) at a concentration of 2 μg/mL (1:100) dilution for 30 minutes at room temperature, washed three times, and incubated with SPIO (20 μg/mL; Dynabeads M-280 with streptavidin [10 mg beads/mL]; Dynal Biotech ASA) for 15 minutes. 21 After washing three times, the cells were scraped and embedded in soft agarose gel (type IX ultralow gelling temperature agarose in 3% solution in PBS buffer; Sigma, St. Loius, MO). 21
In Vitro Molecularly Targeted MRI
T2-weighted images were acquired using a two-dimensional spin-echo imaging pulse sequence (Bruker, Ettlingen, Germany). Pixel-by-pixel relaxation maps were reconstructed from a series of T2-weighted images using a nonlinear two-parameter fitting procedure. A decrease in T2 relaxation (from T2 maps) for SPIO-conjugated molecular targeting agents indicated that anti-c-MET SPIO-conjugated probes were bound to their respective target sites on cells.
Immunohistochemistry
Immunohistochemical staining of c-MET was done by incubating tissue sections with anti-c-MET antibodies conjugated to fluorescein isothiocyanate in PBS containing 0.1% v/v saponin. Immunofluorescence staining of the streptavidin-SPIO component of the molecular targeting agent SPIO-anti-c-MET was done by incubating liver cryosections with goat antirabbit-biotin (11 μg/mL in PBS-saponin (SAP) containing 1% BSA) for 2 hours at 20°C, washed 3 × 10 minutes in PBS, and incubated with donkey antigoat (final concentration 25 μg/mL) in BSA containing PBS-SAP buffer. Stained tissue slices were viewed and photographed with a Nikon C1 confocal laser scanning microscope (Nikon Instruments, USA).
In Vivo MRI
MRI experiments were carried out under general anesthesia (1–2% isoflurane and 0.8–1.0 L/min O2). The MRI equipment used was a Bruker Biospec 7.0 Tesla/30 cm horizontal-bore imaging spectrometer. Anesthetized (2% isoflurane) restrained rats were placed in an MRI probe, and their livers were localized by MRI. Images were obtained using a Bruker S116 gradient coil (2.0 mT/m/A) and a 72 mm quadrature multirung radiofrequency coil. Rats were injected intravenously with c-MET antibodies tagged with SPIO-conjugated contrast agents (5 mL/kg; 1 mg antibody/kg; 20 mg Fe/kg; during the conjugation procedure, 200 μg/mL anti-c-MET antibodies were mixed with 1.38 × 108 SPIO beads [200 μL of 10 mg/mL]).13,15,35–37 Multiple 1H-MRI slices were taken in the transverse plane using a spin-echo multislice sequence (repetition time [TR] 1 second, echo time [TE] 30 milliseconds, 256 × 256 matrix, four steps per acquisition, 6 × 5 cm2 field of view, 1 mm slice thickness). T2-weighted images were acquired using a two-dimensional spin-echo imaging pulse sequence with multiple echoes (eight echoes, 15-millisecond intervals, TE1 = 15 milliseconds; TR = 3 seconds). Streptavidin-SPIO-anti-c-MET was administered via a tail vein catheter (20–200 μL SPIO-streptavidin; 200 μg anti-c-MET antibody). Pixel-by-pixel relaxation maps were reconstructed from a series of T2-weighted images using a nonlinear two-parameter fitting procedure.

Immunohistology of (A) rat control liver cells (clone 9) and (B) rat hepatoma cells (H4IIEC3) stained (red: biotin-fluorescence complex; blue: nuclei stain) for anti-c-MET antibody (×60 original magnification).
Results
The c-MET proto-oncogene encodes a transmembrane tyrosine kinase identified as the receptor for a polypeptide known as HGF found on several cell types, mainly of epithelial origin, and overexpressed in many cancers.
Fluorescence-labeled immunoglobulins were used as immunohistologic probes for the characterization of the c-MET antigen (HGF receptor) in a control cell line (clone 9) (Figure 1A), compared with a rat hepatoma cell line (H4IIEC3) (Figure 1B). Considerably less immunohistologic staining of c-MET was observed for a control (clone 9) cell line (see Figure 1A).
An illustrative diagram of the SPIO-based molecular targeting agent is shown in Figure 2A. A SPIO-streptavidin base was used to couple a biotinylated anti-c-MET antibody. Expression of c-MET in a rat hepatoma cell line that overexpresses c-MET (as shown in Figure 1B) was established to initially determine the binding specificity of the SPIO–anti-c-MET targeting agent.

A, Illustration of the super paramagnetic iron oxide (SPIO)–streptavidin–anti-c-MET molecularly targeted magnetic resonance imaging (MRI) contrast agents, modified from Artemov and colleagues. 22 B, MRI (T2-weighted gradient echo) scan at 7.0 Tesla of clone 9 (control) cells in 3% agarose, with biotinylated anti-c-MET antibodies alone (far left column), or with antibodies plus streptavidin-SPIO (center column), or H4IIEC3 rat hepatoma cells with antibodies alone or biotinylated antibodies and streptavidin-SPIO. Decreased signal is apparent only in hepatoma cells with expression of c-MET antigens.
MRI was subsequently used to characterize alterations in T2 relaxation in cells that were treated with the SPIO–anti-c-MET targeting agent or the biotinylated anti-c-MET antibody alone and then washed prior to being put in vials in a 3% agarose solution. Figure 2B depicts representative T2-weighted images taken from vials containing rat clone 9 (control) or H4IIEC3 hepatoma cells in 3% agarose containing either biotinylated antibody alone (anti-c-MET) or antibody plus streptavidin-SPIO. T2 values for the clone 9 and H4IIEC3 cells with antibody alone were 147.6 and 131.1 milliseconds, respectively. Comparative in vitro MRI T2 value measurements of cancer and control cells in vials with both streptavidin-SPIO and anti-c-MET antibody indicated a substantial decrease in T2 for the SPIO-based agent for the H4IIEC3 rat hepatoma cells with a value of 55.8 milliseconds, but not for the control clone 9 cells, which had a T2 value of 124.1 milliseconds.
Immunofluorescence staining for overexpression of c-MET in rat neoplastic hepatocyte nodules, after being on a CDAA diet for 48 weeks, was also established as shown in Figure 3A. Comparatively, CSAA-treated rats had substantially less c-MET expression (Figure 3B) compared to immunofluorescence staining for the over-expression of c-MET in CDAA-treated rat neoplastic hepatocyte nodules, after being on a CDAA diet for 48 weeks. Western blots of the c-MET levels in CSAA and CDAA rat livers at 48 weeks confirmed overexpression in the CDAA-treated rats (Figure 3C). Morphologic MRI was used to detect hepatic nodules and tumors, as shown in representative image slices of CDAA-treated rats at 48 and 56 weeks in Figures 3, D to F. The nodules appear to be heterogeneous, whereas tumors stand out as hypointense lesions (Figure 3F). From our investigations on rats fed the CDAA diet, we have determined that neoplastic nodules develop at 48 to 52 weeks and that adenomas and small HCCs eventually develop in the livers at about 52 to 66 weeks.
Molecularly targeted MRI was used to visualize in vivo c-MET (tyrosine kinase receptor) expression in neoplastic liver nodules in choline-deficient rats at 48 weeks and liver tumors at 56 weeks (shown in Figures 4 to 6). Figures 4 to 6 depict decreases in T2-weighted MRI signals, as well as changes in T2 values and signal intensities within selected regions of interest (ROI) for some neoplastic nodules resulting from the presence of a SPIO–anti-c-MET targeting agent in a CDAA-treated rat liver, for each rat liver before (precontrast) and postcontrast after administration of the SPIO–anti-c-MET targeting agent. One of the week 48 CDAA-treated rats was administered one-tenth the concentration of SPIO, compared with an additional week 48 CDAA-treated rat and the week 56 CDAA-treated rat, to determine if lower concentrations of SPIO were sufficient for detection by MRI.
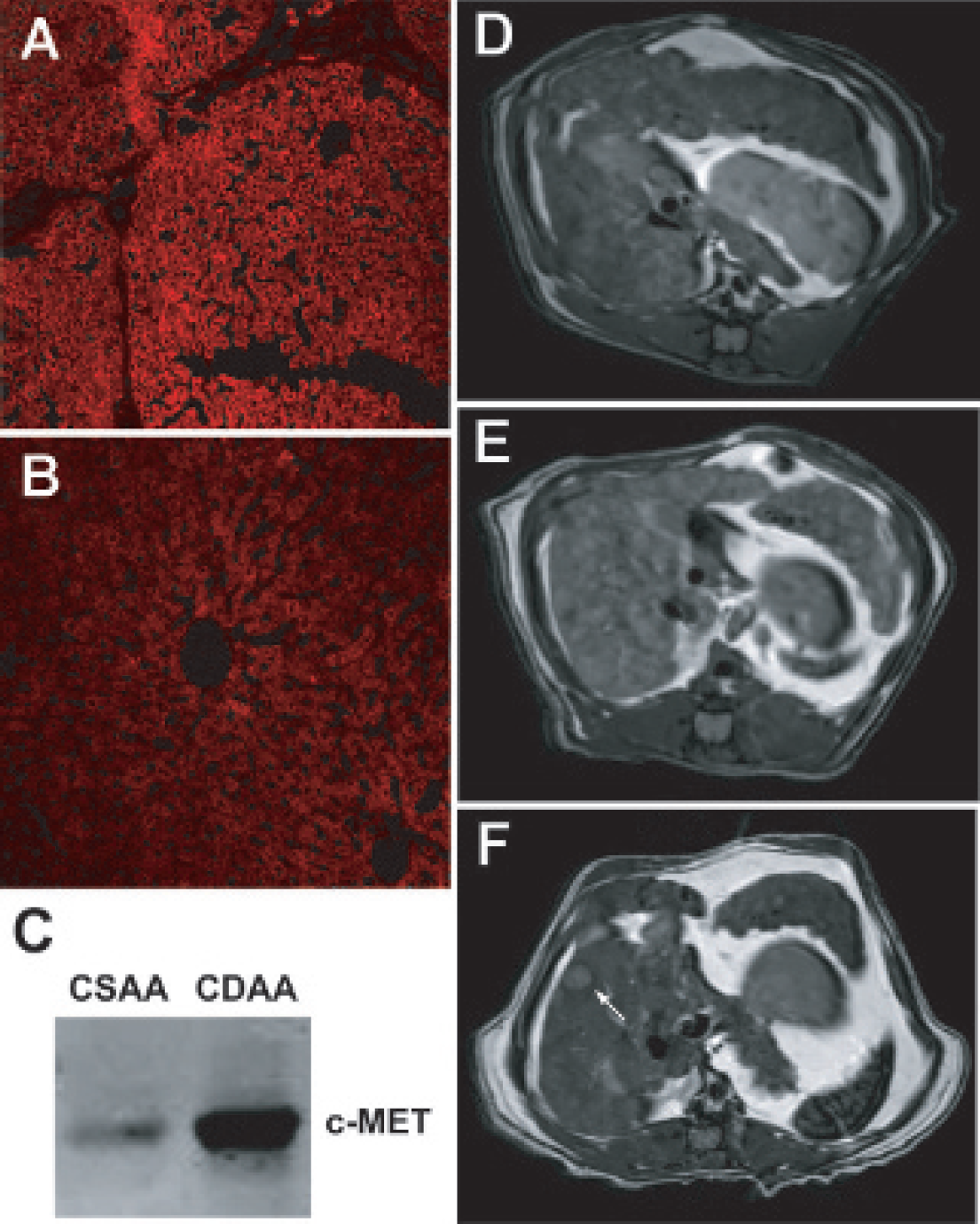
Immunohistology of (A) nodular choline-deficient
Two negative controls were also incorporated into the study, including a week 56 CSAA-treated rat administered with the SPIO-anti-c-MET targeting agent and a week 48 CDAA-treated rat administered SPIO alone (without anti-c-MET antibody), as shown in Figure 7. T2 and MRI signal intensities for the CSAA rat were found to not substantially change for two of the selected ROI and were found to both slightly decrease for ROI 1 (see Figure 7, A–C). The CDAA-negative control administered the SPIO alone showed no substantial changes in both T2 and MRI signal intensities (see Figure 7, D–F).
We also obtained immunofluorescence confirmation of the binding specificity of the SPIO–anti-c-MET targeting agent to the hepatocytes following MRI studies, as shown in Figure 8. The streptavidin-SPIO (no anti-c-MET antibody) control (see Figure 8C) showed no staining for streptavidin, indicating that streptavidin-SPIO without the anti-c-MET antibody does not bind nonspecifically to hepatocytes. The CSAA-negative control administered the SPIO–anti-c-MET targeting agent had some of the targeting agent bind to hepatocytes (see Figure 8D), which correlates with some decrease in T2 and MRI signal intensity in Figure 7, B and C, respectively.

A, T2-weighted magnetic resonance images of a choline-deficient
Discussion
Invasive growth is controlled by a variety of cytokines and growth factors, such as epidermal growth factor, insulinlike growth factor 1, fibroblast growth factor, and the involvement of a family of soluble growth factors known as scatter factors. 38 The prototype of all scatter factors is the HGF. 38 The receptor for HGF is a transmembrane protein with tyrosine kinase activity encoded by the MET proto-oncogene. 38 MET is produced as a single polypeptide chain that is cleaved during biosynthesis into two subunits, an a subunit (50 kDa) and a β subunit (145 kDa), bridged by a disulfide bond. 39 The C-terminal of the β chain consists of an extracellular sequence, a transmembrane domain, and a cytoplasmic protein kinase domain, whereas the N-terminal a chain is only extracellular. 38 The SPIO–anti-c-MET antibody complex that we used in this study would bind to the extracellular regions of Met, as confirmed by fluorescence labeling of the streptavidin component of the targeting agent attached to hepatocyte cell membranes overexpressing Met (see Figure 7).
The multifunctional growth factor scatter factor–HGF and its tyrosine kinase receptor, c-MET, have been implicated in the genesis and malignant progression of multiple human malignancies. 40 c-MET is crucially involved in invasive cell growth and motility during embryogenesis and has been detected in invasive tumors, including HCCs. 3 It is thought that inappropriate activation of HGF-dependent pathways leads to a malignant process via the weakening of tissue constraints by tumor cells, migration, invasion of foreign regions, and subsequent metastasis. 40
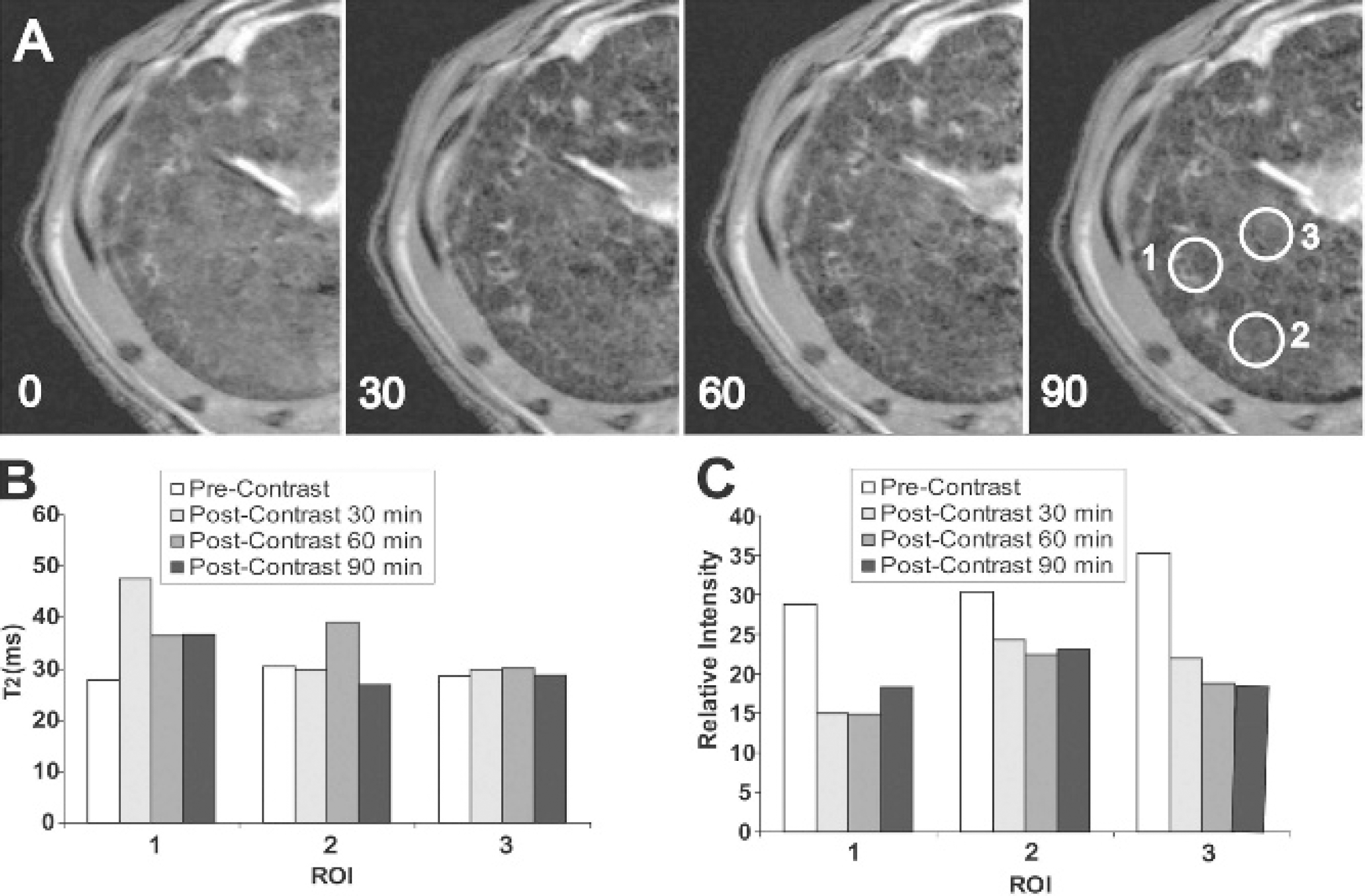
A, T2-weighted magnetic resonance images of a choline-deficient
It is also known that Met is directly associated with regulation of apoptosis. A model for Met-induced Fas sequestration, as studied by Wang and colleauges, indicates that Met promotes cell survival by binding to Fas to prevent Fas from interacting with its ligand (FasL) and to prevent Fas from self-aggregating or clustering. 41 Thus, in the presence of low levels of FasL or no FasL, Fas and Met remain associated, and activation of the apoptosis pathway via programmed death receptor activation is inhibited. 41 Thus, a high Met to Fas ratio elicits cell survival (as is present in many cancer cells). 41 Huh and colleagues found that mice lacking the c-MET gene in hepatocytes were hypersensitive to Fas-induced apoptosis. 42
This is the first attempt to use MRI molecular targeting as an approach to detect in vivo expression of c-MET in neoplastic lesions. A study by Shaharabany and colleagues had previously used the functional MRI method BOLD (blood oxygenation level dependent) contrast MRI at 1.9 Tesla to measure hemodynamic alterations in HGF/scatter factor (SF) in Met-expressing organs and tissues in tumor-bearing (orthotopically implanted D1-DMBA-3 [DA3, mammary adenocarcinoma induced by dimethylbenzan-thracene]| cells in the mammary gland; 3 weeks following injection) BALB/C mice. 43 In this study, HGF/SF was injected (intravenous in saline) at doses of > 130 ng/g to obtain alterations in BOLD signal contrast. 43 It was previously found that in breast cancer, Met expression is typically found to be higher at the tumor margins 44 and that their method may enhance tumor edge detection. 43 Hay and colleagues used scintography to detect over-expression of Met in mice bearing tumors, either autocrine for human HGF/SF and Met [S-114 (NIH 3T3 cells transformed with human HGF/SF and Met), or SK-LMS-1 (human leiomyosarcoma cell line autocrine for human Met and HGF/SF)], or autocrine-paracrine for murine HGF/SF and Met [M-114 (NIH 3T3 cells transformed with murine HGF/SF and Met), or DA3 cells] with an 125I-labeled monoclonal antibody for the extracellular domain of Met. 45

A, T2-weighted magnetic resonance images of choline-deficient
In our study, the SPIO-based anti-c-MET targeting agent was found to specifically bind to neoplastic hepatocyte nodules (in Figure 7), which were detected by MRI with a characteristic decreases in T2 relaxation values and MRI signal intensities (see Figures 4–6) in specified ROI (see Figures 4A, 5A, and 6A). We were also able to differentiate between neoplastic nodules that overexpressed c-MET owing to a decrease in both T2 values and MRI signal intensities from those nodules that had no overexpression of c-MET owing to no substantial change in T2 values following administration of the anti-c-MET targeting agent. For example, the 48-week rat depicted in Figure 4, which was given a lower concentration of the SPIO-streptavidin compound (but the same concentration of antibody as other treated animals), showed a signal decrease in intensity for all three selected ROI, but there was only a decrease in T2 for ROI 1. For the second week 48 CDAA-treated rat (depicted in Figure 5, given a higher concentration of SPIO-stretavidin mixed with anti-c-MET antibody), there was also an MRI signal decrease for all three ROI, but only a slight decrease in T2 for ROI 2. These results may reflect the fact that few nodules overexpressed c-MET at 48 weeks. For the week 56 CDAA-treated rat (see Figure 6), both MRI signal intensities and T2 values substantially decreased for ROI 1 and 2, with little change in both parameters for ROI 3. The longer treatment on the CDAA diet suggests the development of neoplastic nodules with increased expression of c-MET. The SPIO–anti-c-MET agent seems to bind specifically to the hepatocytes in the immunofluorescence histology slides (see Figure 8A), with a few found in the sinusoids that may be due to uptake by Kupffer's cells. The CSAA-negative control also showed some expression of c-MET, which represents ambient levels of c-MET expression in normal liver. The MRI results and the confirmation by immunofluorescence (see Figure 8) of specific c-MET targeting by the SPIO–anti-c-MET agent clearly demonstrate that the targeting agent is specific for c-MET on the cell surface of hepatocytes that overexpressed the antigen. It is feasible that this method can be used for in vivo visualization of nodules or tumors that highly express the c-MET antigen and therefore may predict early detection of lesions that have a high malignant potential.
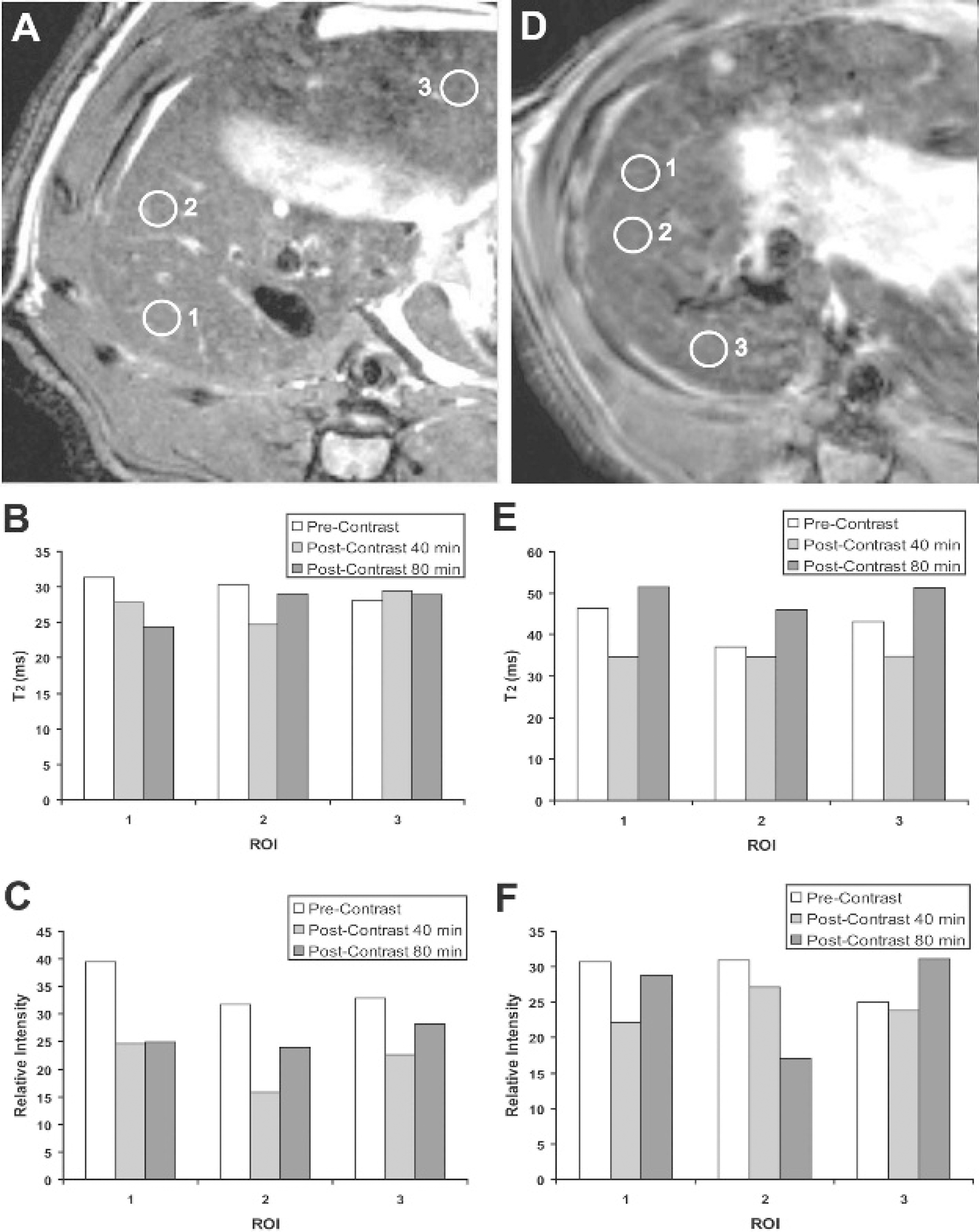
A, T2-weighted magnetic resonance image (MRI) of a choline-sufficient,

Representative immunofluorescence images of fresh-frozen liver sections from (A) a choline-deficient