
Abstract
Ages at menarche and menopause have been shown to be associated with adverse health outcomes in later life. For example, earlier menarche and later menopause have been independently linked to higher risk of breast cancer. Earlier menarche may also be associated with an increased risk of endometrial cancer, menstrual problems and adult obesity. Given the associations of ages at menarche and menopause with future health outcomes, it is important to establish what factors across life, and generations, may influence these. This article examines the associations of early life factors, namely birthweight, bodyweight and growth during childhood, childhood socioeconomic circumstances and psychosocial factors with ages at menarche and menopause. It examines possible explanations of the associations found, including life history theory, and discusses areas for future research.
Reproductive health, from menarche to menopause, is not only understood as being integral to women's overall health and wellbeing, but is increasingly recognized as a sentinel of chronic disease in later life [1–3]. For example, both earlier menarche and later menopause are independently associated with higher risk of breast cancer [4,5], and may also be associated with an increased risk of endometrial cancer [6–8]. When examined together, the length of time between menarche and menopause, which provides a crude indicator of lifetime estrogen exposure, has also been shown to be associated with these outcomes [9]. Earlier menarche may also be a risk factor for adult obesity [10]. By contrast, age at menarche has little bearing on cardiovascular risk, and only premature age at menopause is associated with significantly higher risk of heart disease [1,11].
There is growing evidence that the physical and social environment in previous generations, and from preconception to midlife, influence both reproductive health [1] and, later, chronic disease [12,13]. Menarche and menopause are the two major components of women's reproductive lives, since the interval between them determines the natural reproductive phase [14]. Therefore, the aim of this article is to update information on the role that early life factors play on these key aspects of reproductive health, namely age at menarche and the timing of menopause.
Methods
A comprehensive search of the literature published in the English language between 1980 and July 2008 was performed using the terms ‘childhood, early life, fetal, in utero or epigenetic’ and ‘age at menarche, age at menopause or hysterectomy’. Of the studies identified, we have chosen to include those with a sample size of at least 400, as it has been suggested that these will have sufficient power to study menopause [15]. We assumed this sample size was also sufficient for the study of menarche since this characteristic (excluding precocious menarche) is less variable than age at menopause [16].
Age of menarche
Menarche marks the commencement of the reproductive phase of a woman's life. Records suggest that there was a secular decline in the average age at menarche in developed countries across the 19th and 20th Centuries until the 1950s [17], with an average age at menarche of 15–17 years reported in the mid-19th Century [18,19]. It has been suggested that pubertal timing may then have plateaued in the 1950s, although further small declines have subsequently been reported [17]. While recent estimates of the median age of menarche vary between 13 to 16 years, recent data suggest that the age of 15 years represents the 95–98th percentile for menarche [17]. A review of family and twin studies has highlighted the key role of genetic factors in determining the timing of menarche, with heritability estimates ranging from 0.44 to 0.72 [20,21]. In examining early life predictors of age at menarche, three major themes have emerged, namely the effects of body size, social circumstances and exposure to unfavorable psychological circumstances. The effects of these environmental influences on the timing of menarche can be explained, from an evolutionary-development perspective, in terms of life history theories.
Life history theory perspectives on the timing of menarche
The main life history theories have been elaborated by Ellis [22] as the energetics theory, and the four psychosocial models of pubertal timing: psychosocial acceleration, parental investment, stress suppression and child development theory. However, the theories differ in their conceptualization of the nature, extent and direction of environmental influences on the age of menarche, and the effect that the timing of menarche has on subsequent reproductive characteristics [22].
The energetics theory suggests that energy availability during childhood influences the timing of menarche. It hypothesizes that girls who were exposed to a chronically poor nutritional environment will grow more slowly, experience later pubertal development (relative to their genetic potential), and reach relatively small adult size compared with those children who were exposed to greater food availability. This is consistent with an earlier idea regarding the impact of relative fatness that follows from the close relationship of average critical body-weight on age at menarche [23]. The psychosocial acceleration theory posits that the experience of high levels of emotional stress in and around a girl's family leads to earlier menarche in order that she can maximize her chance of leaving descendents [24,25]. Based on the same logic, the parental-investment theory hypothesizes a special role for the father and other men in influencing the timing of menarche. The stress-suppression theory [26] proposes that early adversity, whether it is by adverse physical or social conditions or psychosocial stress, causes a delay in pubertal development until better times. Last, Ellis describes child-development theory as reconceptualizing ‘the age at menarche as the end point of a developmental strategy that conditionally alters the length of childhood in response to the composition and quality of family environments’ [22].
Birthweight & childhood growth
Several studies have examined birth size and infant growth in relation to age at menarche
Early life factors and their associations with age at menarche.

Other studies have found that rapid prepubertal weight gain and childhood obesity are associated with an earlier menarche [33]. Using the growth status at birth as denoted by the expected birthweight ratio (EBW; a ratio of observed birthweight over median birthweight appropriate for the maternal age, weight, height, infant sex and gestational age), an Australian study found that earlier menarche was predicted by lower EBW combined with higher BMI during childhood [34]. Findings from the UK Newcastle One Thousand Families study support the notion of an interaction between birth-weight and weight much later in childhood; specifically, girls who were large for gestational age and heavy at the age of 9 years had the earliest menarche [35]. From a life history perspective, results from the above studies appear to support the energetics theory for timing of menarche. While improvements in childhood nutritional status over time are thought to underlie the secular declines in age at menarche, the precise role of dietary composition in childhood remains unclear [36]. It is also worth noting that a recent study found no evidence to support a link between the secular rise in the prevalence of childhood obesity and the decline in average age at menarche [37].
Childhood socioeconomic position
Numerous studies have investigated the relationship between childhood socioeconomic characteristics (e.g., urbanization, education, parental occupational class and family size) and age at menarche
Since socioeconomic circumstances often act as proxies for many factors, including the quality and quantity of food intake, energy expenditure, family structure and access to healthcare, few studies are able to separate out the effects of specific factors underlying the observed relationships between socioeconomic circumstances and timing of menarche. For instance, socioeconomic differences established in early childhood may be related to maternal efficiency during pregnancy (i.e., the ability of the mother to cope with and care for her unborn child), the duration of breastfeeding and the quality of the diet immediately after being weaned [48]. Associations between socioeconomic conditions and menarche may also be mediated by childhood growth; however, few studies have tested this hypothesis. Again, as socioeconomic circumstances influence nutritional environment in childhood, its relationship with the timing of menarche is consistent with the energetics theory of life history.
Psychosocial factors: childhood experiences
Role of family structure & relationships
While a mother's age at menarche appears to be a better predictor of the daughter's age at menarche than other external factors [52], research supports the role of family structure in determining pubertal timing. Paternal affection, positive family relationships and paternal involvement in child rearing are related to a comparatively later age of menarche [53,54], while increased family conflict, divorce and longer durations of paternal absence are correlated with earlier menarche [55–57]. It is also found that stepfathering predicts menarcheal age better than absence of a biological father and a longer presence of a stepfather correlates with earlier ages of menarche [58].
Exposure to external stressors/trauma
Results from the 1934–1944 Helsinki Birth Cohort have found that the 396 evacuees who were sent by their parents, unaccompanied, to temporary foster families in Sweden and Denmark because of the Soviet–Finnish wars, had earlier menarche than other girls [59]. Similarly, other studies examining the effect of migration have found increased occurrence of early sexual development, indicated through precocious puberty, in adopted children in cohort studies from European countries [60]. For example, a recent Danish study found that internationally adopted children were more likely (∼15–20-times) to develop precocious puberty compared with the Danish reference group [61]. In addition, children migrating with their families had no increased risk of precocious puberty [61]. Exposure to other types of stressors in childhood, such as sexual abuse, have also been shown to be associated with the early onset of menarche [57,62]. These results concur with the studies associating childhood adversities, lack of paternal investment and father's absence with earlier onset of menarche [22,53,63].
Studies have also found that age at menarche is delayed during periods of war. Authors working on the 1944–1945 Dutch Famine Study attributed this to the effect of food rationing [64]. In another study, it was found that girls living in besieged Srebrenica during the Balkan War had delayed age at menarche [65]. However, the author concluded that psychological trauma, physical injury and low socioeconomic conditions, as a result of war, may have been the causes. A study of girls living in the city of Šibenik, Croatia where the menarcheal status of girls was surveyed three-times in 1981, 1985 and 1996, found that there was a significant increase in mean menarcheal age of approximately 3 months between 1985 and 1996. This increase over time was dependent on the nature of the stress experienced with those girls who had experienced personal tragedies during the Balkan war, demonstrating an even greater delay in menarche of almost 11 months [50].
Research based on observational studies has also demonstrated delayed pubertal development following childhood adversity [50,64]. The effects of adversity may vary depending on the timing of exposure to the stressors [59]. For example, it has been shown that exposure to stress in early childhood is associated with earlier menarche [22], whereas exposure during or shortly before puberty has been associated with later onset of menarche [50,64].
Each of the three psychosocial models of pubertal timing within life history theory, namely psychosocial acceleration theory, parental investment theory and stress-suppression theory, can be invoked to explain the differing effects on age at menarche of each adverse condition in childhood identified previously. As an alternative, Boyce and Ellis have suggested the stress reactivity theory to account for both the delaying and accelerating effects of psychosocial stress on the timing of menarche [66]. They propose the notion that both highly protective and acutely stressful childhood environments trigger stress reactivity systems. If this triggering inhibits maturation of the hypothalamic–pituitary–gonodal axis, then this should produce a U-shaped relationship with age at menarche, whereby high social resources and support levels as well as high psychosocial stress and adversity are both correlated with later timing of menarche.
Age of menopause
Menopause marks the end of the reproductive phase of a woman's life and usually occurs between the ages of 40 and 60 years [67–69], with the most reliable estimates indicating that the median age at menopause in Western industrialized countries is between 48 and 52 years [70]. While secular changes in average age at natural menopause are not as well documented as those in average age at menarche, there is some evidence to suggest that there have been modest increases in average age at menopause over the course of the 20th Century [71–74]. The impact of menopause is felt in physical, psychological and sociocultural domains and continues into the postmenopausal years.
Natural menopause is reached when a woman's follicular reserve is depleted to approximately 1000 follicles, from a peak during fetal life of approximately 5 million at 20 gestational weeks, which has declined to approximately 2 million by birth [75,76]. The most important determinants of age at natural menopause are considered to be factors that affect the duration of decline in ovarian follicle reserve:
Number of primordial germ cells that migrate to the gonadal ridge during intrauterine life;
Mitotic abilities of these cells until gestational age of approximately 16–20 weeks;
Rate of follicular atresia in both intra- and extra-uterine environments [76,77], suggesting that early life factors may play an important role in influencing age at menopause.
Throughout their reproductive lives, the general health and quality of lives of millions of women are affected by gynecological problems. Before reaching natural menopause, a significant proportion of women undergo treatment for such problems. The most radical of these treatments, namely hysterectomy and/or bilateral oophorectomy, end reproductive life. In these cases, women are said to have undergone surgical rather than natural menopause. Hysterectomy rates vary greatly between countries [78]. Our research from the MRC NSHD, a British cohort representative of women born in the 1940s, found that 22% of women had undergone hysterectomy by the age of 57 years with a mean age at hysterectomy of 43.6 years (range: 27–56 years). In this cohort, the most common reasons for undergoing hysterectomy were fibroids (30.5%) and menstrual problems (28.8%). Rates of hysterectomy in some other countries, including the USA and Australia, are higher than those reported in the UK [79,80]. For example, prevalence of hysterectomy was estimated to be between 33 and 43% among women aged 45–59 years in a recent study investigating women in the USA [81]. Despite these differences in prevalence between countries, reasons for performing hysterectomy are similar – US national health statistics suggest that between 1988 and 1993, fibroids were indicated in more than a third of hysterectomy cases [82]. As the processes that lead to the development of gynecological problems and that result in women seeking and undergoing treatment for such conditions are different to the processes that determine natural age at menopause, it may be expected that early life predictors of surgical menopause may differ from those of natural menopause.
Family and twin studies have indicated that the genetic effect on the timing of natural menopause is considerable, with estimates of heritability ranging from 30 to 85% [68,83]. Supporting this is evidence from cross-sectional and cohort studies that have demonstrated that a woman's age at natural menopause is strongly associated with her mother's reported age at menopause [68,84–89]. Studies also suggest that there is a genetic component to hysterectomy (i.e., surgical menopause) risk [90,91]. While the relationships between many adult environmental factors and timing of natural menopause have been investigated, only a few have consistently been related to an earlier menopause. Those that have are cigarette smoking [67,70,88] and nulliparity [90–94]. Some studies have also found a relationship between adult socioeconomic position and the timing of natural menopause with those women of lower position experiencing earlier menopause than women of higher position, even after adjustment for potential confounding factors, such as smoking and parity [95–98]. A range of factors in adulthood, including higher parity, lower socioeconomic position and greater changes in bodyweight over time, have been shown to be related to increased risk of hysterectomy [99–108].
As for age at menarche, a number of studies have examined the association between early life factors and age at natural menopause
Early life factors and their associations with age at natural menopause.
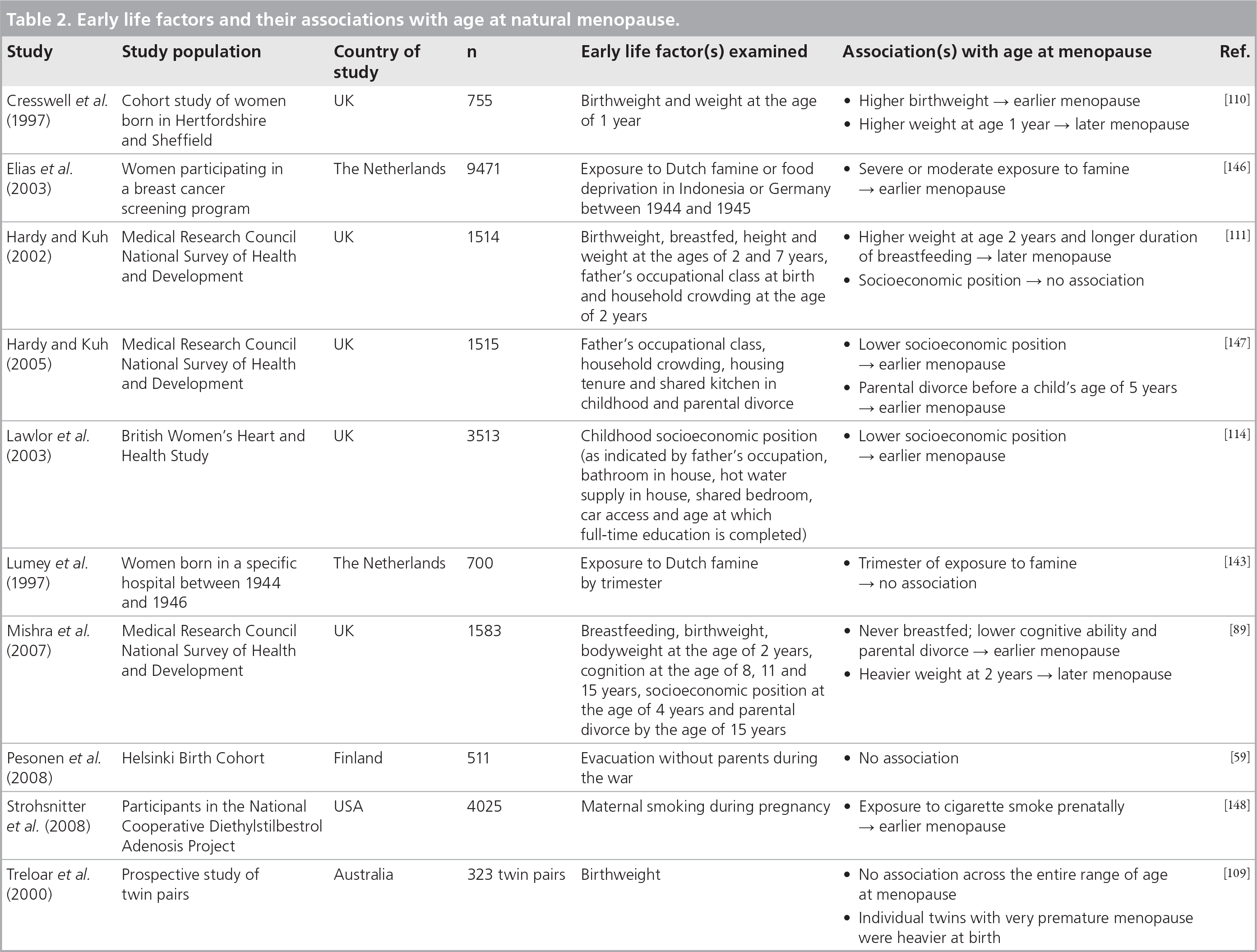
Birthweight, childhood growth & early life nutrition
A number of epidemiological studies have investigated the influence of birthweight, childhood growth and early life nutrition on age at natural menopause. A twin study in Australia [109] and a cohort study of women in Hertfordshire, England [110], found no evidence that higher birthweight was associated with later menopause. However, low weight at 1 year was found to be associated with earlier menopause in the Hertfordshire cohort [110]. From the MRC NSHD study, it was found that women who had been breastfed had later menopause than other women [89,111]. Further evidence that early nutrition or growth might be important comes from a study of women in New Guinea [112], where the median age of menopause in a population who had suffered severe and prolonged malnourishment, and who were consequently of short height and low weight, was estimated to be 4 years earlier than women in the same region with better nourishment. This suggests that malnourishment (possibly acting prenatally through maternal undernutrition or postnatally through poor childhood growth) may play a role in early menopause.
A study of women exposed to the Dutch famine of 1944–1945 provides further support for the role of postnatal nutrition in influencing age at natural menopause – women who had been exposed to severe caloric restriction, especially those who were aged 2–6 years at the time of exposure, had an earlier natural menopause than those who were not exposed to these restrictions [113]. Famine exposure was also related to a higher occurrence of hysterectomy [113], with those women who were exposed to severe famine at increased risk of hysterectomy compared with those who were not (HR: 1.53; 95% CI: 1.27–1.84) [113]. The associations of birth size, childhood growth and early life nutrition with hysterectomy are less well researched. In the MRC NSHD where the association of weight in early childhood with hysterectomy was examined there was no evidence that weight in childhood was associated with subsequent hysterectomy risk [107].
Childhood socioeconomic position
In the MRC NSHD, early life socioeconomic position has been found to be more strongly associated with age at natural menopause than adult position. In another study of over 3000 British women, adverse socioeconomic circumstances in childhood, as well as in adulthood, were associated with age at natural menopause. In this study, it was found that the association between childhood deprivation and early menopause may be, at least in part, mediated via childhood diet, with this affecting both linear growth and age at menopause [114]. An association between childhood socioeconomic position and hysterectomy has also been found in the MRC NSHD with those women living in poorer socioeconomic conditions and with fathers of lower occupational classes at greater risk of hysterectomy than women living in better socioeconomic conditions and with fathers of higher occupational classes [106,115]. When similar associations were tested in two other British cohorts, it was found that the associations were acting in the opposite direction among women born in the 1920s [106]. This suggests that these are dynamic associations that change over time, possibly as a result of changes over time in a range of other factors, including access to medical care, attitudes of doctors, childbearing patterns and the availability of alternative treatments. Associations between childhood socioeconomic position and hysterectomy have not always been consistently found – in an American study no evidence of an association between various indicators of socioeconomic position in early life and hysterectomy risk was found among women enrolled in the Wisconsin Longitudinal Study [116].
Parental divorce during childhood
In previous analyses of the MRC NSHD, it was found that women who experienced parental divorce early in life had an earlier natural menopause than other women, raising the possibility that early emotional stress may also be a contributing factor [2,111]. Parental divorce before the age of 15 years not only exerts a strong influence on bringing forward natural menopause, but its effect is more than doubled for women with menopause before the age of 50 years compared with other women. It may be acting as a marker for early emotional stress and, in earlier findings, has been related to a range of psychological disturbances in childhood, such as bedwetting and delinquency [117,118], as well as poorer psychological health in adulthood [119]. The impact of stress responses both in early and later life, including responses of the hypothalamic–pituitary–adrenal axis, may influence age at menopause [120–123]. The life history theory of psychosocial acceleration as described with respect to the age at menarche may also be at play here. Another potential pathway that may be operating and, hence, explain these findings is the influence of psychological stress on the ovaries by lowering telomerase activity, a cellular enzyme that may act as a marker of ovarian functional age through its association with follicular atresia [124,125].
Interactions between early life factors & age at menopause
More recently, analyses of the MRC NSHD have revealed that some early life factors have a differential effect on age at natural menopause, which is dependent on whether women reach menopause before or after the age of 50 years [89]. The evidence indicates that weight at 2 years, mother's reported age at menopause and parental divorce are more strongly associated with age at menopause if women have reached menopause before the age of 50 years compared with women who have reached menopause after the age of 50 years. In this study, a mother's reported age at natural menopause may be considered as a simple proxy for the heritability of age at menopause, which in turn reflects genetic influences on the overall size of the follicle store before and shortly after birth and the rate of follicle atresia.
The differential effect according to age at menopause may be explained if the stress responses interact with another factor, for instance, the genetic setting of the size of the initial follicular reserve. Similarly, the time-dependent influence of weight during childhood on timing of natural menopause may indicate a postnatal interaction with factors that set ovarian function and development. On the other hand, the effects may still be caused by changes in environmental factors. For instance, weight during childhood may be less strongly related to stress than previously, due to the greater availability of food and lower levels of physical activity in current compared with previous generations. We are not aware of any studies that have examined the associations between parental divorce, psychological stress/trauma and hysterectomy.
Relationship between age at menarche & age at menopause
There is conflicting evidence concerning the relationship between ages at menarche and menopause. Some studies have found a relationship between earlier menarche and earlier natural menopause or perimenopause [92,94,126,127], a few have reported a relationship between earlier menarche and later menopause [128,129], but most others have found no association [69,130–133].
Conclusion & future perspective
Thomas et al. suggested that age at menarche is mainly determined by extrinsic factors, such as living conditions, while age at menopause appears to be influenced by intrinsic factors, such as reproductive history [14]. In this article, we have found consistent evidence to suggest that ages at menarche and menopause are both affected by a woman's exposure to early life factors. The main early life factors associated with earlier menarche are:
Higher growth rate during childhood
Higher childhood socioeconomic position
Family conflict and parental divorce
Presence of stepfather
Exposure to stressors during or shortly before menarcheal age
Those associated with earlier menopause are:
Not having been breastfed
Poor early growth
Poor socioeconomic conditions
Parental divorce
It is worth noting that while some early life factors – such as growth rate, childhood socioeconomic conditions and parental divorce – affect both ages at menarche and menopause, the nature of these relationships differs. For instance, while parental divorce appears to bring about an earlier onset of both menarche and menopause, poor childhood growth reduces reproductive lifespan by delaying menarche and decreasing the age of menopause. Other risk factors also seemed to be unique to the age of menarche, such as the presence of stepfather and exposure to stressors during or shortly before menarcheal age.
Relatively little work has been done to investigate the extent to which childhood diet underlies the relationships between childhood growth and age at menarche or menopause, and further work is required. Future work to disentangle the effects of the different types of stressors in early life on pubertal timing is also required. The full impact of the rise in childhood obesity in the last few decades on the variability in ages at menarche and menopause is as yet unknown. More work is also required before any conclusions can be made about the associations of early life factors with risk of surgical menopause (i.e., hysterectomy) as these have been much less well studied than natural menopause. Furthermore, many of the results we have described need to be replicated in other studies before definitive conclusions about the early life predictors of menarche and menopause can be drawn. In this article, we have focused on menarche and menopause. However, to gain a full understanding of the associations between early life factors and reproductive health, there is also a need to consider the links of early life factors with other markers of reproductive health, such as fertility and gynecological disorders. These reproductive outcomes and their associations with early life factors have yet to be studied extensively and further research is required.
Beyond early life: continuity of reproductive health across the life course
Evidence of continuity in reproductive health across the life course indicates that for some women, the burden of poor reproductive health may be lifelong. For example, women who have early menarche are more likely to have menstrual problems [134]; those with menstrual problems are more likely to experience gynecological problems [135] and subfertility, which have been linked to earlier menopause [136]. A number of biological and social pathways may be operating across the life course that account for this continuity. These include variation in lifelong exposure or susceptibility to estrogen or other hormonal mechanisms, personality and psychological susceptibility and continuity in the socioeconomic environment. Each of these may influence and be influenced by body composition and diet. These findings suggest that the study of reproductive health would benefit from an integrated approach covering the whole life course, rather than each reproductive health outcome being studied in isolation.
From heritability studies, it has been shown that genetic factors play a key role in determining ages at menarche and menopause [20,21,68,83]. However, relatively little work has been done to unravel the complex interplay of genetic, social and behavioral factors (via epigenetic mechanisms), in explaining the variability in ages at menarche and menopause. One such approach would be to use family data and specific family-based studies (intergenerational, sibling and twin studies) across the life course to test specific causal mechanisms and life-course models since they can help in the understanding of whether the timing of risk factors (critical and sensitive periods) are important [137]. This analysis could be done in several ways, for instance, by comparing within and between sibling and twin associations for age at menarche where tight control for fixed factors within twin pairs and sibling groups is possible, and by comparing parental–offspring association to test causality for intrauterine exposure.
It is important to go beyond simple associations to understanding whether or not timing matters with respect to interventions to improve population health. For example, if an exposure during a developmental period in early life is causally related to age at menopause, but only through strong tracking of that exposure across the life course (e.g., bodyweight), then one would need to consider when it is easiest to most effectively alter the exposures. If it is easier to alter it in adulthood, then interventions in adulthood would be most appropriate [138]. With numerous family-based studies now underway, opportunities will arise to undertake such research in the future.
Executive summary
Ages at menarche and menopause have been shown to be associated with adverse health outcomes in later life.
Evidence of continuity in reproductive health across the life course indicates that for some women the burden of poor reproductive health may be lifelong, for example, women who have an earlier menarche are more likely to have menstrual problems and subfertility, which are linked to earlier menopause.
The length of time between menarche and menopause determines the natural reproductive phase in a woman's life.
Main early life factors associated with earlier menarche include:
Higher growth rate during childhood
Higher childhood socioeconomic position
Family conflict and parental divorce
Presence of stepfather
Exposure to stressors during or shortly before menarcheal age
Main early life factors associated with earlier natural menopause include:
Not having been breastfed
Poor early growth
Poor cognitive ability
Poor socioeconomic conditions
Parental divorce
Research on the impact of childhood diet and the rise of childhood obesity on ages at menarche and menopause is currently lacking.
More research is required on the effects of early life factors on the risk of surgical menopause.
There is a need for further consideration of associations between early life factors and other markers of reproductive health, such as fertility and gynecological disorders.
The study of reproductive health would benefit from an integrated approach covering the whole life course.
Collecting family data or conducting a family-based study could help in testing causal mechanisms and in timing interventions.
Footnotes
The Medical Research Council provided financial support for Gita Mishra, Rachel Cooper and Diana Kuh. Supported in part by the Intramural Research Program, National Institute on Aging, NIH. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
No writing assistance was utilized in the production of this manuscript.