
Abstract
Imidacloprid-induced hormesis in the form of stimulated reproduction has previously been reported in green peach aphid, Myzus persicae. Changes in gene expression accompanying this hormetic response have not been previously investigated. In this study, expression of stress response (Hsp60), dispersal (OSD, TOL and ANT), and developmental (FPPS I) genes were examined for two generations during imidacloprid-induced reproductive stimulation in M. persicae. Global DNA methylation was also measured to test the hypothesis that changes in gene expression are heritable. At hormetic concentrations, down-regulation of Hsp60 was followed by up-regulation of this gene in the subsequent generation. Likewise, expression of dispersal-related genes and FPPS I varied with concentration, life stage, and generation. These results indicate that reproductive hormesis in M. persicae is accompanied by a complex transgenerational pattern of up- and down-regulation of genes that likely reflects trade-offs in gene expression and related physiological processes during the phenotypic dose-response. Moreover, DNA methylation in second generation M. persicae occurred at higher doses than in first-generation aphids, suggesting that heritable adaptability to low doses of the stressor might have occurred.
INTRODUCTION
Hormesis is low-dose stimulation coupled with high dose inhibition of a stressor-induced response (Calabrese and Baldwin 2002). Chemical-induced hormesis is widespread across many biological groups and has been studied in medical, evolutionary, and ecological contexts (Calabrese and Baldwin 2003). The biochemical and molecular underpinnings of hormetic responses have been examined using several biological models (Calabrese and Baldwin 2003). Regulation of gene expression is implicit in the hormetic responses (Son et al. 2010), activating adaptive cellular stress response pathways assisted by transcriptional factors such as antioxidant response element (ARE), forkhead box O (FoxO), heat-shock factor (HSF), and nuclear factor-KB (NF-KB).
Gene expression during hormesis has not been well studied in insect-insecticide models, although changes in certain biochemical endpoints have been reported during pesticide-induced stimulations (Cutler 2013; Guedes and Cutler 2013). Stimulated production of total calcium and proteins was seen in Choristoneura fumiferana (Clemens) exposed to sub-lethal doses of organophosphorus and carbamate insecticides (Smirnoff 1983), suggesting that basic metabolites such as sugars, lipids, and total proteins can be measured in addition to organismal and population endpoints to give clues into how insect metabolism changes during hormesis. Larval weight gain and changes in isoesterase profiles were seen in Tribolium castaneum L. exposed to low concentrations of azadirachtin (Mukherjee et al. 1993), and juvenile hormone (JH) III titers were correlated with stimulated fecundity in green peach aphid, Myzus persicae (Sulzer), when exposed to low doses of imidacloprid (Yu et al. 2010). Transcriptional responses of Helicoverpa armigera Hübner to gossypol-induced hormesis were suggested to be a specific transcriptional adaptation rather than general stress response (de la Paz Celorio-Mancera et al. 2011).
The HSF pathway can be triggered by a variety of stressors, including elevated temperatures, heavy metals, viral or microbial infection, and toxins (Son et al. 2010). In agriculture, insecticides are used to kill insect pests but can activate the HSF pathway in insects that survive insecticide exposure (Yoshimi et al. 2002). Other abiotic stressors that disrupt homeostasis can also initiate the HSF pathway. For example, Chironomus tentans Fabricius (Karouna-Renier and Zehr 1999), T. castaneum L. (Mahroof et al. 2005), Cydia pomonella (L.) (Yin et al. 2006), and Liriomyza huidobrensis Blanchard (Huang et al. 2007) accumulated heat shock proteins (Hsp) following heat stress, resulting in thermotolerance. Nutritional stress with heat stress induced higher transcription of Hsp70 than heat stress alone (Salvucci et al. 2000). In M. persicae, Hsp are essential for survival and several biological processes, such as detoxification of xenobiotics (Figueroa et al. 2007; Ramsey et al. 2007).
M. persicae exhibits dimorphism as alate (winged/dispersal) and apterous (wingless) forms (Braendle et al. 2006). The switch to the alate form in M. persicae seems to be triggered by several mechanisms in response to sub-optimal environmental conditions such as high density and poor nutrition (Braendle et al. 2006). Alate production (dispersal) is accompanied by up-regulation of olfactory segment D (OSD), take-out like / JH binding protein (TOL), and mitochondrial adenine nucleotide translocase (ANT) (Ghanim et al. 2006). The role of JH in aphid wing dimorphism is still uncertain (Braendle et al. 2006; Schwartzberg et al. 2008), but is probably governed by environmental and genetic stimuli (Brisson 2010). Imidacloprid-induced reproductive stimulation has been associated with wing dimorphisms in M. persicae (Wang et al. 2008). JH biosynthesis via the mevalonate pathway includes a precursor farnesyl diphosphate synthase I (FPPS I) gene (Zhang and Li 2008) and its expression may regulate reproduction in insects (Cusson et al. 2006).
When first instar M. persicae were continuously exposed to sublethal concentrations of imidacloprid, stimulated fecundity in first and second generation adults was observed (Ayyanath et al. 2013). To understand the molecular underpinnings of this phenotypic response, in the present study we examined gene expression in M. persicae exposed to hormetic concentrations of imidacloprid for two generations. Stress response (Hsp60) (Stanley and Fenton 2000), dispersal (OSD, TOL, and ANT) and developmental (FPPS I) genes were examined. We hypothesized that OSD, TOL, and ANT gene expression would correspond to fecundity responses previously reported (Ayyanath et al. 2013), and that Hsp60 gene expression would be up-regulated in initial generation (G0) second instar nymphs. We also predicted that FPPS I gene expression would be down-regulated (Keeling et al. 2004) during adult exposure corresponding to fecundity responses.
In addition, we examined whether insecticide-induced hormesis in insects results in heritable changes in gene expression. DNA methylation is an important epigenetic mechanism in insects that may provide critical contributions to insect developmental and phenotypic variation (Glastad et al. 2011). Therefore, in addition to measuring gene expression, global DNA methylation was measured in our test aphids. We hypothesized that hypermethylation would occur when Hsp60, FPPS I, OSD, TOL and ANT genes were up-regulating.
METHODS AND MATERIALS
Plants and insects
Potato, Solanum tuberosum L. (cv. Kennebec), was grown in a greenhouse in 12.5 cm diameter pots containing Pro-Mix® (Halifax Seed, Halifax, Nova Scotia) potting soil. Plants were watered as needed. Foliage from these plants was used for insect rearing and experiments. M. persicae was obtained from a population infesting broccoli plants (Brassica oleracea L.) in a pesticide free-greenhouse at the Faculty of Agriculture, Dalhousie University, Truro, NS. Aphid cohorts were maintained on excised leaves in clear plastic boxes (37 L × 24 W × 14 H cm) lined with moistened paper towels. Boxes were held in a growth chamber (22 ± 2° C, 16:8 L:D, 65 ± 5% RH) and every second day a layer of freshly excised leaves was placed on one end of the box. The infested foliage on the opposite end was discarded when about 80% aphids moved to fresh foliage. Paper towels were replaced every 10 days.
Chemicals
Imidacloprid (Admire® 240 SC, 240 g a.i. L−1; Bayer CropScience Canada, AB, Canada) was suspended in deionized water to obtain a 1000 μg a.i. L−1 stock solution. The working solutions contained 0.15% Triton™ X 100 (Fisher Scientific, Fair Lawn, New Jersey) as an emulsifier. We previously found that concentrations of 25 μg a.i. L−1 adversely affected M. persicae reproduction. Because we were interested in gene regulation at concentrations below the no observable adverse effects concentration (NOAEC), insecticide concentrations of 0.025, 0.1, 0.25, 2.5 and 10 μg a.i. L−1 were used in the current experiments. Controls in these experiments consisted of 0.15% Triton in water.
Leaf-dip exposure
Potato leaf discs (1.8 cm diameter) were excised using a stainless steel cork borer. Using forceps, leaf discs were dipped in control or insecticide solutions for 5 sec, air-dried for 1 h, and then two discs each were placed in 5.5 cm Petri plates lined with a Whatman No. 1 filter paper. Ten first instar M. persicae (∼24 h old) were transferred to each treated leaf disc. Dishes were covered with a Petri plate cover, placed in sealable plastic containers and held in growth chamber as described above.
Aphids were exposed to and reared continuously on treated leaf discs for two generations (G0, G1). Leaf discs were replaced every second day. The experiment was a completely randomized design, with imidacloprid concentration being the main factor of interest. Each container represented a biological replicate of a concentration and had five Petri dishes, each with ten aphids. For gene expression analyses, five aphids from each Petri plate were randomly collected at day 4, 9, 13 and 17, representing G0 second instar, G0 adult, G1 second instar and G1 adult stages, respectively. Excess nymphs were discarded as soon as 10 nymphs per Petri plate were obtained on day 9 and 17. A total of three biological replicates were set up in the experiment. For the global DNA methylation experiment, the same experimental procedure and design was used.
Sample preparation
Gene expression analyses
Collected aphids were flash frozen in liquid nitrogen. Total RNA was isolated using a RNeasy® mini kit (Qiagen, Toronto, Ontario). Quality (A260/280 > 2.0) and quantity of total RNA was assessed with a Nanodrop ND-1000 (NanoDrop Technologies, Wilmington, Delaware) and gel electrophoresis (rRNA band intensity: 28s = 2X18s). Later, cDNA was synthesized from 1 μg of total RNA using a QuantiTect® Reverse Transcription kit (Qiagen, Toronto, Ontario) and stored at −20° C until further analyses. The primers (Sigma-Aldrich, Oakville Ontario) used for quantitative Real-Time (qRT) PCR are listed in Table 1. Internal controls with cycle of threshold (ct) values closer to the ct of selected genes were chosen to calculate expression fold-change (Pfaffl 2001). For quantification of OSD and TOL genes, the β-actin gene was used as an internal control. Ace was used as an internal control for ANT, Hsp60, and FPPS I genes. qRT PCR was performed on a StepOne™ RT PCR System (Applied Biosystems, Burlington, Ontario) in a 10 μL reaction following the manufacturer's instructions using SYBR green reagent (Applied Biosystems, Burlington, Ontario). The reaction mixture contained 2X SYBR green reagent master mix, 2 μL cDNA, 2.5 μL ultrapure water, and 0.25 μL each of forward and reverse primers (final concentration of 2.5 μM). Data were analyzed from three independent runs using mixed model analysis of variance (PROC MIXED) (SAS 2008). If means were significantly different, they were separated using a LSD test (α = 0.05). Relative quantification (RQ) of gene expression above or below that of controls is reported.
Primer sequences (5‵ → 3‵) for selected and internal control genes used to measure gene expression in Myzus persicae exposed to sublethal concentrations of imidacloprid.

For all the time points, gene expression data was subjected to dose-response modeling (Ayyanath et al. 2013; Cedergreen et al. 2005). Fecundity responses of G0 and G1 adults previously reported (Ayyanath et al. 2013) were correlated/regressed against gene expression in G0 and G1 adults found in the present study using R statistical software with addon package drc (http://www.bioassay.dk).
Global DNA methylation analysis
Samples from aphids were collected as described above, and genomic DNA was isolated using a DNeasy® Blood and Tissue Kit (Qiagen, Toronto, Ontario). From aphids of each concentration and time point, 500 ng of gDNA was digested, using Nuclease P (Sigma-Aldrich, Oakville, Ontario), and phosphorylated, using alkaline phosphatase (Sigma-Aldrich, Oakville, Ontario), to examine global DNA methylation changes using a DNA Methylation EIA™ kit (Cayman Chemical, Ann Arbor, Michigan). A standard curve with r2 > 0.9 was obtained using the kit standard. Data analysis was performed as per the manufacturer instructions after calculating the amount of methylated DNA based on the obtained equation from the standard curve. Relative methylation of DNA to controls was analyzed using PROC MIXED (SAS 2008), with means separations done using a LSD test (α = 0.05).
RESULTS
Gene expression analyses
Continuous exposure to sublethal concentrations of imidacloprid resulted in up- and down-regulation of the genes analyzed, with intra- and inter-generational differences observed (Table 2). Gene expression in treatments is reported as relative quantity (RQ) to that in control aphids and a 2-fold up- or down- regulation was considered as biologically significant (Ghanim et al. 2006).
P-values for a two generation (G0, G1) experiment examining effects on gene expression (up- or down-regulation) and global DNA methylation during imidacloprid-induced hormesis in second instar (N) and adult (A) Myzus persicae. Significant treatment effects were subjected to a least square means separation (LSD, α = 0.05) and reported graphically.

Models attempting to describe gene expression against concentration did not fit (f > 0.1) for any of the studied genes at imidacloprid concentrations that induced hormesis. A weak relationship was found for OSD (R2 < 0.325; P = 0.0130) and TOL (R2 < 0.312; P = 0.0126) gene regulation and G1 adult fecundity. All other correlations and regressions were not significant.
Hsp60 gene
In the first generation (G0), there was 2.5-fold down-regulation of Hsp60 in second instars exposed to 0.25 μg a.i. L−1. In G0 adults, 2.0-fold down-regulation was found in the 0.025 μg a.i. L−1 treatment, and less down-regulation at higher concentrations (Figure 1). In G1, there was 3.2-, 2.5- and 5.3-fold up-regulation of the Hsp60 gene in second instars at 0.25, 2.5 and 10 μg a.i. L−1, respectively. Two- to 3.3-fold down-regulation of the gene was observed at all concentrations in G1 adults.

Heat shock protein (Hsp) 60 gene expression during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instars and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above or below them are significantly different (P ≤ 0.05, LSD test).
FPPS I gene
Approximately 2.0-fold down regulation of the FPPS I gene was observed in G0 second instars exposed to 0.25 and 2.5 μg a.i. L−1 imidacloprid (Figure 2), and in G0 adults from the 0.025 μg a.i. L−1 treatment. At other imidacloprid concentrations changes in FPPS I gene expression was relatively minor in G0. G1 second instars from the 0.025, 2.5 and 10 μg a.i. L−1 treatments had 2.2-, 2.7 and 5.0-fold up-regulation, respectively, of the FPPS I gene. In G1 adults, ∼2.0- to 5.0-fold down-regulation was observed across the range of imidacloprid treatments.

Farnesyl diphosphate synthase (FPPS I) gene expression during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instar nymphs and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above or below them are significantly different (P ≤ 0.05, LSD test).
OSD gene
When G0 first instars were exposed to 0.1 μg imidacloprid L−1, 2.3 fold up-regulation of the OSD gene was observed in G0 second instars, and at 0.25 μg a.i. L−1 ∼2.0-fold down-regulation was observed (Figure 3). Exposure of G0 nymphs to other concentrations did not change the expression of the OSD gene. In G0 adults, only at 2.5 μg a.i. L−1 was a change in gene expression observed, with ∼2.0-fold down-regulation. G1 second instars exposed to 0.1 and 0.25 μg a.i. L−1, had 3.3- and 2.5-fold down-regulation, respectively. Other concentrations did not alter OSD gene expression at that time point. All concentrations except 0.25 μg a.i. L−1 up-regulated OSD gene expression in G1 adults; 3.5-, 4.0-, 3.3- and 2.8fold up-regulation of the gene was observed at 0.025, 0.1, 2.5 and 10 μg a.i. L−1, respectively.
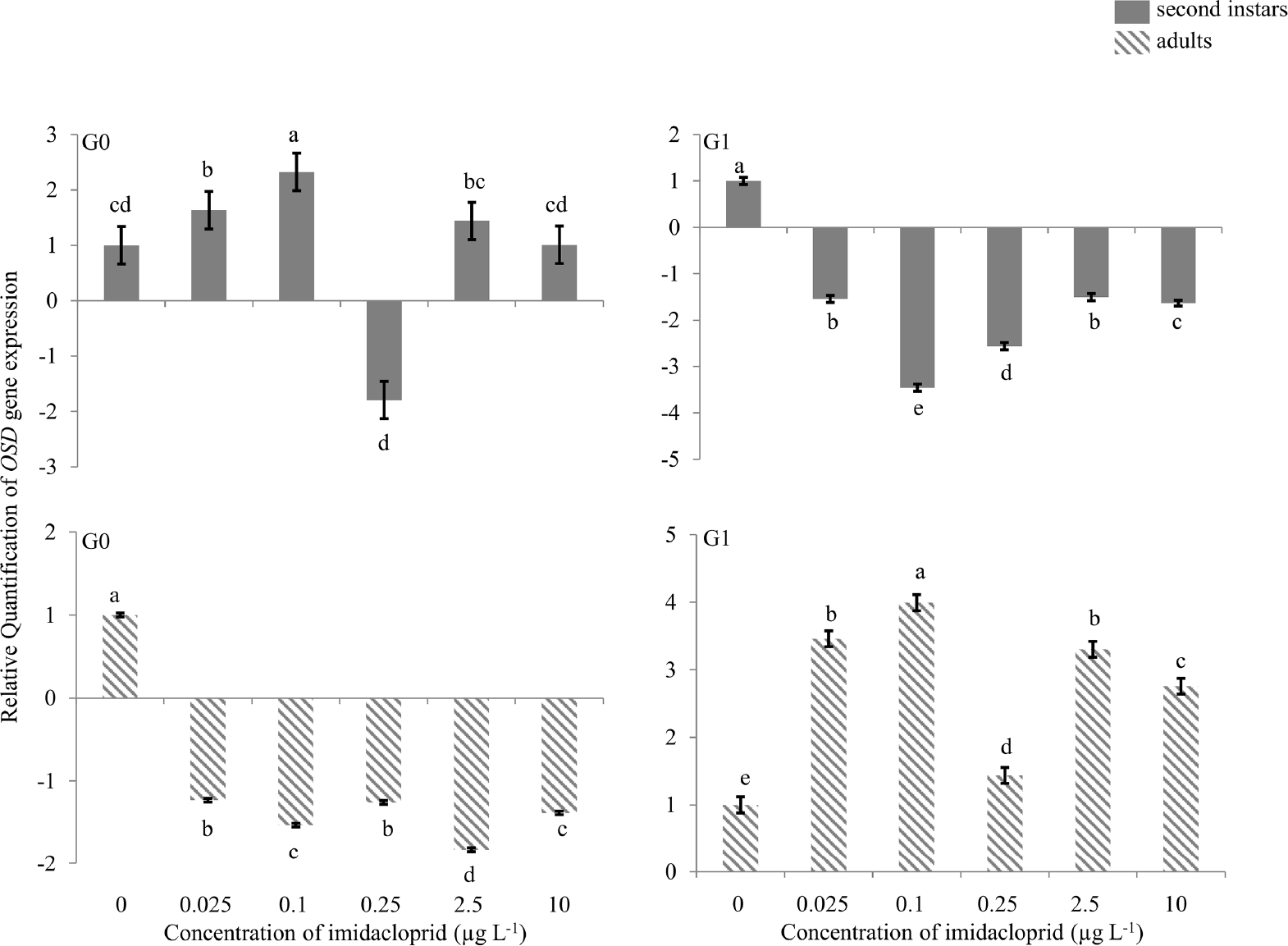
Olfactory Segment-D (OSD) gene expression during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instar nymphs and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above or below them are significantly different (P ≤ 0.05, LSD test).
TOL gene
There was 2.0-fold up-regulation of the TOL gene in G0 second instars exposed to 0.1 μg a.i. L−1 imidacloprid (Figure 4), and 3.3- and 10.0-fold down-regulation at 0.25 and 10 μg a.i. L−1, respectively. In G0 adults, a change in TOL gene expression (2.0-fold down-regulation) was only observed at 10 μg a.i. L−1. In G1 second instars, there was 4.0-, 4.0-and 6.0 fold up-regulation at 0.025, 0.25 and 2.5 μg a.i. L−1, respectively, and 2.0-fold down-regulation at 10 μg a.i. L−1. In G1 adults, 2.6- and 3.0fold up-regulation of the TOL gene was observed at 0.025 and 10 μg a.i. L−1 of imidacloprid.
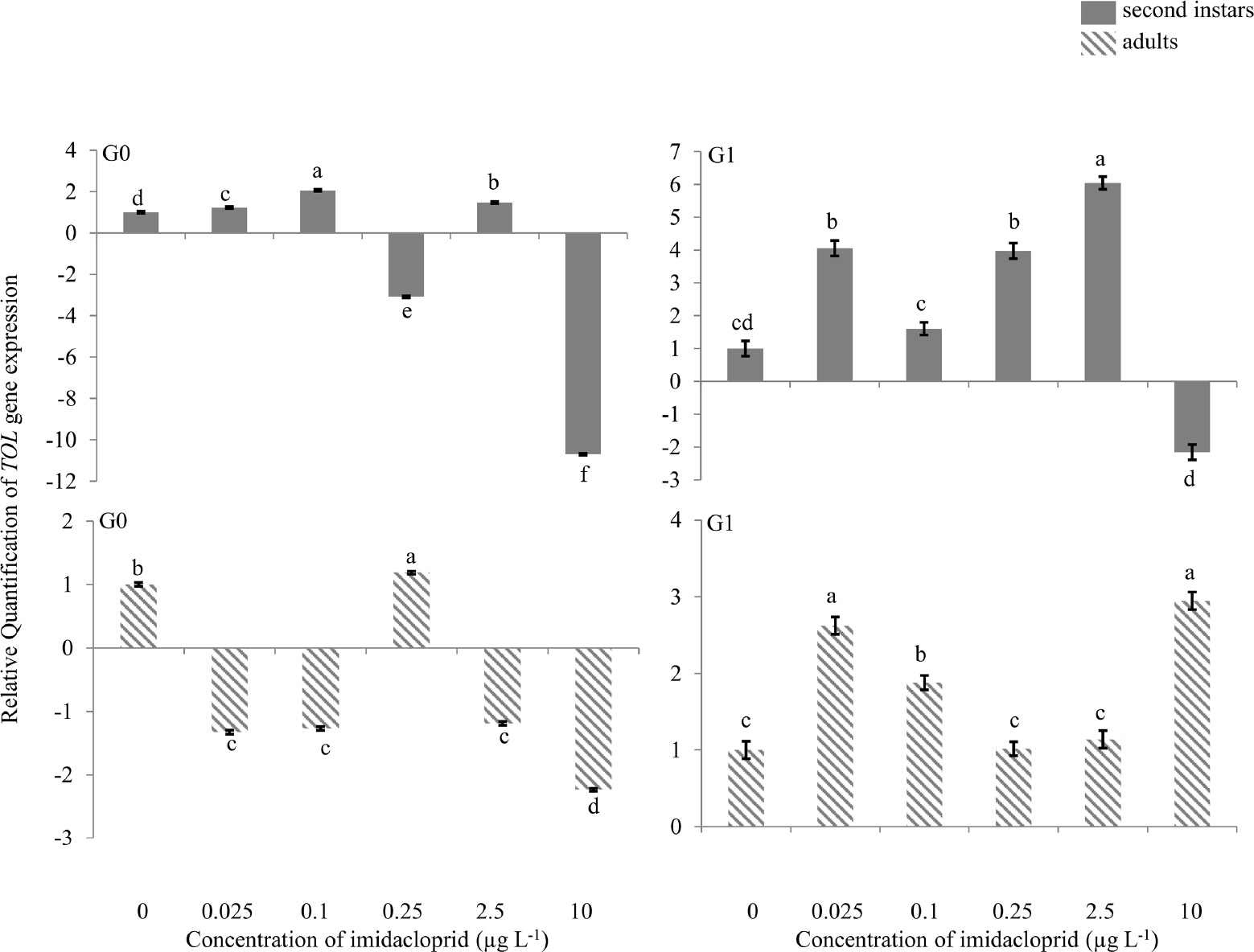
Take-out like (TOL) gene expression during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instar nymphs and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above or below them are significantly different (P ≤ 0.05, LSD test).
ANT gene
There was no change in expression of the ANT gene in G0 nymphs or adults (Figure 5). In G1 second instars, there was 4.5-, 2.0-, 6.4-, 4.2- and 4.2-fold up-regulation at 0.025, 0.1, 0.25, 2.5 and 10 μg a.i. L−1. In G1 adults, 2.5- to 5.0-fold down-regulation was observed at all imidacloprid exposure concentrations.
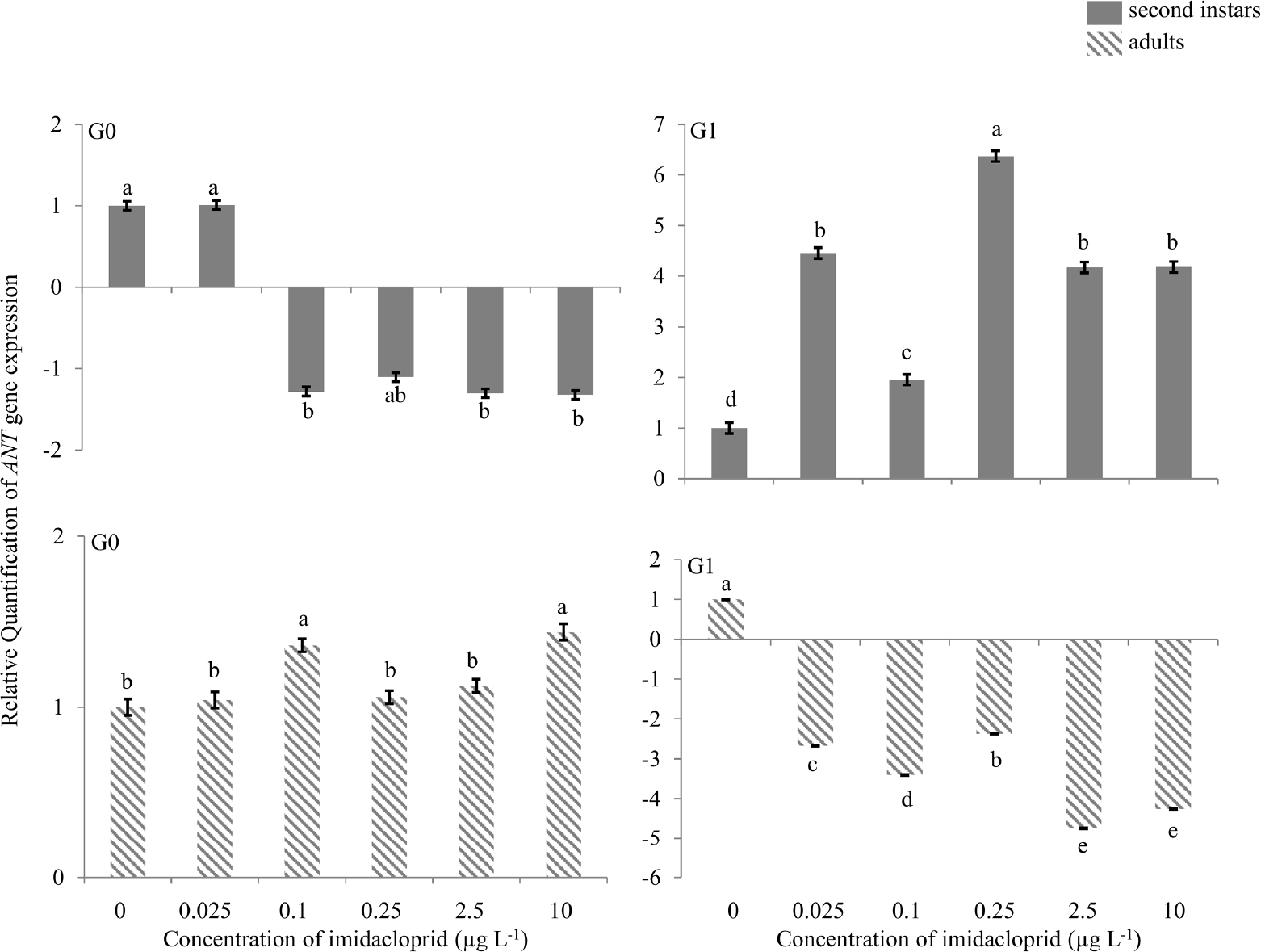
Adenosine nucleotide translocase (ANT) gene expression during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instar nymphs and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above or below them are significantly different (P ≤ 0.05, LSD test).
Global DNA methylation analysis
When first instar M. persicae were exposed to sublethal concentrations of imidacloprid and G0 second instars were analyzed for global DNA methylation, approximately 25, 10 and 20% hypermethylation above the control was observed at 0.025, 0.1 and 0.25 μg a.i. L−1, respectively, and about 10% hypomethylation was observed at 2.5 μg a.i. L−1 (Figure 6). No change in methylation was detected at 10 μg a.i. L−1. In G0 adults, 10–60% hypermethylation was seen at various imidacloprid concentrations. In G1 second instars, significant hypermethylation in the order of 50–60% above the control was found only at 2.5 and 10 μg a.i. L−1. In G1 adults, hypomethylation of 25% and 10% occurred at 0.25 and 10 μg a.i. L−1, respectively, while 10% hypermethylation was observed at 2.5 μg a.i. L−1. Other concentrations did not show any change in global DNA methylation relative to control (Figure 6).
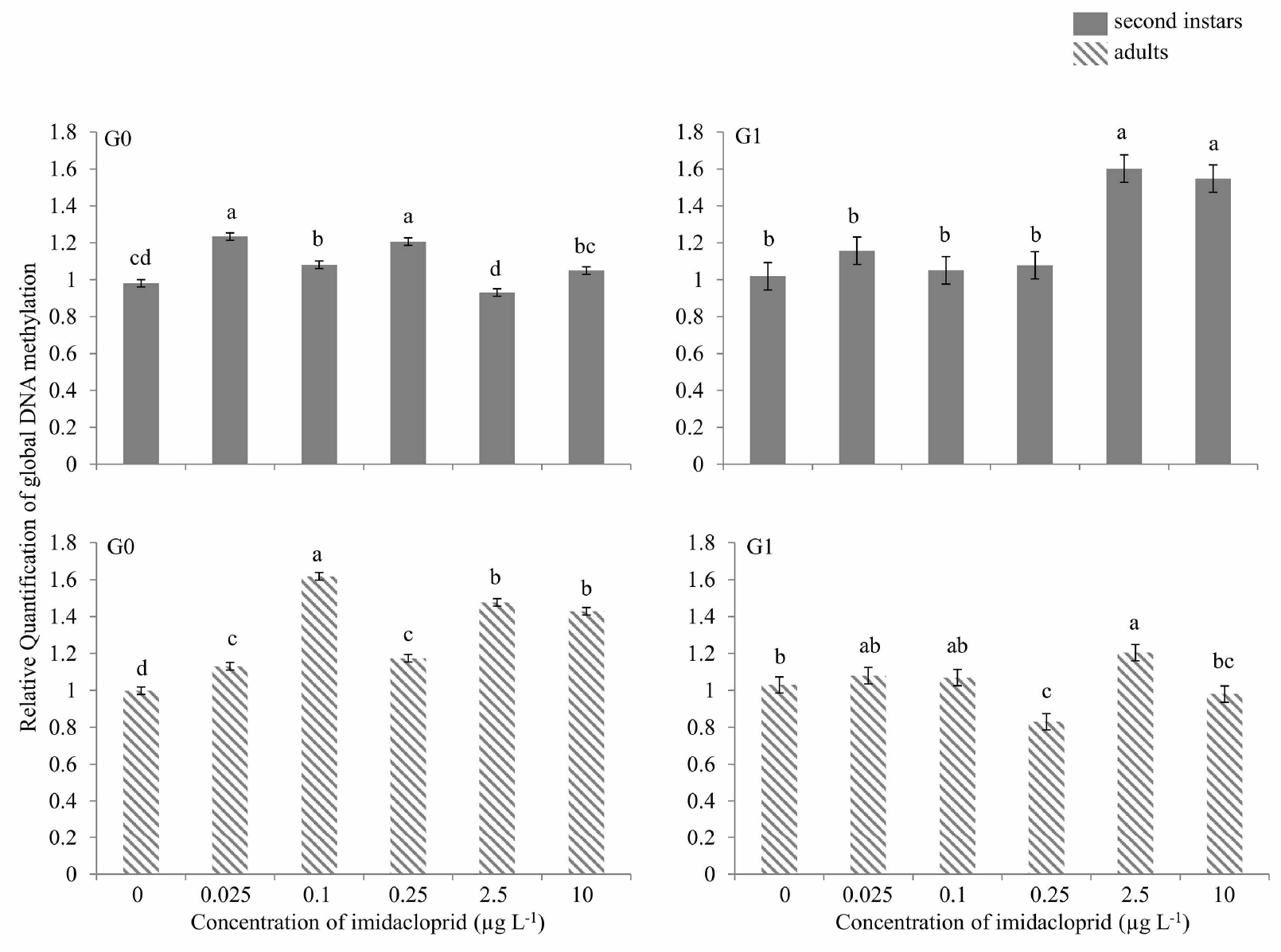
Global DNA methylation during imidacloprid-induced hormesis in Myzus persicae. Test aphids representing second instar nymphs and adults from initial (G0) and succeeding (G1) generations were used for down-stream analyses. Error bars represent standard error of means. For a given time point, bars with different letters above them are significantly different (P ≤ 0.05, LSD test).
DISCUSSION
We previously reported intra- and inter-generational differences in reproduction hormesis when green peach aphids were exposed to low doses of imidacloprid (Ayyanath et al. 2013). Using the same insect, insecticide, and doses, here we show that up- or down-regulation of stress, mitochondrial function, dispersal, and developmental genes occurs when first instar M. persicae are continuously exposed for two generations to sublethal concentrations of imidacloprid. In multiple instances, there was evidence of up-regulation of genes in one life stage or generation that was countered by down-regulation of the same genes in subsequent life stages or generations.
Hsp70 expression has been associated with reduced fecundity/reproduction in L. huidobrensis (Huang et al. 2007), Drosophila melanogaster (Hercus et al. 2003), Trialeurodes vaporariorum (Westwood), and Bemisia tabaci (Gennadius) (Cui et al. 2008). This is possibly attributable to accumulation of Hsp and associated traits (thermo-tolerance) at a cost of impaired fecundity. In this study, continuous exposure of M. persicae to sublethal concentrations of imidacloprid previously found to stimulate reproduction had no effect or resulted in down-regulation of the Hsp60 gene in second instars and adults of the initial generation, followed by up-regulation in second instars of the succeeding generation. Hsp expression has been shown to vary depending on the stressor type (Sanders et al. 1995; Veldhuizentsoerkan et al. 1991) and scenario of exposure to a stressor (Helmcke and Aschner 2010).
Down-regulation of Hsp expression in insects not well understood (Mahroof et al. 2005). Alternation of up- and down-regulation occurred across generations and between nymphs and adults within a generation in the present study. For example, up-regulation of Hsp expression in G1 second instars was followed by down-regulation in adults. Similar alternations were found in Hsp70 expression in egg, larva, pupa, and adult of T. castaneum exposed to mild heat stress (Mahroof et al. 2005). Stimulated fecundity in M. persicae exposed to hormetic concentrations of imidacloprid (Ayyanath et al. 2013) could be due to down-regulation of Hsp gene in G0 and G1. Down-regulation of Hsp90 and Hsp70 was reported in rat brains exposed to various concentrations of DDT (Shutoh et al. 2009) and bronchial cancer in smokers with chronic obstructive pulmonary disease (Cappello et al. 2006). In these studies, down-regulation of Hsp expression was suggested to be a recovery response to regain homeostasis following chronic exposure to mild levels of stress. Reduced Hsp70 expression can also be associated with short-term stress, as seen when C. tentanus recovers from exposure to heat shock (Karouna-Renier and Zehr 1999).
Up-regulation of Hsp60 follows an accumulation of unfolded (damaged) proteins resulting from stress or injury in an organism (Parsell and Lindquist 1993), and accumulation of Hsp often results in decreased fecundity (Huang et al. 2007). Hsp60 up-regulation was observed in M. persicae G1 nymphs exposed to higher sublethal concentrations of imidacloprid (0.25–10 μg a.i. L−1), and reduced reproductive outputs were previously seen in G1 aphids exposed to these imidacloprid concentrations. Due to overcompensation following insecticide exposure in G0, the insect probably became conditioned to higher insecticide concentrations in G1 by elevating expression of Hsp60. Alternatively, accumulation of damaged proteins triggered the up-regulation of the gene.
Fecundity, reproduction, and metamorphosis in insects is controlled by JH (Dawson et al. 1987; Hartfelder 2000; Verma 1981). In aphids, the FPPS I gene is an important enzyme regulator and its down-regulation may increase JH titers in females (Keeling et al. 2004). It catalyzes formation of farnesyl diphosphate (FPP), a precursor needed in biosynthesis of JH, alarm pheromones and sex pheromones (Dawson et al. 1987; Vandermoten et al. 2009). JH is known to stimulate reproduction in M. persicae and elevated JH titers in maternal aphids inhibit wing development and promote development of apterous forms (Tamaki 1973; Verma 1981). In pea aphid, Acrythosiphon pisum, the mother that perceives stress cues such as crowding transmits the information to unborn progeny (Brisson 2010). Similarly, it is possible that G0 M. persicae adults stressed by sublethal imidacloprid exposure passed the signal to G1 nymphs, as indicated by the strong up-regulation of the FPPS I gene. This was countered (compensated) in G1 adults, as observed by down-regulation of the FPPS I gene, again coinciding with the hormetic (stress) response previously seen at this stage (Ayyanath et al. 2013).
It was previously shown that stimulated fecundity in M. persicae exposed to hormetic concentrations of imidacloprid resulted in increased JH III titers (Yu et al. 2010). This suggests that increased M. persicae fecundity and JH titers may be correlated with down-regulation of the FPPS I gene, since down-regulation of this gene was previously shown to accompany increase JH titers in an insect (Keeling et al. 2004). These deductions should be made with caution and considered only where the time points used in correlating JH titers and fecundity correspond. In the present study, down-regulation of FPPS I observed in G0 adults exposed to 0.025 μg a.i. L−1 and in G1 adults exposed to 0.025 and 0.1 μg a.i. L−1 was previously associated with higher aphid fecundity (Ayyanath et al. 2013). Significant down-regulation of FPPS I was also seen at imidacloprid concentrations that did not result in stimulated reproduction, suggesting that other genes, also needed for JH production, were not affected by the insecticide treatment.
Dispersal-related genes such as OSD, TOL and ANT are more highly expressed (2–5 fold) in alates than apterous aphids (Ghanim et al. 2006). In the present experiment, OSD, TOL and ANT gene expression varied in apterous individuals exposed continuously to sublethal hormetic concentrations of imidacloprid. Complete wing development was not observed, but at higher concentrations small wing pads appeared in some insects (data not shown).
OSD gene up-regulation has previously been shown to be inversely related to aphid fecundity (Bos et al. 2010). We found that up-regulation of the OSD gene in G0 second instar M. persicae exposed to certain imidacloprid concentrations was followed by down-regulation of the gene in G0 adults, and even greater down-regulation in G1 second instars. Immature G0 aphids stressed at certain imidacloprid concentrations that resulted in up-regulation of the OSD gene may have conditioned subsequent adults for increased reproduction. In G0 adults, OSD gene down-regulation occurred at all concentrations, possibly indicating compensation to the stress, coinciding with increased reproduction at those same concentrations (Ayyanath et al. 2013). Likewise, OSD gene down-regulation in G1 second instars occurred at concentrations of imidacloprid identical to those previously shown to elicit a hormetic reproductive response in M. persicae. Trends similar to OSD gene regulation were observed with TOL. This gene is associated with chemoreception (Fan et al. 2011; Jacobs et al. 2005; Weil et al. 2009) and circadian control of feeding behavior. TOL can be induced by starvation (Fujikawa et al. 2006; Weil et al. 2009), juvenile hormone binding proteins in response to fluctuating JH titers (Bohbot and Vogt 2005) during courtship and mating, or when regulating antennal responses to food, hosts, or pheromones (Dauwalder et al. 2002). The complexity of this gene and its involvement in multiple functions does not readily permit direct correlation with the hormetic responses previously observed (Ayyanath et al. 2013).
The ANT gene regulates mitochondrial proteins that function as carriers of important metabolites involved in a number of mitochondrial processes, mainly catalyzing exchange of ADP for ATP across inner mitochondrial membranes (Zhang et al. 1999). The unresponsiveness of the ANT gene in G0 suggests that exposure to hormetic concentrations of imidacloprid results in no additional energy expenditures for the aphids. However, as with the TOL, Hsp60 and FPPS genes, significant up-regulation of ANT occurred in G1 nymphs, indicating extra energy was expended. This suggests that even if no extra energy was expended in G0 adults, the insect can still have stimulated reproduction, with energy expenditure effects felt in offspring. Down-regulation of the ANT gene in G1 adults probably reflects the cessation of energy requirements and or other regulatory process (up-regulation in second instar) during the development phase in G0 nymphs.
Recent experiments indicate that environmental fluctuations can induce specific and predictable epigenetic-related molecular changes (Vaiserman 2012). It is unclear if insecticide-induced hormesis in insects is an epigenetic process, resulting in heritable changes in gene expression (Glastad et al. 2011; Gressel 2011). DNA methylation is an important epigenetic mechanism for regulation of gene expression. In insects, DNA methylation is thought to play a role in developmental responsiveness to environmental factors and may provide critical contributions to insect developmental and phenotypic variation (Bass and Field 2011; Glastad et al. 2011; Gressel 2011). Increases in hypermethylation, typically reduce DNA transcription and usually result in inactivation of genes, although this is not always the case (Suzuki and Bird 2008). Concurrent amplification and methylation of an esterase gene in greenbug, Schizaphis graminum (Rondani), suggested heritability rendering increased resistance to organophosphorus insecticides (Ono et al. 1999). Similarly, in M. persicae, the E4 gene is important in expression/amplification of insecticide-detoxifying esterases, and reduced DNA methylation in this insect coincides with a loss of E4 gene expression (Field et al. 2004; Hick et al. 1996). In the present study, methylation occurred initially in G0 nymphs and adults, and in at only high imidacloprid concentrations G1 nymphs. This was followed by an absence of methylation at any concentration in G1 adults, possibly indicating a heritable adaptation to the sub-NOAEC concentrations of imidacloprid we tested. This suggests that trans-generationally the insect would be able to cope with higher concentrations, which has repercussions for resistance development (Gressel 2011).
Overall, there are complex patterns of gene expression across generations in insects exposed to hormetic concentrations of insecticide. We interpret the up- or down-regulation of Hsp60, FPPS I, OSD, TOL and ANT genes in G0 to be a priming response (Costantini et al. 2010) in preparation for further adverse conditions, vis a vis insecticide exposure. Imidacloprid resistance development in insects via mutations has been reported in the past (Bass and Field 2011; Wen et al. 2009) and insecticide resistance is associated with amplification and methylation of esterase genes (Field 2000; Field et al. 2004; Ono et al. 1999). We speculate that insecticide-induced hormesis might serve as a precursor to insecticide tolerance and ultimately resistance (Gressel 2011). Further investigation is needed in this area to fully comprehend how insect response to low levels of stress in the form of hormesis relates to induction of detoxification enzymes and methylation changes across generations.
Footnotes
ACKNOWLEDGEMENTS
Financial support for this project was through the Ontario Graduate Scholarship Program and several University of Guelph internal scholarship donors (scholarships to M-M.A), the Natural Sciences and Engineering Research Council of Canada (NSERC Discovery Grants to G.C.C. and B.P.), and the Canada Foundation for Innovation (Leaders Opportunity Funds to G.C.C. and B.P.)