
Abstract
The genetic consequences resulting from environmental exposure to ionizing radiation have a significant impact on both radiation regulatory policies and the comprehension of the human health risks associated with radiation exposure. The primary objectives of the study were to assess 1) genotoxicity of exposure to radiation as a function of absorbed dose and dose rate, and 2) induction of a radio-adaptive response following a priming dose at varying dose rates. Results demonstrated that sub-acute environmental exposures of 10cGy gamma radiation resulted in indistinguishable levels of chromosomal damage as compared to controls. A radio-adaptive response was observed in all experimental groups, exposed to a subsequent acute challenge dose of 1.5 Gy, demonstrating that low dose rates of low energy transfer (LET) radiation are effective in reducing genetic damage from a subsequent acute low-LET radiation exposure. Furthermore, the data presented herein demonstrate a potential beneficial effect of sub-chronic exposure to low levels of low-LET radiation in an environmental setting and do not support the Linear No Threshold (LNT) hypothesis.
INTRODUCTION
Radioactive contamination in an environmental setting is particularly useful to evaluate biological responses, since environmental radionuclides are the most likely source of human exposure. Due to the lack of access to contaminated sites, in which to study ionizing radiation, many studies are performed utilizing acute or fractionated dosing from 137Cs, 60Co, and X-ray sources. Although low dose rates may be obtained in the laboratory by protraction, these may not adequately represent true environmental radiation exposures. Differential biological responses result from radiation administered acutely, via an irradiation source, versus low dose rate continuous irradiation in an environmental setting. Few studies report the effects of continuous, low dose rate irradiation in whole animal systems in an environmentally relevant context. Furthermore, previously documented in vitro cellular responses need to be assessed in terms of comparable absorbed gamma ray doses, and dose rates, as well as the resulting response of an organism (Bosi & Olivieri, 1989; Cai & Liu, 1990; Cai & Wang, 1995). Due to the complexity of cellular interactions within biological systems, organismal responses to ionizing radiation may differ from responses of cultured cells. Therefore, studies designed to assess responses in animal model systems are necessary to develop a more comprehensive understanding of the potential risks of long term low dose rate gamma irradiation (Calabrese & Baldwin 2002).
Exposures in the Chernobyl environment currently are from low-LET (linear energy transfer), low dose rate ionizing radiation, primarily γ radiation from 137Cs and 90Sr as well as limited amounts of β radiation. The highest levels of radiation are found in the Red Forest, 1.5 km West of the destroyed reactor, although levels are nonetheless considered low dose, low dose rate radiation. Several studies have sought to define what constitutes a low dose. Epidemiological studies have demonstrated that doses of ≤ 20 cGy cause few biological effects, regardless of the dose rate, whereas similar results are found for studies conducted on cultured cells receiving doses of ≤ 2 cGy (UNSCEAR 2000). Additional studies evaluating tumor induction report that regardless of the total dose, when the dose rate is about 0.1 mGy min−1, the dose should be considered low (UNSCEAR 2000). In the 1993 UNSCEAR report, the recommendation was that in evaluating the risk of tumor incidence in humans, a reduction factor should be applied to doses below 200 mGy or dose rates under 0.1 mGy min−1. Most recently, a low dose has been defined as a total dose of 10cGy or less (Low Dose Radiation Risk Assessment, 2006; U.S. Department of Energy Low Dose Radiation Research Program; http://www.lowdose.energy.gov). However, this subject remains controversial and more data are needed to establish appropriate radiation regulatory guidelines regarding the mitigation of human health risk.
Studies examining cancer incidences in individuals living in areas of high natural background radiation, documented no significant differences in the frequency of cancers than those in adjacent areas with significantly lower background radiation levels. In fact, lower cancer rates occurred in the areas with higher radiation background levels (Wang 1993; UNSCEAR 1994). Observations of the beneficial effects of in vivo low dose radiation exposures include enhanced longevity, growth, immune response and DNA repair (Wang and Cai 2000; Upton 2001; Atvi et al. 2005). This beneficial or stimulatory effect has been termed hormesis. Calabrese & Baldwin (2002) concluded that many studies were not designed to directly assess the phenomena of hormesis, yet supported it via unexpected experimental outcomes.
Radio-adaptive response
One form of hormesis is referred to as the “radio-adaptive response”. A radio-adaptive response is defined as a reduction in the adverse effects of exposure to a low dose of a toxicant due to the activation or up-regulation of endogenous physiological mechanisms. Often, the response to low level exposures will confer protection to the organism by enhancing the ability to withstand exposure to higher doses of a stressor. Reported herein are data from experiments designed specifically to document a radio-adaptive response to low dose, low dose rate, low-LET gamma radiation and generate samples for further exploration of the mechanisms involved. Due to the specificity of the experimental design, to examine only low-LET effects, it should be noted that the findings reported herein may not apply, or be suitable for comparison to, studies examining high-LET radiation sources such as radon and plutonium-associated alpha radiation.
In this study, male BALB/c mice were exposed to environmental radiation contamination, in the Red Forest near the Chernobyl Nuclear Power Plant (ChNPP). The primary objectives of the study were 1) to assess genotoxicity after exposure to radiation as a function of dose and dose-rate, and 2) to assess induction of a radio-adaptive response following a priming dose administered at varying dose rates.
Biological Endpoints for Detection of Radiogenic DNA damage
There are a variety of assays available to detect DNA damage produced by ionizing radiation. Methods which detect double strand breaks (DSB) are particularly useful since unrepaired DSB are the primary lesions of concern and are relatively simple to detect. One cytogenetic biomarker with widespread applicability is the micronucleus (MN) test. Micronuclei are formed during the last mitosis in erythroblast differentiation in the bone marrow (Hayashi et al. 1984; Krishna and Hayashi 2000). The MN in vivo test is especially useful as the test allows consideration of naturally-occurring biological processes such as DNA repair, metabolism, and pharmacokinetics, all of which influence the yield of MN (Krishna and Hayashi 2000). When ionizing radiation produces a DSB during erythroblast differentiation, a chromosomal fragment will be produced that likely lacks a centromere; therefore, there is no region for the spindle fiber to attach. Thus, as the erythroblast proceeds through the cell cycle, the chromosome fragment is not incorporated into the nucleus of the daughter cell. Immediately following cellular division, the erythroblast matures into a polychromatic erythrocyte (PCE) and the nucleus is extruded. Chromosome fragments will remain in the cytoplasm of the otherwise DNA-free PCE. DNA left behind in a mature retic-ulocyte (PCE or NCE, normochromatic erythrocyte) is classified as a micronucleus (MN) and is indicative of DNA damage. Micronuclei may be effectively evaluated following exposure to clastogenic agents in both the bone marrow (Chaubey et al. 1977; Heddle 1973) and peripheral blood (Chaubey et al. 1993; Dertinger et al. 1996). Many researchers have documented elevated MN frequencies in precancerous tissues (Benner et al. 1993; Garewal et al. 1993; Rosin 1993; Herens et al. 1995). Furthermore a link between MN and cell malignancy has been discovered, indicating that MN formation could be indicative of early steps in carcinogenesis (Cai 1988).
METHODS
Dosimetry
Soil concentrations of radiocesium (137Cs) and radiostrontium (90Sr) were determined from 420 sample stations in the Red Forest, located 1.5 km west of the destroyed reactor. Knowledge of the 137Cs and 90Sr activities from previous studies (Chesser et al. 2004) facilitated estimating the dose rates that would be encountered by mice at each location. We revisited three sites to confirm that total whole body absorbed doses of 10 cGy gamma radiation could be administered over 10, 20, and 45 days of exposure. Soy-paraffin phantoms (density = 0.98) were constructed using molds made to the average dimensions of laboratory mice (body length = ca. 85mm, determined from measurements of BALB/c during previous experiments; Figure 1). Channels in the phantoms allowed insertion of lithium fluoride (LiF) thermolumiscent dosimeters (TLDs) at the positions of the liver, heart, brain, surface, and gonad. The channels were then sealed with clay to prevent moisture from damaging the LiF chips.

Soy wax phantom (density = 0.98) used to confirm total absorbed dose by the insertion of thermoluminescient dosimeters (TLDs). Dimensions of the phantom were determined from averages of body size and mass of BALB/c mice. Dosimetry data are presented in Table 1.
For dose calibration, assembled phantoms were placed into field enclosures for 24 hours prior to the introduction of the mice (N=2 per enclosure, where the mice were to be placed). Phantoms were also exposed in the laboratory to a 137Cs source, for acute doses of 10 cGy and 1.5 Gy, to confirm the intended acute doses (N=3 per dose, in the same chambers where the mice were to be placed). Fluorescence of the TLDs was measured with a Harshaw 3600 TLD reader and compared to calibrated LiF results. Raw scores of fluorescence were converted to total dose (Grays; Gy) and dose rates (cGy h−1) for each chip. Converted doses for all chips were then averaged for a measure of total absorbed gamma dose.
Exposure and Sampling
All animal husbandry and experimental treatments were in accordance with Animal Care and Use Committee guidelines and recommendations as outlined in the approved protocol # 04009–03.
Age-matched, male BALB/c mice were purchased from a breeding colony in Chernobyl City, Ukraine. Mice were randomly selected and assigned to experimental and control groups. All mice were weighed and peripheral blood samples taken to establish pre-exposure MN frequencies. Mice were also given a unique toe clip for subsequent identification. The toe was preserved in lysis buffer as a pre-exposure DNA source.
Mice were placed in laboratory cages at the three experimental field sites (N=10 per exposure group; expected gamma dose rates of 1.0 cGy d−1, 0.5 cGy d−1, and 0.22 cGy d−1). The cages were protected from disturbance by resident species by placement in covered, wired enclosures as previously described (Rodgers et al. 2001). In addition to the enclosure design, sterile water and Purina Lab Chow were supplied ad libitum, to ensure that radiation exposures were from external sources only. Therefore, internal exposure due to inhalation was considered negligible. Mice were exposed to the above dose rates for a total absorbed dose of 10 cGy over 10, 20, and 45 days, respectively. Dose rates were confirmed using TLDs placed into wax mouse phantoms as described above.
Twenty four hours after removal from the field sites, one half of each experimental group and controls (N=5 per group), was given an acute dose of 1.5 Gy via a 137Cs source in a laboratory in Chernobyl City (Dose rate = 33 cGy h−1). Additionally, an acute dose of 10 cGy (Dose rate = 33 cGy h−1) was administered to mice not previously exposed (N=10), half of which (N=5) were also challenged 24 h later with 1.5 Gy. Control mice were sham-irradiated by being placed under the 137Cs source when it was not in operation (Fig. 2). The 24h post-exposure sampling and challenge dosing regime was based on previous time-course studies in our laboratory (unpublished data) and reports of other researchers demonstrating significant MN-PCE levels at 20 hours post-exposure (Abramsson-Zetterberg et al., 1996; Kagawa et al., 2004; Mozdarani and Saberi, 1994).
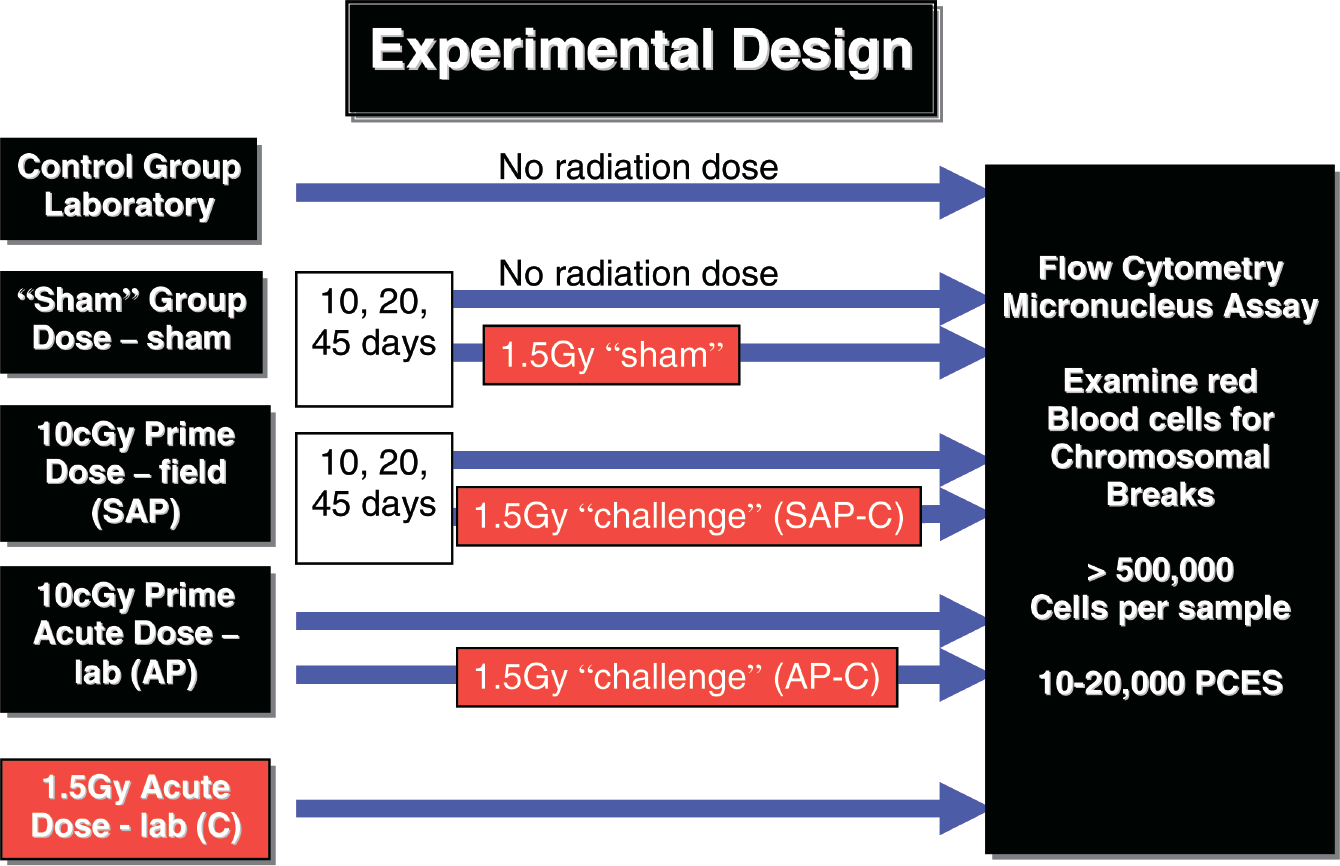
Experimental design of exposure regimes for male BALB/c mice irradiations (N=10 per group). “Control” = laboratory control; “Sham” = sham-irradiated control; SAP (sub-acute primed) = mice receiving a total absorbed gamma dose of 10cGy at varying dose rates over either 10, 20 or 45 days; AP (acute primed) = mice receiving an acute laboratory dose of 10cGy via a 137Cs source; C (challenge dose) = mice receiving an acute laboratory dose of 1.5Gy via a 137Cs source; SAP-C (sub-acute primed – challenged) = one half of SAP group (N=5) receiving an acute laboratory dose of 1.5Gy; AP-C (acute primed-challenged) = one half of AP group (N=5) receiving an acute laboratory dose of 1.5Gy. The micronucleus assay was used to assess the response in terms of the number of polychromatic erythrocytes (PCES) containing DNA damage in the form of micronuclei (MN).
Flow Cytometry
Mice were euthanized, according to approved protocol, 24 hours following the final irradiation. Vital organs and tissue (heart, kidney, liver, spleen, gonads, muscle, and bone) were flash frozen in liquid nitrogen for subsequent analyses. Peripheral blood was obtained immediately following euthanization by severing the hepatic artery. Approximately 100 μL of blood was drawn into a pipet coated with an anti-coagulant and transferred to a tube containing 250 μL of the same. Peripheral blood samples were fixed according to the μFlow Basic-M® kit instructions (Litron Laboratories, Rochester, NY, USA). Samples were analyzed according to the methods of Dertinger et al. (1996). At least 500,000 total erythrocytes were analyzed per sample.
MN Frequencies
Data were analyzed initially as individual groups making all possible comparisons between groups (t-test, p ≤ 0.05; data not shown). No significant differences were observed and for ease of interpretation, exposure groups were subsequently analyzed by pooling all sub-acute groups (those that received 10 cGy over 10, 20, or 45 days) and classifying them as either sub-acute primed (SAP) or sub-acute primed and challenged (SAP-C). Experimental groups that received acute irradiation were classified as either acute-primed (AP) or acute primed challenged (AP-C). The exposure group that received only the acute challenge dose was classified as (C).
To test for normality of the original data, t-tests were employed. Micronucleus frequency data were then normalized by arcsine transformations and subsequently analyzed by a univariate ANOVA, followed by Tukey's multiple comparison test. The significance level (α) was set a priori at 0.05 for all analyses.
RESULTS
Dosimetry
Targeted versus observed gamma dose and dose rate values were confirmed by TLDs from soy wax phantoms. Data are presented in Table 1. The observed dose and dose rates in all exposure groups were within 91.3–97.8% of the expected values.
Expected and observed total absorbed gamma dose values obtained from TLDs at 10, 20, and 45 day field sites.

MN Frequencies
To evaluate the extent of chromosomal damage resulting from 10 cGy, comparisons were made between SAP, AP, control, and sham-irradiated groups. Mean % MN frequencies, after 10cGy total absorbed gamma dose, are presented in Figure 3. The mice receiving 10cGy in the field (Fig. 3, 10, 20, 45d; pooled for SAP group in Fig. 4) did not demonstrate a significant increase in MN induction when compared to either the control or sham-irradiated groups; however, the mice receiving an acute 10cGy dose (Fig. 3, Acute; AP in Fig. 4) had significantly higher MN levels than any other group among these comparisons (*, Fig. 3, t-test, p= 0.006, 0.004, 0.01 compared to 10, 20 and 45d, respectively). These results highlight the importance of dose rate in the clastogenic potential of ionizing radiation.
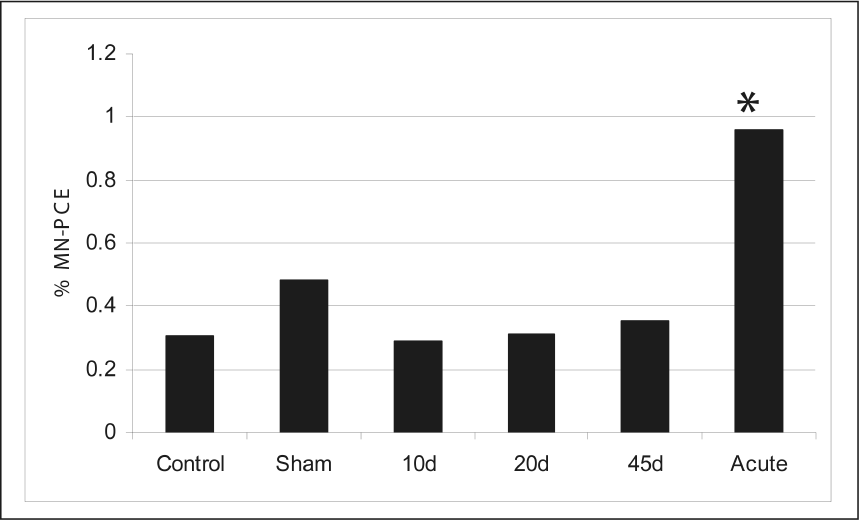
Mean micronucleus (MN-PCE) frequencies in male BALB/c mice following 10cGy total absorbed gamma dose (N=10 individuals per group; ≤ 20, 000 cells per individual analyzed). “Control” and “sham” mice received no irradiation. Mice in the 10d, 20d and 45d groups, received a total dose of 10cGy in field enclosures. “Acute” mice received a total acute dose of 10cGy in the laboratory via a 137Cs source. The acute group was significantly different than all other groups (*p ≤ 0.01). No other significant differences were observed among other comparisons.

Comparison of mean micronucleus (MN-PCE) frequencies in all groups with pooled data from sub-acute groups (SAP = 10cGy in 10, 20, or 45d in Fig. 3). Control, Sham, SAP, AP, C, SAP-C and AP-C designations correspond to and are defined in more detail in the legend of Figure 2. The group of mice receiving only the acute challenge dose of 1.5Gy (*, C; p≤0.05) had a significantly greater %MN-PCE than all other groups. Mice receiving a priming dose of 10cGy administered sub-acutely (SAP) were not significantly different than the control or the sham values. The AP (acute primed) group had a significantly greater level of chromosome damage (♦; p≤0.01) than the SAP (sub-acute primed) group after the 10cGy priming dose. Both SAP-C and AP-C groups had significantly lower MN-PCE (; p≤0.05) than the mice that received only an acute dose of 1.5Gy (C), but AP-C had significantly fewer MN than the SAP-C (§, p≤0.05).
To determine the induction of an adaptive response, groups that were both primed and challenged (SAP-C and AP-C, Fig. 4) were compared to mice that received only the challenge dose (C, Fig. 4). Both groups of mice that were challenged, subsequent to receiving a10cGy gamma dose, either administered as a sub-acute ( , SAP-C, p=0.01; Fig. 4) or an acute dose (
, SAP-C, p=0.01; Fig. 4) or an acute dose ( , AP-C, p=0.001; Fig. 4), presented significantly lower MN counts than did the group receiving only the 1.5Gy challenge dose (C, Fig. 4). Interestingly, the mice receiving an acute priming dose (AP-C, Fig. 4) had significantly lower MN counts after the challenge dose (§, p=0.04) than mice that received the priming dose over 10, 20 or 45d (SAP-C, Fig. 4). The reverse was observed in response to the 10cGy exposure; the MN frequency increased significantly when the dose was administered acutely (*, Acute, p≤0.01; Fig. 3.). These data again demonstrate the importance of dose rate. If chromosomal damage resulting from multiple exposures were additive, it would be expected that the mice receiving the acute priming dose would incur significantly more damage from the subsequent challenge dose than mice receiving the sub-acute priming dose, but notably, the mean %MN-PCE was not significantly different after the challenge dose (AP compared to AP-C in fig. 4).
, AP-C, p=0.001; Fig. 4), presented significantly lower MN counts than did the group receiving only the 1.5Gy challenge dose (C, Fig. 4). Interestingly, the mice receiving an acute priming dose (AP-C, Fig. 4) had significantly lower MN counts after the challenge dose (§, p=0.04) than mice that received the priming dose over 10, 20 or 45d (SAP-C, Fig. 4). The reverse was observed in response to the 10cGy exposure; the MN frequency increased significantly when the dose was administered acutely (*, Acute, p≤0.01; Fig. 3.). These data again demonstrate the importance of dose rate. If chromosomal damage resulting from multiple exposures were additive, it would be expected that the mice receiving the acute priming dose would incur significantly more damage from the subsequent challenge dose than mice receiving the sub-acute priming dose, but notably, the mean %MN-PCE was not significantly different after the challenge dose (AP compared to AP-C in fig. 4).
Expected and observed MN-PCE frequencies are reported in Table 2. These data demonstrate that the sub-acute priming dose administered 24 hours prior to a subsequent challenge dose reduced MN-PCE by 42% when compared to mice that received the challenge dose only. Furthermore, the acute priming dose administered 24 hours prior to a subsequent challenge dose reduced MN-PCE by 69%.
Expected and Observed Mean MN-PCE Frequencies

Note. Expected values were calculated by adding the observed priming dose %MN-PCE value and the observed %MN-PCE value of 1.5 Gy (4.50).
DISCUSSION
The data reported herein describe an assessment of chromosomal damage resulting from ionizing radiation at varying gamma dose and dose rate regimes in a whole animal system, utilizing exposures to contamination in the Chernobyl environment. MN-PCE frequencies were evaluated following a total absorbed dose of 10 cGy, as well as MN-PCE induction levels in mice receiving 10 cGy, followed by a 1.5 Gy challenge dose.
An absorbed dose of 10 cGy gamma, administered acutely (dose rate = 33 cGy h−1) resulted in significant chromosomal damage registered as MN-PCE; however, the same dose administered sub-acutely (gamma dose rates = 0.04167 h−1, 0.02083 h−1, and 0.009167 h−1) resulted in MN-PCE induction levels that were indistinguishable from the control and sham-irradiated groups. This outcome could be explained by either highly efficient DNA repair processes and/or cytotoxicity via apoptosis at these dose rates.
The comparison of various primed and challenged groups enabled determination of whether a radio-adaptive response was induced, and comparison of the efficacy of varying dose rates for induction. A significant reduction in MN-PCE levels was documented when a priming dose of 10cGy was administered prior to a 1.5Gy acute challenge dose. Micronuclei induction in both groups were significantly lower than in mice receiving only the 1.5Gy acute dose, yet significantly higher than either the control or sham-irradiated mice. Thus, a radio-adaptive response was observed in both sub-acute and acute dosing regimes. However, acute administration of the priming dose appears to confer greater protection against a subsequent dose of 1.5 Gy. The priming dose indeed induced some radio-adaptation, but was not completely protective, reflecting the importance of dose rate when assessing both chromosomal damage and a radio-adaptive response.
Interestingly, there was no significant difference observed in mice receiving an acute 10cGy priming dose and those subsequently challenged with 1.5Gy (Fig. 4, AP versus AP-C). If radiation effects were truly synergistic it would be expected that damage from the priming and subsequent challenge doses would be additive. However, in comparison with the expected values, the observed MN-PCE values were reduced by 69% in the acute AP-C group, and by 42% in the SAP-C group (Table 2), suggesting that although a sub-acute priming dose may be more efficient at inducing repair after low doses, an acute priming dose may be more effective in reducing adverse effects of radiation after challenge doses.
An adaptive response, in terms of MN-PCE, was clearly observed following both acute and sub-acute priming doses. In addition, it was demonstrated that a sub-acute, absorbed dose of 10 cGy resulted in no distinguishable chromosomal damage, whereas the acute 10cGy dose resulted in significantly greater number of chromosome breaks (Fig. 3) when compared to either the controls or sub-acute groups. These data are in concordance with a previous study which evaluated the ability of the Chernobyl environment (dose rate = 2.3 × 10−3 cGy min−1) to induce point mutations in Big Blue® mice (Wickliffe et al. 2003), a sensitive model for detecting point mutations (Winegar et al. 1994). Mice were exposed to the Chernobyl environment for 90 days, accumulating a total dose of 3 Gy (the same total absorbed gamma dose as in Winegar et al. 1994). In contrast, no significant increases in point mutations were observed in comparison with controls and the mutational spectra between exposed and reference animals were the same. A similar study that assessed MN frequencies following exposure of a native species of vole (Myodes glareolus, formerly, Clethrionomys glareolus) to environmental levels of radiation at Chernobyl reported no significant differences in exposed versus reference animals (Rodgers and Baker 2000); a follow-up study demonstrated that the results were not due to the development of radioresistance from living in the radioactive environment for 14 years (Rodgers et al. 2001) but rather to endogenous mechanisms induced by exposure to low dose rate, low-LET radiation.
To gain insight into the mechanism of the radio-adaptive response, tissues were excised (liver, bone marrow, and skeletal muscle) from mice in order to examine transcriptional responses in genes suspected to play a role in adaptation. Preliminary experiments utilizing archived tissue samples from the study reported herein, have revealed that expression of the radical scavenging gene, SOD1, was significantly elevated in both mice receiving a sub-acute 10cGy dose, (SAP and SAP-C groups); however, expression was not elevated in mice receiving the same dose administered acutely (AP or AP-C groups). The fact that both groups exhibited an adaptive response may be indicative of more than one mechanism involved in radio-adaptation and presents further evidence against the validity of the LNT model, under which one would expect a uniform response to the same total dose, regardless of dose rate. Furthermore, these initial results regarding the role of SOD1 are in congruence with other research, which report cells deficient in SOD, exhibited an adaptive response and that adaptation may involve stimulation of DNA repair mechanisms “priming” the cells to efficiently repair damage incurred during subsequent dosing (Wolff 1996). The addition of 3-aminobenzamide, an inhibitor of poly(ADP-ribose) polymerase, involved in DNA strand break repair, erased any protective effect and further supported the hypothesis that adaptation likely has multiple pathways (Wiencke et al. 1986; Shadley and Wolff 1987; Wolff 1996).
It is tempting to speculate that the manner in which the priming dose is received directs the response upon a subsequent challenge dose. For example, low dose rates, such as those encountered in the Chernobyl environment, may not be of the magnitude required to induce significant transcription of DNA repair genes, but may be high enough to significantly induce transcription of radical scavenging genes, such as SOD1. Alternatively, acute doses may be sufficient to induce elevated DNA repair enzymes, which would remain elevated during the challenge dose; thus, the primary adaptive mechanism would be enhanced DNA repair.
Recent advances in molecular techniques, such as microarrays, have spurred research seeking the mechanism of the adaptive response and are providing new insight into this previous black box. Ding et al. (2005) conducted the first study which evaluated comparative global gene expression in low (2 cGy) and high (4 Gy) exposures. Normal human skin fibroblasts were the model studied, since skin is the first barrier that external radiation encounters. The cDNA microarray hybridizations demonstrated that 2345, out of 7458 genes examined, exhibited significant changes in expression. Of these, the expression of 16 genes were significantly altered after only the low dose exposure (no change after exposure to the high dose), whereas 148 genes were found differentially expressed only after exposure the high dose. Genes responding to only the low dose were of the functional groups involved in cell to cell signaling, signal transduction, and DNA damage responses; those that responded only to the high dose were genes primarily involved in apoptosis and cell proliferation. Overall, a dose-dependent response was observed, indicating that biological responses differ from low and high dose exposures, and furthermore, that linear extrapolation from effects resulting from high doses to expected results for low dose exposures is unjustified.
Numerous studies have reported results to low dose radiation exposures contrary to those expected according to the tenets of the Linear No Threshold (LNT) Model. Data presented herein were generated from experiments designed to assess low dose, low dose rate exposures and are not in agreement with the LNT model, which asserts that any amount of radiation is harmful regardless of dose or rate of administration. Continued investigation into the mechanisms underlying the observed response is ongoing by this research team and is necessary to accurately assess risks associated with of long-term exposures to low dose, low dose-rate radiation. However, based on the data from this study, it appears probable that current radiation risk models overestimate human health risks at low doses and dose-rates of low-LET radiation.
Footnotes
ACKNOWLEDGMENTS
The authors thank Drs. Robert Baker and Ron Chesser for their advice, support and contributions to experimental design and dosimetry. We also thank Adam Brown for assistance in the field and laboratory exposures, and Heather Meeks and Kenneth Griffith for editorial assistance. The International Radioecology Laboratory in Ukraine provided logistical support and facilitated permits and access to the Chernobyl Exclusion Zone. This research was supported by the Office of Science (BER), U.S. Department of Energy, Grant No. DE-FG02-02ER63439.