
Abstract
One of the main challenges to the research on allelopathy is technically the separation of allelopathic effect from competition, and quantitatively, the assessment of the contribution of each component to overall interference. A simple mathematical model is proposed to calculate the contribution of allelopathy and competition to interference. As an example of applying the quantitative model to interference by barley (Hordeum vulgare cv. Triumph), the approach used was an addition of allelopathic effect, by an equivalent amount, to the environment of the test plant (white mustard, Sinapis alba), rather than elimination of competition. Experiments were conducted in glasshouse to determine the magnitude of the contributions of allelopathy and competition to interference by barley. The leachates of living barley roots significantly reduced the total dry weight of white mustard. The model involved the calculation of adjusted densities to an equivalent basis for modelling the contribution of allelopathy and competition to total interference. The results showed that allelopathy contributed 40%, 37% and 43% to interference by barley at 6, 12 and 18 white mustard pot−1. The consistency in magnitude of the calculated contribution of allelopathic effect by barley across various densities of receiver plant suggested that the adjusted equivalent density is effective and that the model is able to assess the contribution of each component of interference regardless of the density of receiver plant.
Keywords
INTRODUCTION
Predicting the outcome of plant interference of crops with weeds has become an important topic in agricultural systems since increasing attention has been placed on whether endogenous ‘crop protection’ can be developed in a strongly-interfering crop species (Romeo and Weidenhamer, 1999; Caamal-Maldonado et al., 2001). Harper (1961, 1964) proposed the term interference to describe changes in the environment of plants which result from the proximity of neighbours. Competition and allelopathy are the two components of interference. Muller (1969) defined competition as a process in which one plant depletes some essential elements for plant growth to a level that is limiting to the growth of a second plant sharing that habitat. Molisch (1937) defined allelopathy as any biochemical interaction among plants of all levels of complexity, including micro-organisms. Naturally, competition is physical interference, while allelopathy is chemical interference (Harper, 1977). The response to competition is usually reduction in plant growth, while the response to allelopathy is characteristically stimulation at low concentration and inhibition as the concentration of allelochemical increases (Rice, 1984; Liu and Lovett, 1989).
Interference can undoubtedly occur in field by proximity to other plants. The growth and development of plants may be influenced by both chemical and physical interference (Harper, 1977; Rice, 1984). In some cases, competition may be the predominant contributor to such interference, and in other cases, allelopathy (Bais et al., 2003) may be the major contributor. A classic example is that the sparseness of vegetation under black walnut (Juglans nigra) represents interference via the allelopathic effect of the tree (Rice, 1984). In most cases, observed interference reflects competition and allelopathy acting together (Rice, 1984, Jennings and Nelson, 2002). The potential use of crop plants with high interference ability, as an alternative means of weed control, has increased the importance of identifying and separating chemical and physical interference (Lockerman and Putnam, 1981). Furthermore, ultimate utilisation of the phenomenon of plant interference must rely on effectively separating competition and allelopathy.
However, separation of allelopathy from other aspects of plant interference remains one of the most challenging tasks in the studies of plant interference (Harper, 1977). It would be an advantage to distinguish the relative magnitudes of the separate contributions to total interference so that the strategy of applying crop allelopathy to weed control can be evaluated. To facilitate the separation of allelopathy from resource competition, Liu and Lovett (1993) reported a number of techniques for measuring allelopathy without competition. They are siphoning bioassay apparatus, hydroponic system, and stair-step apparatus. All these techniques are designed to test the living plant without resource competition. The effects of the leachates from living barley growing in these techniques were confirmed as due to allelopathy, and two allelochemicals, gramine and hordenine, were identified in the leachates of the living barley roots (Liu and Lovett, 1993). In our previous reports (An et al., 1996, Liu et al., 2003), modelling methodologies were developed to assist with the study of allelopathy. In this paper, a simple model is proposed to quantify the relative contribution of allelopathy to interference against white mustard by barley. In order to determine the magnitude of allelopathy in terms of its contribution to inference, the approach used is an addition of allelopathic effects to the test plants without introduction of any artificial barrier to plant growth. The interference by barley was assessed and the relative contributions of allelopathy and competition to overall interference were modelled.
MATERIALS AND METHODS
A. Modelling Approach to Separating Allelopathy Contribution from Interference
In a two species mixture, the yield of i-species can be generally expressed as

where wi is the yield of i-species in the presence of j-species, and wio is the yield of i-species in the absence of j-species. fij(ρj) is the effect of j-species on i-species and is greater than, equal to, or less than unity, if the interference of j-species exhibits stimulation, no effect, or inhibition to i-species, respectively. Thus, the magnitude of interference (I) by j-species on i-species, Rij,I, can be calculated as

Combining Eqs (1) and (2) gives

Let wi,C and wi,A be the yield of i-species in the presence of competitive (C) and allelopathic (A) effects of j-species, respectively. The observations from the techniques (Schreiber, 1967; Eagles, 1972; Snaydon, 1979; Liu and Lovett, 1993) reported for separation of allelopathy or competition from interference can be readily applied to this approach. The magnitude of the allelopathic effect (Rij,A) and competitive effect (Rij,C) of j-species on i-species can be respectively calculated by


As interference is a plant density related phenomenon, if the general ability to interfere by one species is to be assessed, the density effect should be considered and adjustment made on an equivalent density basis, whereupon the magnitude of interference can be effectively compared between species. It is proposed that the yield of i-species at a density of m plants per unit area measured in a mixture with j-species at a density of n plants per unit area (wij), may be adjusted by

where Wio and Wij are the yields per plant of i-species in monoculture at density of m plants per unit area, and that in mixture with j-species at density of n plants per unit area, respectively. Figure 1 illustrates the concept. When the i-species coexists at its density less than j-species (m/n < 1), the yield of i-species per plant, wij is adjusted to a bigger value as the yield resulting from the ‘battle’ against greater numbers of competitors, while when the i-species coexists at density higher than j-species (m/n > 1), the yield is adjusted to a smaller value as resulting from the ‘battle’ against fewer number of competitors.

The illustration of yields adjusted (wij) on an equivalent density basis at various plant densities in mixture. The plant yield of i-species in the absence of j-species, (Wio) is set to 1.0, while that in the presence of j-species, (Wij) varies up to 1.0.
Liu (1991) obtained an equation for calculation of total interference (RmI) by barley on white mustard as
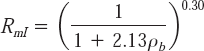
where ρb is the density of barley. As the experiments reported in this paper were conducted under the same conditions as Liu (1991), Equation (7) was used to predict the interference as a reference by comparison to the interference calculated by the current model.
B. Experiments
Examples of applying the approach were illustrated by using the experiments conducted in April 1989 in glasshouse at University of New England, Armidale (Lat. 30°31′S, Long. 150°40′E). A mixture of soil and sand in a 1:1 ratio was used to fill draining-free plastic pots (15cm in diameter). Each pot received 100–150 ml of complete Hoagland's solution (Hoagland and Arnon, 1950) each week and was supplemented by quantities of water every day. As plants were growing up, the distances between pots were increased up to 60 cm for elimination of shading between pots.
Experiment I The experiment was designed to investigate the allelopathic effect of barley leachates, equivalent to 12 plants pot−1, on white mustard (Sinapis alba) in monoculture at three densities, namely 1, 12, and 24 pot−1. These three densities not only provided a wide range, but also had an average density of about 12 plants pot−1, which was close to the donor density (12 barley) tested.
Split-plot designs were used with four replications. The main-plot factors were three densities, namely 1 plant, 12 plants, 24 plants pot−1. The sub-plot factors were treatments with barley leachates (see ‘C. Leachates’) and without leachates. The resultant treatments were i) main-plot factor 1: one white mustard in monoculture (1M) and one white mustard in monoculture plus leachates (1M+L); ii) main-plot factor 2: twelve white mustard in monoculture (12M) and 12 white mustard in monoculture plus leachates (12M+L); and iii) main-plot factor 3: twenty four white mustard in monoculture (24M) and 24 white mustard in monoculture plus leachates (24M+L).
Experiment II To assess interference of barley at the test density of 12 plants pot−1, replacement of the two species to achieve a total of 24 plants was used. The treatment was based on a modified replacement series. In each of the treatments, a monoculture of white mustard, and the second combination (totalling 24 plants) plus leachates were added. The combinations of the two species in the modified replacement design provided an insight into the effect of frequency at constant density. Thus, it enhanced the assessment of interference at the tested density. Further, to assess the magnitude of contributions of competition and allelopathy to the interference by barley, an equivalent amount of allelopathic effect to that involved in the interference was added, not eliminated.
Split-plot designs were used with four replications. The main-plot factors were three mixtures and the sub-plot factors were that without interference, with interference, and with interference plus allelopathic effects. The treatments were i) main-plot factor 1: six white mustard in monoculture (6M), 6 white mustard with 18 barley in mixture (6M+18B) and 6 white mustard with 18 barley in mixture plus leachates (6M+18B+L); ii) main-plot factor 2: twelve white mustard in monoculture (12M), 12 white mustard with 12 barley in mixture (12M+12B) and 12 white mustard with 12 barley in mixture plus leachates (12M+12B+L); iii) main-plot factor 3: eighteen white mustard in monoculture (18M), 18 white mustard with 6 barley in mixture (18M+6B) and 18 white mustard with 6 barley in mixture plus leachates (18M+6B+L).
White mustard in both Experiment I and Experiment II were harvested 5 weeks after sowing. The harvested material was separated into leaf, stem and root components. Leaf area was determined using an electronic planimeter (Paton, Stepney, Australia). Dry weight of leaf, stem and root were obtained after being placed in an oven at 65°C for 48 hours.
Leachates Barley was grown in an incubation tray, as described by Liu and Lovett (1993), with density of 0.3 barley seeds cm−2. Barley was sown one week earlier than Experiment I and II, under the same conditions of both experiments. Barley leachates were collected on day 1 and day 2 of each week, on day 3 one litre of Hoagland's solution was applied to the tray. On the day of leaching, three litres of water were passed through the top and collected from the bottom of the tray twice. One hundred ml of leachates was applied to each appropriate pot. The leachates were equivalent to the leachates in 12 barley plants pot−1.
RESULTS
The leachates of barley roots significantly reduced (P < 0.01) the total dry weight per white mustard in the monoculture experiment at each density (Table 1). The allelopathic effects of barley on white mustard, expressed as RmL, were −0.17, −0.16 and −0.14 in the densities of 1, 12, and 24 white mustard pot−1, respectively. The yield components, that is, leaf dry weight, stem dry weight and root dry weight were also affected, but reduction in stem dry weight, was more pronounced than reduction in leaf dry weight or root dry weight. The average of RmL, −0.16, would be due to allelopathic effects by barley.
Allelopathic Effects of Barley Root Leachates on the Growth of White Mustard

Comparison is only applicable within each column. Different letters denote a significance level of 0.01. RmL is allelopathic effect of barley leachates on white mustard.
Interference by barley significantly depressed the leaf area and dry weight of white mustard (Figures 2 and 3). The leachates significantly exhibited a further reduction of leaf area and dry matter. This further reduction was attributable to allelopathic effects by barley.

Effects of barley with and without additional leachates on leaf area of white mustard.

Effect of barley with and without leachates on total dry weight of white mustard.
The total dry weight of white mustard was separated into the yield components (Table 2). Interference (competition and allelopathy) or interference plus leachates, significantly depressed leaf dry weight, stem dry weight and root dry weight of white mustard (P < 0.05), except in the case of leaf dry weight between the treatments ‘6M+18B’ and ‘6M+18B+L’.
Relative Contributions of Competition and Allelopathy to Interference by Barley on White Mustard

Comparison is only applicable within each column. Different letters denote a significance level of 0.01. The symbols of m, n, W and w are defined in equation (6).
The assessment of interference by barley and the combination of interference with allelopathic effect by barley, and the estimations of the contribution to interference, on an equivalent density basis, are shown in Table 2. The averaged relative interference as assessed by the data obtained from the current experiments was −0.56, which was reasonably similar to the value (−0.61) predicted by the model of equation (7). With the calculations based on equivalent density, allelopathy contributed 40%, 37% and 43% of the interference by barley, while competition by barley contributed to the 60%, 63% and 57% of interference at the densities of 6, 12 and 18 white mustard pot−1, respectively (Table 2).
DISCUSSION
Coble and Ritter (1978) assessed the contribution of competition and allelopathy to the total interference by Polygonum pensylvanicum on Glycine max and found that yield reduction of G. max linearly increased as the density of the weed increased. However, such ‘additive interference’ was totally attributable to competitive interference since no allelopathic effects of P. pensylvanicum were observed in greenhouse studies using a stair-step system recycling nutrient solution. The results of the current experiments confirmed the hypothesis that allelopathy is operative as one component of barley interference. In Experiment I the competitive interference by barley was eliminated and 16% reduction in dry weight of white mustard by leachates was contributed by barley allelopathy. It should be noticed that the relative allelopathic effects of barley on white mustard slightly declined from the magnitude of 0.17 at 1 white mustard to 0.14 at 24 white mustard (Table 1); weak evidence supporting the density-dependent phytotoxicity theory of Weidenhamer et al. (1989). In Experiment II, the further reduction of white mustard in mixture treated with leachates was similarly due to allelopathy, but the reduction attributable to allelopathy was −0.14, −0.22 and −0.32 at the densities of 6, 12 and 18 white mustard, respectively. The contributions of competition and allelopathy to the interference by barley were averaged as 60% and 40%, respectively.
Bell and Koeppe (1972) estimated the relative contributions of allelopathy and competition to interference by giant foxtail (Setaria faberii) and reported that allelopathy contributed to 35% reduction of corn yield in a total 90% reduction by interference (competition and allelopathy). Walters and Gilmore (1976) reported that 19%–69% was attributable to allelopathy in 28%–95% of interference by fescue (Liquidambar styraciflua) with sweet-gum. These two reports had in common a comparison between the single-pot experiment where donor species were grown around the edge of pots with receiver species grown in the centre, and the stair-step experiment, where competition was eliminated. The methodology in such comparisons may be open to question as to the following points. Firstly, since the single-pot experiment had different water supply conditions, compared to the stair-step which received recycling solution daily, the allelopathic or competitive effects may be different. Therefore, the estimation of the relative contribution of the two components to the total interference may not be valid, due to their being conducted under different circumstances. Secondly, interference is a density-dependent phenomenon (Weidenhamer et al., 1989). If allelopathy contributed to interference in the single-pot, the number of donor plants in the stair-step should be equivalent to the number of donor plants in the single pot. It is clear that the authors neglected this factor. In addition, interference is a time-course dependent phenomenon (Liu, 1991). Plants in both the single-pot and stair-step experiments should have had a similar age. It is clear that, taking Walters and Gilmore (1976) for example, fescue growing for two months in the stair-step was not comparable to that growing for four months in single-pot, in terms of relative contribution to allelopathic effects.
In the current experiment, keeping in mind such limitations as above, the number of barley, age of the plant and conditions of the experiment were taken into account in the design and the modelling. Therefore, the relative magnitude of the two components of interference by barley would reasonably reflect the two contributions.
Since it is impossible to eliminate allelopathic effects when any attempt is made to assess contributions of the two components of interference, the approach of adding an equivalent amount of leachates to the system was the preliminary solution to this problem. In order to improve the assessment, it is important to ensure that such equivalent addition is as close as possible to the actual amounts involved in allelopathy. As an average density of donor plants was selected and the variation of allelopathic effects by interaction between density and frequency was also considered, the addition of leachates and assessment of their contribution would be reasonably valid.
The method of Weidenhamer et al. (1989) was designed to determine allelopathic effects as dependent on the density of target plants. They used the classic log-log plot of the response relationship to demonstrate whether allelopathy was involved in the interference. The magnitude of the contribution from allelopathy was unable to be measured in their method. The herein model reported attempted to quantify the allelopathic effect and may be considered as one step forward in the separation of allelopathy from competition in studies on plant interference. However, the method has a number of limitations. First, the equation (6) is only one of many such ways for adjusting the plant yield and not necessarily the best. In particular, the power m/n may be further modified by a constant which is greater or less than a unit. Second, quantitatively assessing the contributions is valid if there are no autotoxins in either species.
One of the main challenges in research on allelopathy is the separation of allelopathic effect from competition (Jensen et al., 2001) and quantitatively determining the magnitude of the contribution to the interference by each component. Although many attempts (Nilsson, 1994; Mallik and Prescott, 2001) have been made to address the issues, such problems remain unsolved in full. Nevertheless, the mechanisms of interference and the contributions of each component must be fully understood before any strategy of crop protection can be effectively developed.