
Abstract
A mathematical model is presented to estimate the effects of phytochemicals on seed germination. According to the model, phytochemicals tend to prevent germination at low seed densities. The model predicts that at high seed densities they may increase the probability of seed germination and the number of germinating seeds. Hence, the effects are reminiscent of the density-dependent effects of allelochemicals on plant growth, but the involved variables are germination probability and seedling number. The results imply that it should be possible to bypass inhibitory effects of allelopathy in certain agricultural practices and to increase the efficiency of nature conservation in several plant communities.
INTRODUCTION
Chemical interference between plants is known to alter the relationship between plant density and growth of plants (Weidenhamer et al. 1989; Sinkkonen 2001). If phytotoxic compounds are homogenously present in soil, plant growth may be totally inhibited at low plant densities, and individual plant biomass may be highest at intermediate plant densities. Plants may grow more than control plants at high densities, but the effect of chemical interference is negligible at very high densities. Therefore, seedling vigor is not proportional to seedling density if phytotoxins are abundant in soil (Yoda et al. 1963; Weller 1987). It is also known that phytochemicals often change the germination percentage of seeds (Williams and Hoagland 1982; Wardle et al. 1992; Chiapusio et al. 1997), and that seed density and solution volume significantly affect seed germination and early growth of seedlings (Weidenhamer et al. 1987; Bergelson and Perry 1989; Wardle et al. 1991; Crawley 1997). Since chemical interference has been distinguished widely in nature (Romeo 2000; Mallik 2002), it is probable that seeds of many wild and cultivated species are prone to chemical interference while they are germinating.
The response of plants to different doses of toxic phytochemicals is well known (Carballeira et al. 1988; An et al. 1993; Romeo 2000). Usually stimulatory response occurs at low doses. As doses increase, the response gradually reaches total growth inhibition, and plants die. Doses per plant usually decrease when plants live close to each other because plants compete for the same phytochemicals. Therefore, maximal total growth may occur at intermediate or high plant densities while plant growth may be zero at low plant densities (Weidenhamer et al. 1989). The biological response model that is based on enzyme substrate reaction is suitable to describe the effects of density-dependent chemical interference mathematically (An et al. 1993; Sinkkonen 2001, 2003). This model separates the stimulatory and inhibitory attributes of plant's response as the dose per plant changes. In the model, the response (P jtot ) of plants that grow in phytotoxic conditions is:

in which Cjtot is the response of control plants, usually their total weight. Im is the inhibitory attribute at saturating concentration, and KI is the concentration at which I = Im / 2 (An et al. 1993). Sm and KS are the respective parameters of the stimulatory attribute. The constant q controls the shape of the curve and is connected to the number of active sites per enzyme molecule for the substrate (An et al. 1993). In the biological response model by An et al. (1993), the dose that affects a plant changes as phyto-chemical concentration changes. This, in turn, changes the biological response of the plant. In the density-dependent extension of the biological response model, plants are assumed to share phytochemicals equally at every concentration and density studied (Sinkkonen 2001). Therefore, the dose (Xj ) per plant at density j is supposed to be a fraction (or a multiple) of the dose (Xi ) per plant at a known number of individuals (Ni ) per unit area (Sinkkonen 2001). If Ni = 1, phytochemical concentration of the substratum can be taken as the basis of Xi . As a result, Xj can be derived at every density if plant number (Nj ) is known (Xj = NiXi/Nj ). Thus, doses per plant are supposed to be known at a ratio scale of measurement (see Sinkkonen 2001). This makes it possible to mathematically estimate the biological response of plants to phytochemicals at different plant densities. The benefits and limitations of this model are discussed in detail by Sinkkonen (2001). Other alternatives to model dose-response curves are discussed by Schabenberger et al. (1999).
Phytotoxic phenomena are often density-dependent. However, present models describe only the growth of plants (Sinkkonen 2001, 2003). For model seed germination and seedling emergence, these models must be modified. The modification should include the possibility of density-dependent changes in germination probability. It should also estimate how the amount of germinating seeds depends on seed density at different phytochemical concentrations. This way, it is possible to assess the impact of density-dependent chemical interference on the number of emerging seeds.
MATERIALS AND METHODS: Construction of the model
It is assumed that the model of Sinkkonen (2001) is applicable, and that soil phytochemicals change the probability of seed germination. The effect may be stimulatory, or inhibitory, and other factors are supposed to be negligible. Let the germination probability of a viable seed be Cg if soil is free from phytochemicals. Let Xi be the dose per seed at seed density i at a certain phytochemical concentration, let Ni be seed number per unit area at the same density and concentration, and let Nj be seed number per unit area at density j at the same concentration. If Ni = 1, the phytochemical concentration of the substratum can be taken as the basis of Xi . Then, it is possible to estimate the germination probability of an average, viable seed in soil that is not free from phytochemicals:

Pg is the germination probability of an average, viable seed in phytotoxic substratum, and Cg is the same probability in non-toxic substratum. Note that 0 ≤ C ≤ 1. Note also that if Ni .→ ∞, Pg → Cg . Due to the dynamics of the model, Pg may assume theoretical values that are higher than one. In such cases, soil phytochemicals do not limit seed germination. This model assumes that there is not any maximal distance (Dmax ) that limits the redistribution of phytochemicals between seeds at existing seed densities. If Dmax exists, Xj is constant as Dj > Dmax . If seeds are not evenly distributed, equation (2) may be valid in seed patches where Dj ≤ Dmax . To get the number of germinating seeds (PN ) per unit area, Pg must be multiplied with the number of viable seeds per unit area:

Equation (3) calculates how many seeds are germinating in homogenously phytotoxic substratum, if the number of viable seeds per unit area is Nj If Pg ≥ 1, PN = Nj . Then, phytotoxins do not limit germination of viable seeds. In equation (3), it is assumed that other factors do not limit seed germination (see Crawley 1997).
RESULTS: Dynamics of the model
Equation (2) predicts that germination probability is highly dependent on seed density in soil (Figure 1). If almost all seeds germinate in phytotoxin-free soil, phytochemicals tend to diminish germination probability at low seed densities unless phytochemical concentration in soil is very low (Figure 1a.). As germination probability of control seeds declines, the possibility of stimulatory effects increases. When 50 % of phytotoxin-free control seeds tend to germinate, the model predicts that intermediate and high seed densities may result in significantly increased germination probabilities across a wide range of densities (Figure 1b). If germination probability of control seeds is very low, it may increase quickly and drastically as seed density increases (Figure 1c). In nature, these changes are obviously dependent on the characteristics of seeds and phytochemicals involved.
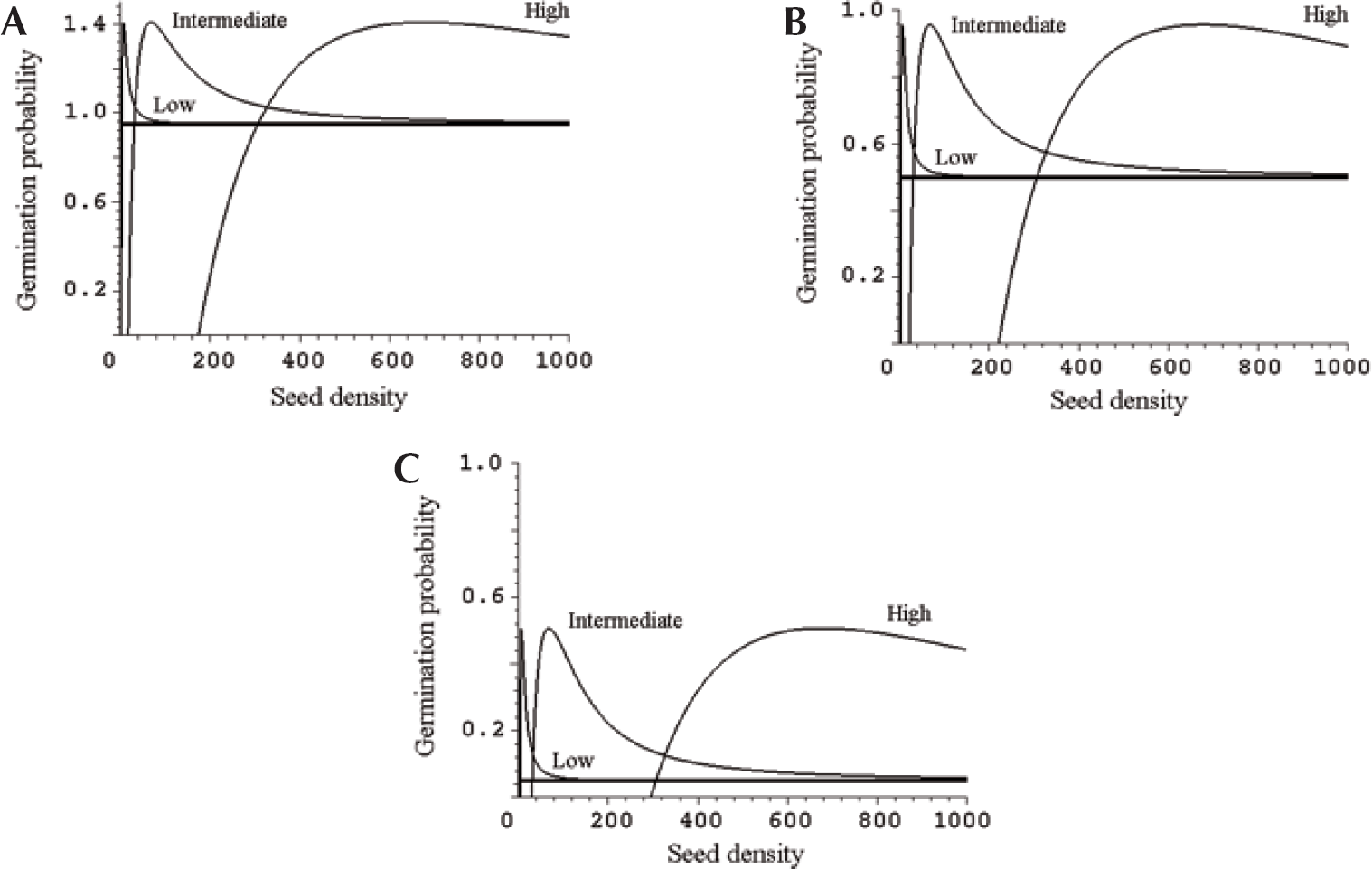
The probability of germination depends on seed density if seeds are affected by phytochemicals in soil. Three initial concentrations of phytochemicals are modeled in each figure: low, intermediate, and high. Control (Xi = 0) is a straight, bold line. Model parameters: Ni = 1, Xi (low) = 0.5, Xi (intermediate) = 5, Xi (high) = 50, Sm = 1, I m = 4, KS = 0.05, KI = 0.3. Figure 1a: Cg = 0.95. Figure 1b: Cg = 0.5. Figure 1c: Cg = 0.05. All units and parameters are arbitrary. Probabilities higher than 1 illustrate model dynamics.
If phytochemical concentration in soil is low, germination probability is predicted to be higher than in the absence of phytochemicals at low seed densities (Figure 1a–1c, lines Low). Thereafter, the probability decreases relatively rapidly to the level of control seeds. In case of mild phytotoxicity, inhibition may not exist at any seed density. At intermediate concentrations, germination probability may be lowered or is zero at low seed densities (Figure 1a–1c, lines Intermediate). As seed density increases, germination probability increases very rapidly, achieves a maximum at intermediate seed densities, and declines thereafter. As phytochemical concentration is high in soil, there may be a relatively high threshold density, below which phytotoxins prevent all germination (Figure 1a–1c, lines High). Beyond that threshold, germination probability may increase steeply. In all cases, extremely high seed densities lead to probabilities that are equal to the germination probability of control seeds.
The model also predicts that density-dependent chemical interference has significant effects on the amount of germinating seeds per unit area (Figure 2). The effects are comparable to changes in germination probability. If practically all seeds tend to germinate in phytotoxin-free soil, phytotoxins usually decrease the amount of germinating seeds at low seed densities, and they can hardly increase seedling numbers at higher seed densities (Figure 2a). The model predicts that if 50 % of viable seeds germinate without phytotoxins, these will significantly increase seedling numbers beyond low plant densities (Figure 2b). If germination is poor with no phytotoxins, the model predicts that these may multiply the amount of seedlings emerged at intermediate and high seed densities (Figure 2c).

Number of germinating seeds depends nonlinearly on seed density if seeds are affected by phytochemicals in soil. Three initial concentrations of phytochemicals are modeled in each figure: low, intermediate, and high. Control (Xi = 0) is a straight, bold line. Model parameters: Ni = 1, Xi (low) = 2.5, Xi (intermediate) = 25, Xi (high) = 100, Sm = 1, Im = 4, KS = 0.05, KI = 0.3. Figure 2a: Cg = 0.95. Figure 2b: Cg = 0.5. Figure 2c: Cg = 0.05. All units and parameters are arbitrary. Seedling numbers illustrate model dynamics if they are higher than the number of viable seeds.
Beyond the case-dependent threshold of zero germination, seedling numbers may increase rapidly if phytochemical concentration in soil is high enough (Figure 2a–2c, lines High). As concentrations in soil decrease, the threshold of zero germination occurs earlier and the increase in the amount of germinating seeds is steeper at low seedling densities (Figure 2a–2c, lines Intermediate). If phytochemical concentration is very low, there may not be total inhibition of germination at any density (Figure 2a–2c, lines Low). Instead, seedling numbers may be increased at relatively low plant densities. The exact shape of the response curve depends always on the seeds and chemicals involved, and soil phytochemical concentration. In theoretical calculations, estimates of stimulatory effects depend on model parameters and they range from significant to negligible. In general, if PN < CgNj , phytochemicals reduce the amount of seedlings emerged. If PN > CgNj , phytochemicals have a stimulatory effect. Since stimulation often occurs as Xj is low, PN may be higher than the number of viable seeds in soil.
DISCUSSION AND CONCLUSIONS
The model developed in this paper implies that density-dependent chemical interference will prevent germination at low seed densities, except at very low allelochemical concentrations. Furthermore, density-dependent chemical interference on germination will tend to increase seedling numbers at intermediate and high seed densities. These results suggest some hypotheses about the operation of allelopathy in certain environments. Since seeds of many cultivated species are sown patchily or into rows where seed density is high, it is probable that allelopathy against seeds of useful plants is often density-dependent in agricultural plant communities. In such cases, it might be possible to bypass the toxic effects of weeds on germinating seeds if seed density is adjusted. Further, natural selection may have minimized harmful allelopathic effects on many weedy species. This may have led those species to produce prodigious numbers of seed.
In nature, allelopathy may prevent germination if viable seeds are of short supply. Further, allelopathy may affect seedling densities wherever viable seeds lie close to each other (Wardle et al. 1991). Traditionally, clumps of seeds have been used to regenerate boreal forest stands. Phenolic compounds are abundant in upper soil layers in boreal and coniferous forests (Northup et al. 1995; Nilsson et al. 1999; Pellissier and Souto 1999; Loponen et al. 2003). Chemical interference between plants is regularly distinguished as a significant ecological factor in those ecosystems in many parts of the world, and several donor species are prevalent there (Kärenlampi and Kauhanen 1972; Nilsson et al. 1998; Wardle et al. 1998; Pellissier and Souto 1999; Mallik 2002). Therefore, density-dependent allelopathy against seed germination is likely to be frequent there (see Nilsson et al. 1998; Nilsson et al. 2000). It may even have modified the evolution of plant species there. For example, allelopathy could be one of the factors behind the high prevalence of berried plants species in those ecosystems (see Kärenlampi and Kauhanen, 1972; Kron et al. 2002). In animal feces, clumps of seeds from fleshy fruits may dilute phytotoxicity and allow seed germination in soil that is rich in plant-born allelochemicals.
The effects of the degradation of phytotoxic plant residues on the growth of many plant species (An et al. 1996, Sinkkonen 2003), and abiotic growth conditions may regulate the concentrations of allelopathic agents in nature (An et al. 2002). These phenomena do not disqualify the significance of density-dependent allelopathy against seeds, they merely complicate the evaluation of the significance of allelopathic interactions in the field. Despite this, the effects of allelopathy on seed germination have special characteristics that probably can be estimated mathematically in several situations if the model presented in this paper is utilized.
Footnotes
ACKNOWLEDGMENTS
Mikko Ojanen and two anonymous reviewers gave constructive comments, and Finnish Cultural Foundation and Wäinö E. Miettinen Foundation funded this work.