
Abstract
YKL-40 is a growth factor for chondrocytes and fibroblasts. The aim was to evaluate YKL-40 expression in the musculoskeletal system during early human development. We studied sections from 15 human embryos [weeks 5.5-8; 7- to 31-mm crown-rump length (CRL)] and 68 fetuses (weeks 9-14; 33- to 105-mm CRL) for YKL-40 protein expression by immunohistochemistry. YKL-40 mRNA expression was evaluated in two human embryos (days 41 and 51). Initially YKL-40 is expressed in all germ layers: ecto-, meso-, and endoderm. YKL-40 mRNA and protein expression are found in tissues of the ecto-, meso-, and endoderm, and YKL-40 protein expression is present during development of cartilage, bone, joints, and muscles. At the cellular level, YKL-40 protein expression is high in tissues characterized by rapid proliferation, marked differentiation, and undergoing morphogenetic changes. Examples of rapid cell proliferation include the chondrogenic inner layer of perichondrium and the osteogenic inner layer of periosteum. Differences in YKL-40 expression during differentiation are found in the chondrogenic and osteogenic cell lineages. The initial shaping of cartilage and bone models and joints is concomitant with a strong outline of YKL-40-positive cells. This indicates that YKL-40 is associated with cell proliferation, differentiation, and tissue morphogenesis during development of the human musculoskeletal system.
YKL-40, a 40-kDa phylogenetically highly conserved heparin- and chitin-binding glycoprotein without chitinase activity, is a member of “mammalian chitinase-like proteins” and has been described in the human (Hakala et al. 1993), chimpanzee, pig (Shackelton et al. 1995), cow, goat (Mohanty et al. 2003), sheep, guinea pig (De Ceuninck et al. 2001), rat, and mouse (Morrison and Leder 1994). Furthermore, the fruit fly Drosophila melanogaster (Kawamura et al. 1999), the mosquito Anopheles gambiae, the zebra fish Danio rerio, the pacific oyster Crassostrea gigas (Badariotti et al. 2006), and the nematode Caenorhabditis elegans have multiple putative YKL-40-like proteins (The National Center for Biotechnology). The human YKL-40 gene is located on chromosome 1q32.1, has a size of 7948 bp and contains 10 exons (Rehli et al. 1997), and the protein's crystal structure is known (Fusetti et al. 2003; Houston et al. 2003). The protein has several names: YKL-40 (Johansen et al. 1992), human cartilage glycoprotein-39 (HC gp39) (Hakala et al. 1993), 38-kDa heparin-binding glycoprotein (Gp38k) (Shackelton et al. 1995), and chitinase-3-like protein 1 (CHI3L1) (Rehli et al. 1997).
In vitro YKL-40 stimulates proliferation of guinea pig chondrocytes, rabbit chondrocytes, and synovial cells and increases proteoglycan synthesis by chondrocytes (De Ceuninck et al. 2001). The growth rates of fibroblasts derived from human osteoarthritic synovium, fetal lung, and adult skin after YKL-40 stimulation are similar to those elicited by insulin-like growth factor-1 (Recklies et al. 2002). YKL-40 initiates mitogen-activated protein (MAP) kinase and PI-3K signaling cascades in fibroblasts, leading to phosphorylation of the extracellular signal-regulated kinase (ERK)-1/2 MAP kinase and protein kinase B (AKT)-mediated signaling cascades, which are associated with control of mitogenesis and cell survival. YKL-40 mRNA expression and protein synthesis by human articular chondrocytes and rat chondrocytes and osteoblasts are induced by tumor necrosis factor (TNF)α and interleukin (IL)-1 and dependent on nuclear factor (NF)-κB activity (Recklies et al. 2005). If TNFα− and IL-1-stimulated human articular chondrocytes and skin fibroblasts are co-stimulated with YKL-40, a reduction of p38 and stress-activated protein kinase (SAPK)/c-Jun N-terminal kinase (JNK) phosphorylation, and metalloproteinase and chemokine IL-8 production is observed (Ling and Recklies 2004). This suppressive effect of YKL-40 is dependent on kinase activity and results in AKT-mediated serine/threonine phosphorylation of the apoptosis signal-regulator kinase, ASK1 (Ling and Recklies 2004). It has recently been shown that YKL-40 binds specifically tocollagen types I, II, and III and modulates the rate of type I collagen fibril formation (Bigg et al. 2006). Furthermore, YKL-40 contributes to chondrocyte differentiation by inducing the transcription factor SOX9 and type II collagen expression, and the induction of SOX9 depends on ERK1/2 and PI-3K activities but not on p38 and JNK MAP kinase (MAPK) (Jacques et al. 2007). These observations suggest that YKL-40 may promote the maintenance or expression of a chondrocytic phenotype and plays a protective role in inflammatory environments, limiting degradation of the extracellular matrix and thereby controlling tissue remodeling.
To elucidate the possible functions of YKL-40, an immunohistochemical study of YKL-40 protein appearance and distribution in the early human musculoskeletal system was performed.
Materials and Methods
Tissue Samples
Fifteen human embryos (6th week, n=3; 7th week, n=6; and 8th week, n=6) and 68 fetuses (9th week, n=9; 10th week, n=11; 11th week, n=9; 12th week, n=14; 13th week, n=12; and 14th week, n=13) were examined. They ranged from 7- to 105-mm crown-rump length (CRL), corresponding to the 6th-14th ovulation weeks. The samples were obtained from spontaneous and legal abortions or ectopic pregnancies. Informed consent was obtained according to the Helsinki Declaration II. Immediately after surgery, the samples were dissected into blocks and fixed for 12-24 hr at 4C in either 10% neutral-buffered formalin, 4% formol-calcium, or Lillie's or Bouin's fixatives. The specimens were dehydrated with graded alcohols, cleared in xylene, and paraffin embedded. Serial sections, 3–5 μm thick, were cut in transverse, sagittal, or horizontal planes and placed on silanized slides.
Antibodies
One mouse monoclonal antibody (MAb) directed against human YKL-40 (MAb 201.F9, 3.8 mg/ml, IgG2bκ, epitope GAWRGTTGHHS corresponding to aa 210-220) and a rabbit polyclonal antibody against human YKL-40 (R667, 0.276 mg/ml, affinity purified on an YKL-40 column) were evaluated on parallel sections of paraffin-embedded tissue using the method described below. The antibodies were independently generated to human YKL-40 purified from serum-free, conditioned media from monolayer cultures of the YKL-40-producing human osteosarcoma cell line MG63, and subsequently purified with high-pressure liquid chromatography (HPLC). The reactivity of R667 has been tested on Western blotting (Volck et al. 1998). Different antibody dilutions and different incubation times were tested, and the mouse MAb 201.F9 was chosen in a concentration of 7.5 μg/ml and the rabbit polyclonal affinity purified antibody 667 in a concentration of 13.8 μg/ml.
Immunohistochemistry
Sections were deparaffinized and rehydrated in xylene followed by a series of graded alcohols, treated with a 0.5% solution of hydrogen peroxide in methanol for 15 min to quench endogenous peroxidase, and rinsed in TRIS buffered saline (TBS, 5 mM Tris-HCl, 146 mM NaCl, pH 7.6). Nonspecific binding was inhibited by incubation for 30 min with blocking buffer (ChemMate antibody diluent S2022; DakoCytomation, Glostrup, Denmark) at room temperature. The sections were incubated overnight at 4C with a monoclonal mouse antibody against human YKL-40 (201.F9; concentration, 7.5 μg/ml) in blocking buffer (ChemMate antibody diluent S2022). The sections were washed with TBS and incubated for 30 min with a peroxidase-labeled polymer conjugated to goat anti-mouse immunoglobulins (DAKO EnVision System/HRP K4007; DakoCytomation). In the experiments with the rabbit polyclonal affinity purified antibody R667 (13.8 μg/ml in blocking buffer), the peroxidase-labeled polymer conjugated to goat anti-rabbit immunoglobulins (DAKO EnVision System/HRP K4011; DakoCytomation) was used. The sections were washed with TBS, followed by incubation for 10 min with 3, 3′-diamino-benzidine chromogen solution. Positive staining was recognized as a brown color. The sections were counterstained with Mayers hematoxylin, dehydrated in graded alcohols followed by xylene, and coverslipped with DPX mounting media. Staining specificity was tested by preincubation of the MAb 201.F9 with purified human YKL-40 for 2 hr at room temperature to block YKL-40 binding sites and reveal possible nonspecific staining to the sections. Positive controls included staining of tissue known to be immuno-reactive for YKL-40. Non-immune mouse IgG1 (X0931; IgG1 concentration, 7.5 μg/ml, DakoCytomation) was used as negative control.
RT-PCR Analysis
Universal human reference RNA (cat. no.: 740000) was purchased from Stratagene (La Jolla, CA). YKL-40 mRNA expression was examined in rib/arm, heart, lung, adrenal gland, kidney, brain stem, spinal cord, and choroid plexus from two human embryos (days 41 and 51). Total RNA was isolated from the embryonic tissues using TRIzol Reagent (Invitrogen; Carlsbad, CA), and cDNA was synthesized with SuperScript II (RNase H−) RT (Invitrogen) according to the manufacturer's instructions. PCR was performed in a total reaction volume of 15 μl containing 20 ng cDNA, 0.1 units of HotStarTaq DNA polymerase (Qiagen; Hilden, Germany), 1× PCR buffer with Tris-Cl, KCl, (NH4)2SO4, 1.5 mM MgCl2 (Qiagen), 0.7 μM of each primer, and 0.2 mM dNTP. The primer sequences were GGACCACAGGCCATCACAGT, CTGCTGGCCGAGGATTCTATG (YKL-40) and GGAAGGTGAAGGTCGGAGTCAA, GATCTCGCTCCTGGAAGATGGT (GAPDH). YKL-40 and GAPDH were amplified using 38 and 24 PCR cycles, respectively. The YKL-40 primer set simultaneously detects at least two splice forms of YKL-40: isoform 1 (containing exon 1-10; GenBank accession number NM 001276) and isoform 2 (in which exon 8 has been spliced out). The identity of PCR products was confirmed by DNA sequencing.
Results
General Pattern of YKL-40 Distribution
The human embryonic and fetal specimens are well preserved and exhibit good to excellent morphology. Immunostaining for YKL-40 results in a clear and well-defined staining pattern, which can be a combination of uniformly or granular cytoplasmic staining and/or membrane-associated staining. This staining pattern is similar irrespective of the fixative used. The specificity and sensitivity of the immunohistochemical method were tested with two different antibodies directed against different epitopes of human YKL-40. Both the mouse MAb 201.F9 and the rabbit polyclonal antibody R667 stained the same cellular structures in the tissue (Figures 1A and 1D). The MAb 201.F9 was selected for further immunohistochemical analysis because of a specific, strong staining and absence of nonspecific background staining (Figure 1A). Control sections with the MAb 201.F9 preabsorbed with YKL-40 antigen showed a lack of or a strongly diminished reactivity (Figure 1B). Control sections, where no antibody or a non-immune mouse IgG was used, (Figure 1C), are blank.
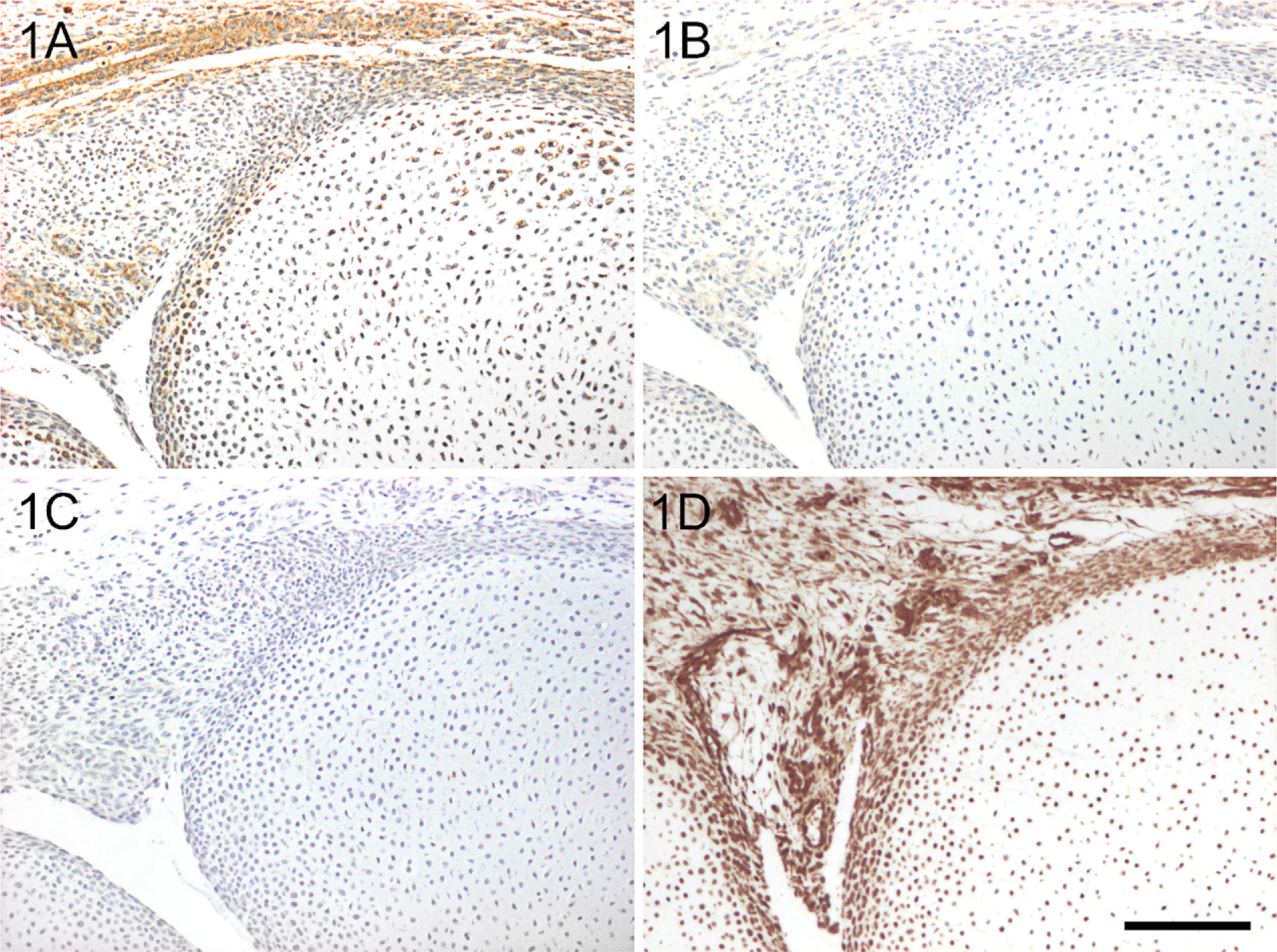
YKL-40 protein expression in a hand from a 12-week-old fetus [crown-rump length (CRL): 90 mm]. Sections were stained with the monoclonal antibody (MAb) 201F9 (
The distribution of YKL-40 immunoreactivity is described in relation to the major developmental stages in bone formation, that is, before chondrification and intramembranous ossification (6th week of embryonic age), during chondrification and initial intramembranous and endochondral ossification (7th-8th week of embryonic age), and finally during chondrocyte differentiation and bone formation (9th-14th week of fetal age). Segmentation of the skeletal blastema and joint formation that represent more longitudinal developmental processes from 6 to 14 weeks are summarized at the end of Results.
YKL-40 mRNA Expression in Embryonic Tissues
YKL-40 mRNA expression was found in rib/arm, arm, heart, lung, adrenal gland, kidney, brain stem, spinal cord, and choroid plexus from 2 human embryos (days 41 and 51) with the strongest expression in kidney and choroid plexus (Figure 2). At least one alternative spliced mRNA, isoform 2, in which exon 8 has been spliced out was detected in all tissues.
YKL-40 Protein Expression in 5- to 6-Week-old Embryos
The earliest embryos studied include three sixth week embryos, 37, 38, and 40 days (CRL: 7-10 mm). They represent the precartilagenous (blastemal) stage, with somite differentiation, limb bud formation, and mesenchymal condensations. The most caudal somites are still present as somites per se, whereas the upper sacral and lumbar somites have differentiated into sclerotomes and dermomyotomes. In the more developed lower cervical and upper thoracic regions, it is possible to distinguish differentiating dermomyotomes, as well as myotomes and ventrolateral somatic epithelial buds with dorsally migrating precursors of limb and abdominal muscles.
Pronounced cellular staining for YKL-40 is observed in all major ecto-, meso-, and endodermal derivatives (Figure 3A). The ectoderm, in particular the thickened ectodermal ring and the entire lining of the gastrointestinal tract, shows strong cytoplasmic YKL-40 protein expression. The notochord and somatic mesoderm also exhibit strong cytoplasmic YKL-40 expression, as well as other mesodermally derived structures such as myotomes and the urogenital (mesonephros) and vascular systems. Neuroectodermal migrating neural crest cells and circulating blood islands from the ventral aortic wall show a distinct YKL-40 expression.
In the early developing musculoskeletal system, the chorda-mesoderm in the tail bud (Figure 3B) and the notochord in the thoracic and abdominal regions (Figures 3C and 4A) have particularly high cytoplasmic granular YKL-40 staining in combination with membrane-associated staining (Figure 3C). The dorsolateral epithelial wall in the distal somites exhibits a distinct cytoplasmic fine granular staining for YKL-40 in contrast to the more weakly reacting ventromedial part (Figure 3B). The more proximal somites are in the process of transformation into sclerotomes and dermomyotomes. The large rounded migrating sclerotomal epithelial-like cells in the ventromedial somite are weakly positive for YKL-40 when they leave the ventromedial wall of the somite (Figure 3B). Shortly thereafter, they become mesenchymal and migrate toward the notochord and the neural tube. In this process, the migrating cells exhibit increasing YKL-40 expression (Figures 4A and 4B) before they settle down as strongly positive YKL-40-stained chondroblast precursors in a position corresponding to the future vertebral chondrogenic center surrounding the notochord (Figures 3C and 4A).
Very high YKL-40 expression is found in the early myoblasts in the dorsomedial lip of the dermomyotomes and in the epithelial cells of the remaining part of the differentiating dermomyotome (Figure 3A). There is also YKL-40 staining in the extracellular space of the dermomyotomes. Separated from the dermomyotomes per se by a slight longitudinal constriction, the ventrolateral epithelial bud forming the hypomere has less YKL-40 expression. Along the lateral border of the bud, YKL-40-positive muscle progenitor cells migrate out through the disappearing basement membrane of the dissociating epithelium (Figure 3D) toward the base of the limb bud, where they show stronger YKL-40 expression. The skeletal blastema in the axial part of the limb bud has only weak YKL-40 expression.
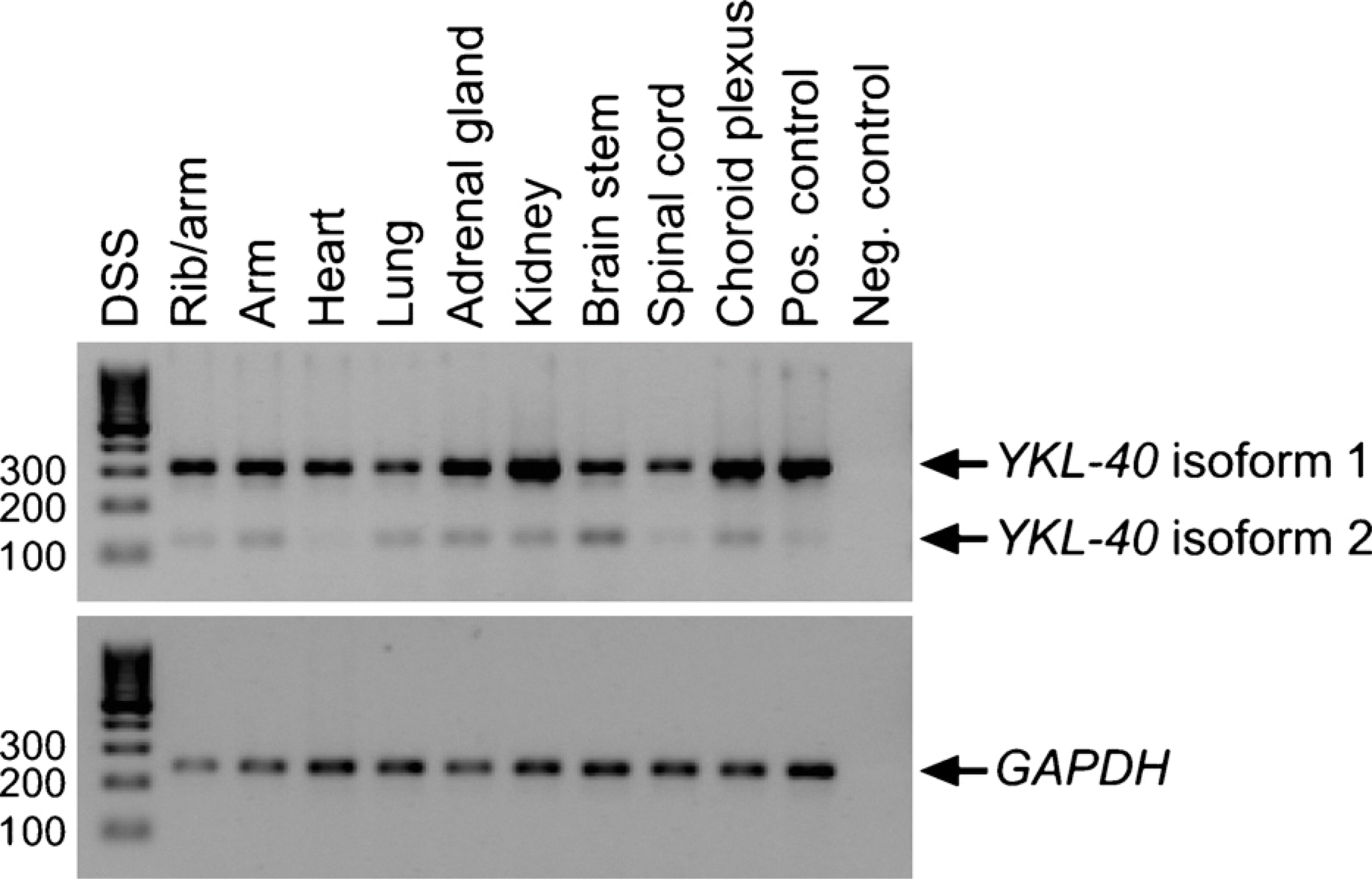
YKL-40 mRNA expression in embryonic tissues. RT-PCR analysis of YKL-40 and the housekeeping gene GAPDH. Total RNA was extracted from tissues dissected from two human embryos 41 and 51 days of age. Top panel: YKL-40 isoform 1 (305 bp) and isoform 2 (122 bp). Bottom panel: GAPDH (240 bp). Lane 1: DNA size standard (DSS). Lanes 2-10: embryonic tissues. Lane 11: positive control (universal human reference RNA). Lane 12: negative control (no template).
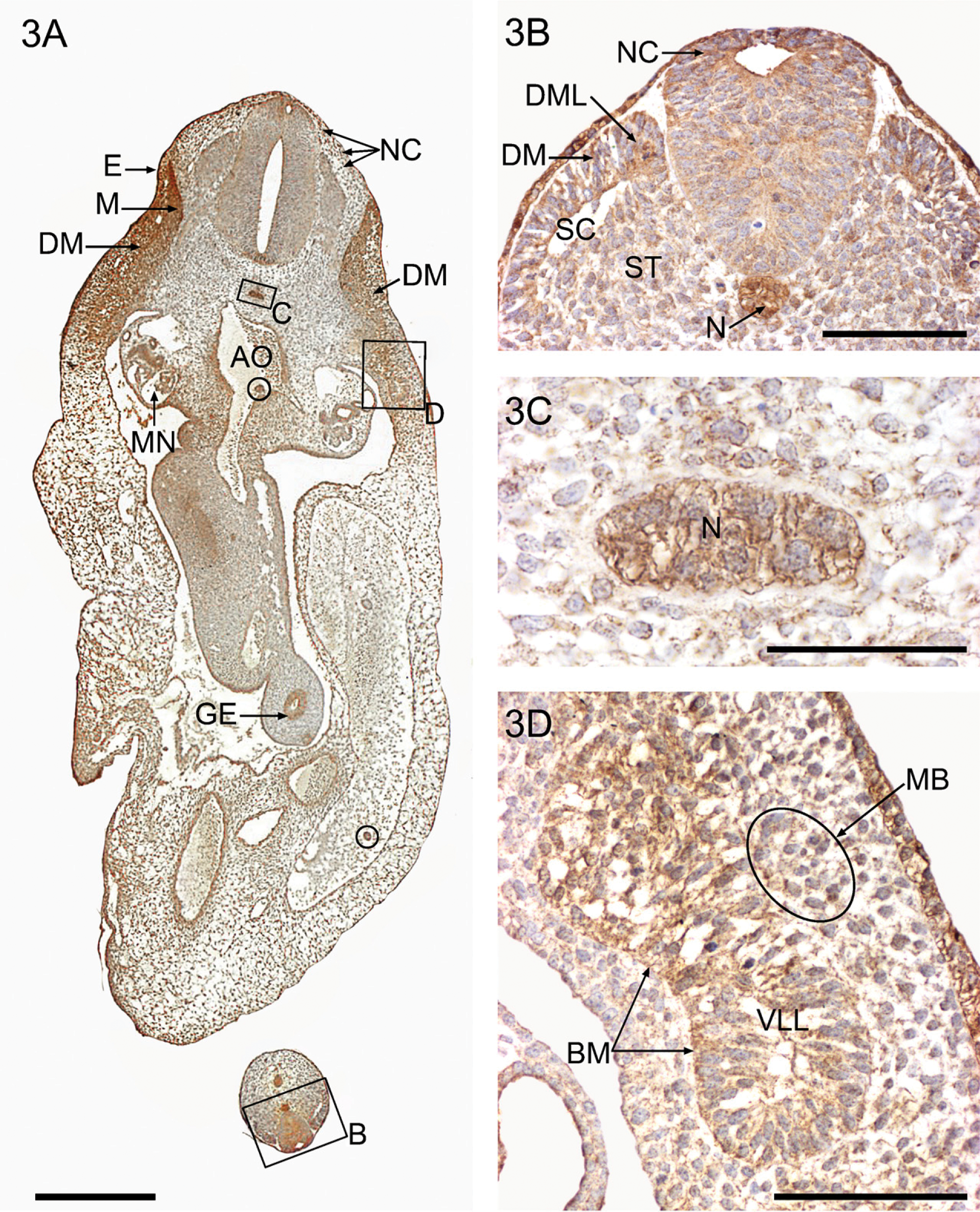
YKL-40 protein expression in a frontal section from a 37-day-old embryo (CRL: 7 mm) (
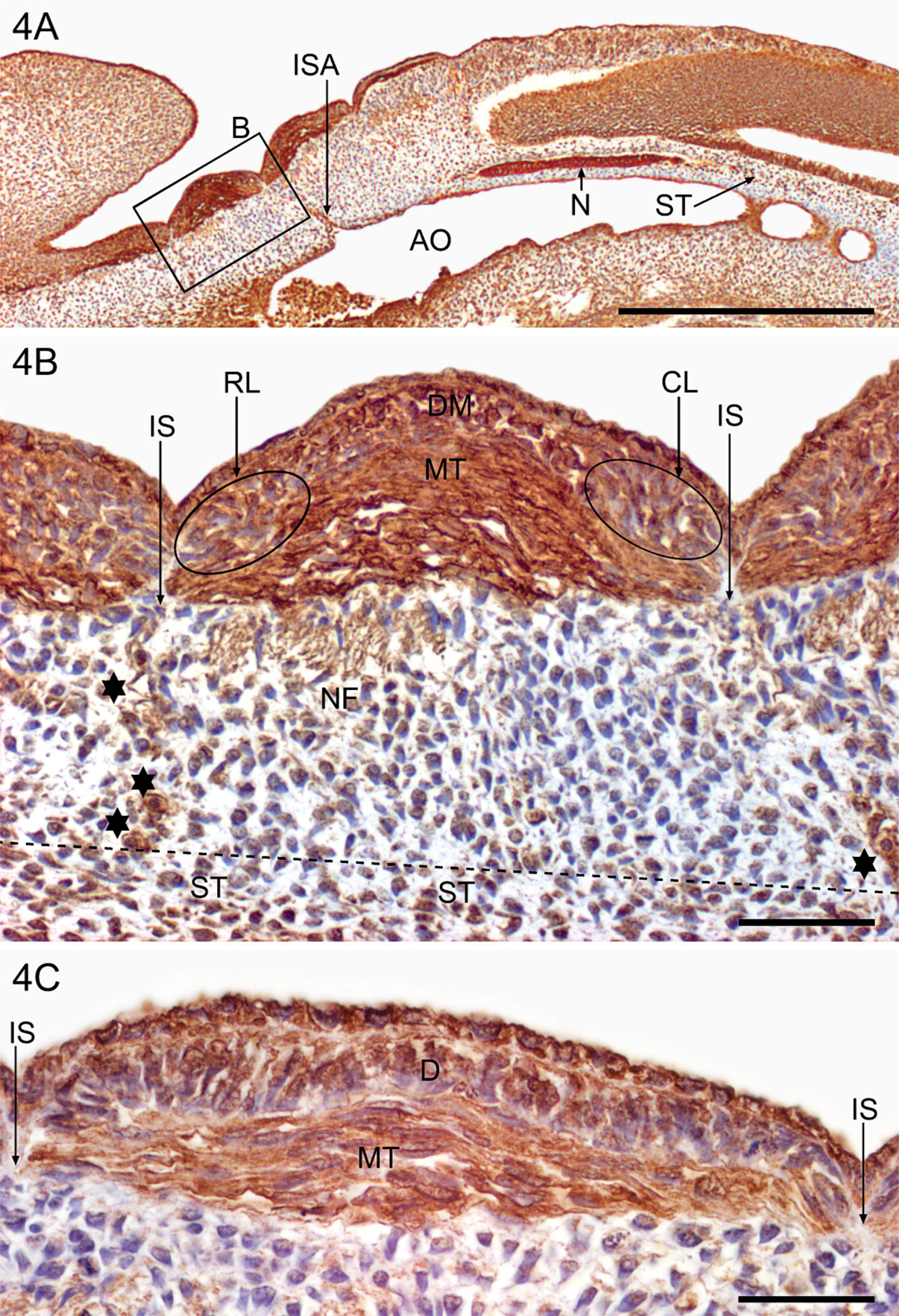
YKL-40 protein expression during early human musculoskeletal development depicted in a sagittal section from a 38-day-old embryo (CRL: 7 mm). Sections were stained with the MAb 201F9. Low power (
Myotome precursor cells translocate from the dermomyotomes to form the underlying myotomes. The myotome cells elongate over the next few days in a direction parallel to the long axis of the embryo and appear as relatively thick spindles with strong YKL-40 expression (Figures 4B and 4C). The YKL-40-negative intersegmental fibrous septa that separate neighboring YKL-40-expressing myotomes extend down toward the strongly YKL-40-positive notochord. YKL-40-positive intersegmental arteries from the aorta occupy a central position within the intersegmental space (Figures 4A and 4B), whereas the segmental nerve fibers are more weakly stained (Figure 4B).
In the pharyngeal arches, the neural crest-derived chondroblasts of the developing chondrogenic centers for Meckel's and Reichert's cartilages show YKL-40 expression among ectomesenchymal cells with no YKL-40 expression (data not shown).
YKL-40 Protein Expression in 7- to 8-Week-old Embryos
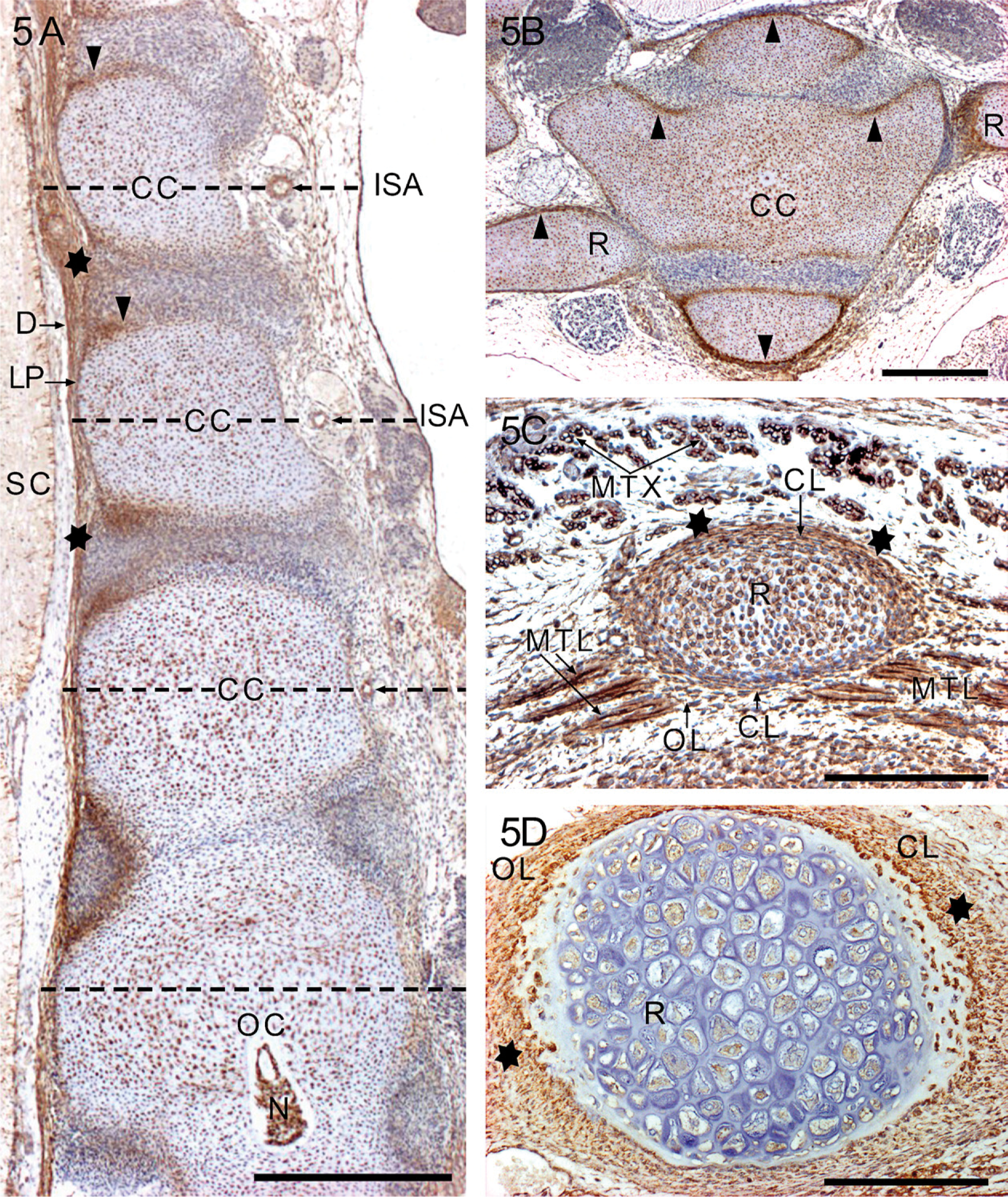
In the seventh week embryos (CRL: 14-20 mm), small-scale embryonic cartilage models, templates of the future bones, are formed. Initially, the models consist of closely packed YKL-40-positive chondroblasts, which are irregular in shape and arrangement, and surrounded by a narrow YKL-40 positive perichondrium. Chondrogenic centers in the form of aggregated chondroblasts first appear in the future vertebral bodies in the upper thoracic region (Figures 5A and 5B) and in the shafts of the long bones of the limbs where chondrification occurs in a proximo-distal sequence. Toward the end of the seventh week, the epiphyses consist of YKL-40-positive chondroblasts.
During the eighth week (CRL: 21-31 mm), most chondroblasts differentiate into young proliferative chondrocytes, most of which have YKL-40 expression. The strongly YKL-40-positive chondroblasts, still present in the primary chondrification centers and the transition zone between peripheral chondroblasts and the chondrogenic deep layer of the perichondrium (Figures 5B and 5C), are responsible for continued interstitial and appositional growth of the former embryonic cartilage models. Furthermore, the extracellular matrix has YKL-40 staining in regions with differential appositional growth (Figure 5B) and in the intervertebral discs (Figure 5A). The notochord still has YKL-40 expression.
The eccentric distribution of the heavily YKL-40-stained perichondrial chondroblasts corresponds well with a differential appositional growth of the cartilage templates (Figure 5C, outer curvature of the rib). Although the demarcation of the perichondrial chondrogenic layer from recently formed peripheral cells of the embryonic cartilage is somewhat indefinite, there is a narrow zone of gentle transition (Figures 5B, 5C and 5D). It is evident that the outer YKL-40-positive layer of chondroblasts of the preformed cartilage template defines the initial shape and position of bones that will subsequently develop in the process of endochondral ossification. Thus, the template provides a temporal plastic framework for bone development.
The hypaxial limb and interlimb muscles, which develop from the ventrolateral epithelial bud, consist at first of growing primary and second of growing multi-nucleated myotubes with centrally located nuclei. Strong cytoplasmic and membrane-associated YKL-40 protein expression is present in the myotubes, particularly in their growing tips from 7 to 14 weeks, whereas fully differentiated muscle cells, that is, muscle fibers with peripherally located nuclei, are YKL-40 negative. The differentiating myotubes and the young muscle fibers get into contact with the outer more fibrous layer of the perichondrium (Figure 5C), but it is not clear in which way the muscles are anchored to the developing bones at later stages.
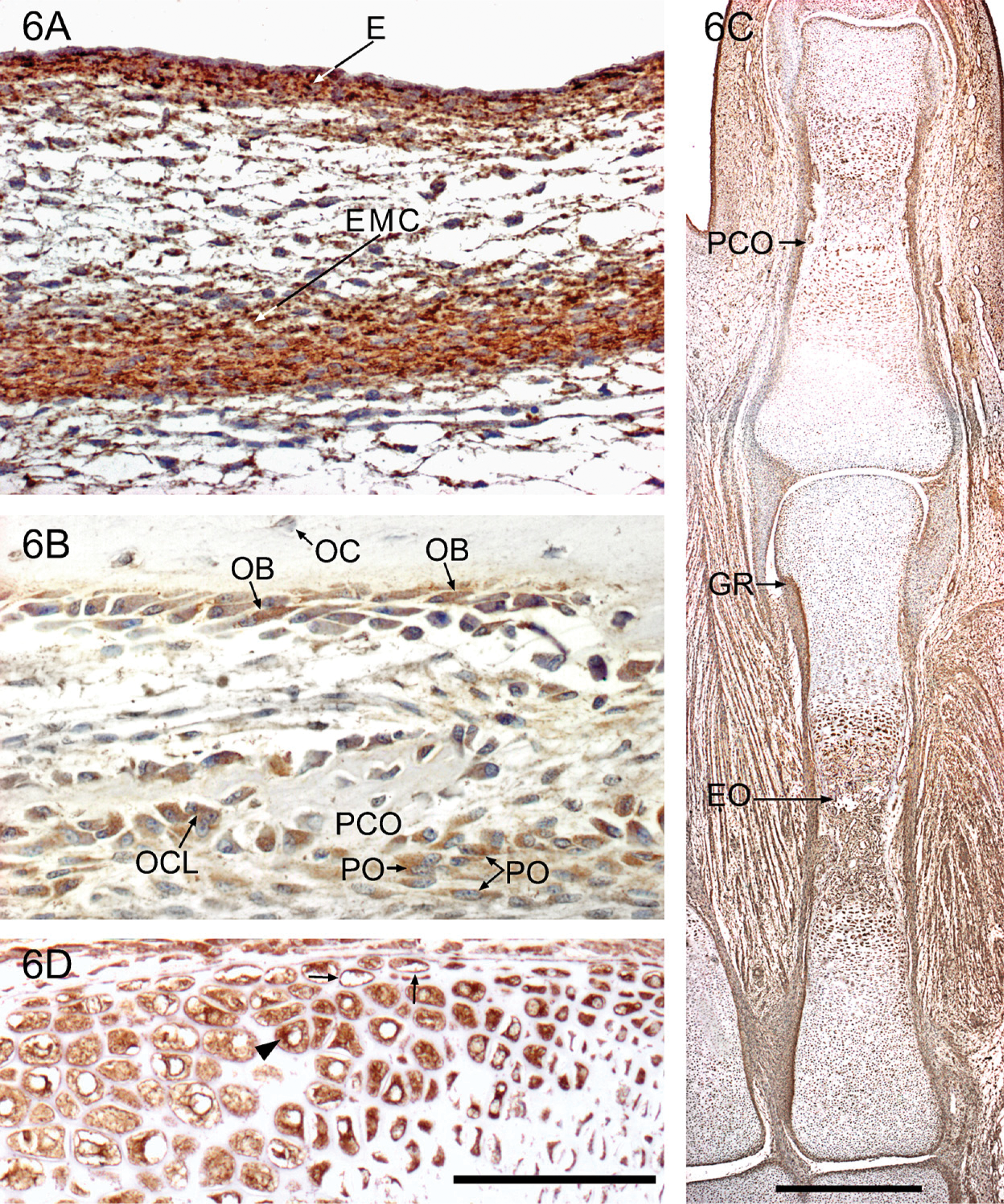
The first sign of bone formation in the human embryo is the appearance of strongly YKL-40-positive ectomesenchymal condensations during the seventh week in areas where the flat bones of the skull and most bones of the viscero-cranium will appear. A condensation corresponding to the future parietal bone is shown in Figure 6A. At the same time, a YKL-40-positive mesenchymal condensation matching the future shaft of the clavicle emerges. During the next few days, YKL-40-positive preosteoblasts and osteoblasts differentiate directly from the condensed mesenchyme in the process of intramembranous ossification. They form small islands of bone, which enclose early developing osteocytes (data not shown). The islands enlarge and fuse and are remodeled by early YKL-40-expressing osteoclasts.
YKL-40 expression in midsagittal (
YKL-40 expression during early human bone formation depicted in sections from a 7-week-old embryo (CRL: 22 mm) (
Early diaphyseal primary ossification is evident during the eighth week in the form of perichondral ossification, which represents a direct osteoblastic differentiation in the perichondrial mesenchyme, that is, a special type of intramembranous ossification. Delicate collars of subperiosteal bone lined by YKL-40-positive preosteoblasts surround the central hypertrophic chondrocytes in the midregion of the shafts of long bones. Genuine endochondral ossification is initiated when vascular osteogenic buds that are also strongly stained for YKL-40 invade the primary ossification centers. Positively stained osteoclasts of hematopoietic origin are contained in the buds (Figure 6B). Perichondral and endochondral ossification from a later stage is shown in Figure 6B.
YKL-40 Protein Expression in 9- to 14-Week-old Fetuses
Between 9 and 12 weeks, the epiphyseal growth plates of the long bones of arm and leg and of “miniature long bones” of hand and foot exhibit the characteristic zones (resting, proliferative, prehypertrophic, and hypertrophic zone), which represent subsequent stages in cartilage differentiation. Resting and proliferative zone chondrocytes are only weakly positive for YKL-40, whereas prehypertrophic and hypertrophic chondrocytes have strong YKL-40 protein expression (Figure 6C). Prehypertrophic and hypertrophic chondrocytes are shown in higher magnification in Figure 6D. Mineralization of cartilage takes place initially in the matrix located between the columns of these YKL-40-positive hypertrophic chondrocytes. The groove of Ranvier—which is the specific structural and functional region where endochondral cartilage fuses with the intramembranous perichondrium—is also strongly stained (Figure 6C, GR). The osteogenic inner layer of the periosteum forms a strongly YKL-40-positive aggregation at depths of the perichondrial ossification groove.
YKL-40 staining is negative when calcification of the matrix is established in the vertebral column after week 11. However, a small number of resting and proliferative chondrocytes within the vertebral growth plate remains positive throughout gestation. The notochord has strong YKL-40 expression, although it is somewhat diminished toward the 12th week (data not shown).
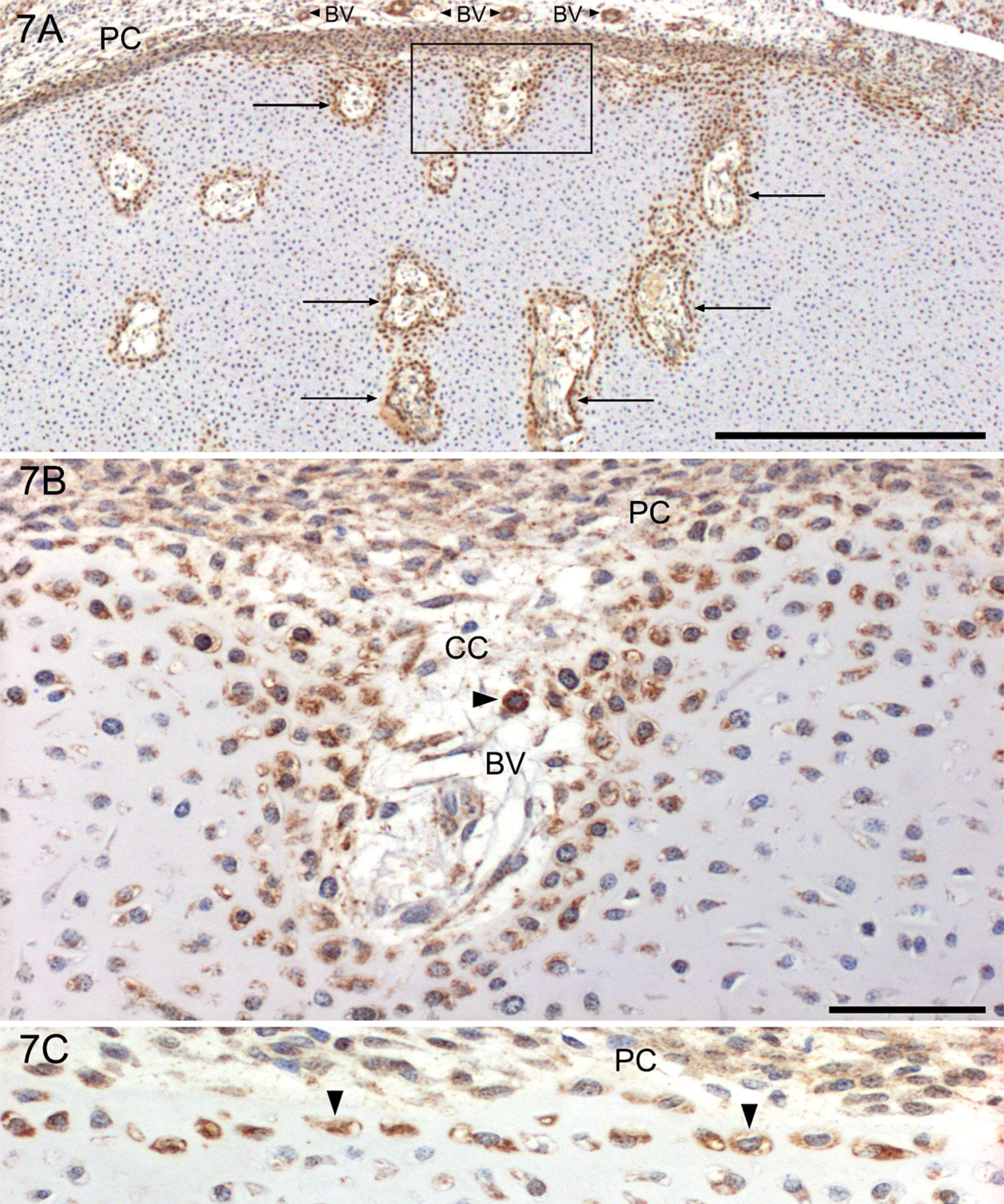
The initial phase of vascularization of the epiphyseal cartilages is seen in fetuses of 75-mm CRL. During week 12 (CRL: 90 mm), secondary centers of ossification start to develop in the epiphyseal cartilage of the long bones. Cartilage canals increase in size and number and invade the epiphyseal cartilage from the deep layer of the perichondrium. They bring in vascular and osteoprogenitor cells to the coming growth plate of the secondary ossification center. The walls of these canals are outlined by YKL-40-positive chondrocytes (Figures 7A and 7B), and the inside is made up of extracellular matrix, YKL-40-positive blood vessels, and other mesenchymal-derived cellular elements (e.g., macrophages, preosteoblasts, and mesenchymal stem cells). The outermost one to two layers of chondrocytes in the part of the epiphyseal cartilage adjacent to the growth plate are strongly YKL-40 positive (Figure 7C). Cartilage canals similar to those found in the epiphyseal cartilages with a similar YKL-40 reaction pattern are present in the cartilaginous vertebral bodies, which are invaded mainly from the posterior surface (data not shown).
Segmentation of the Skeletal Blastema and Joint Development
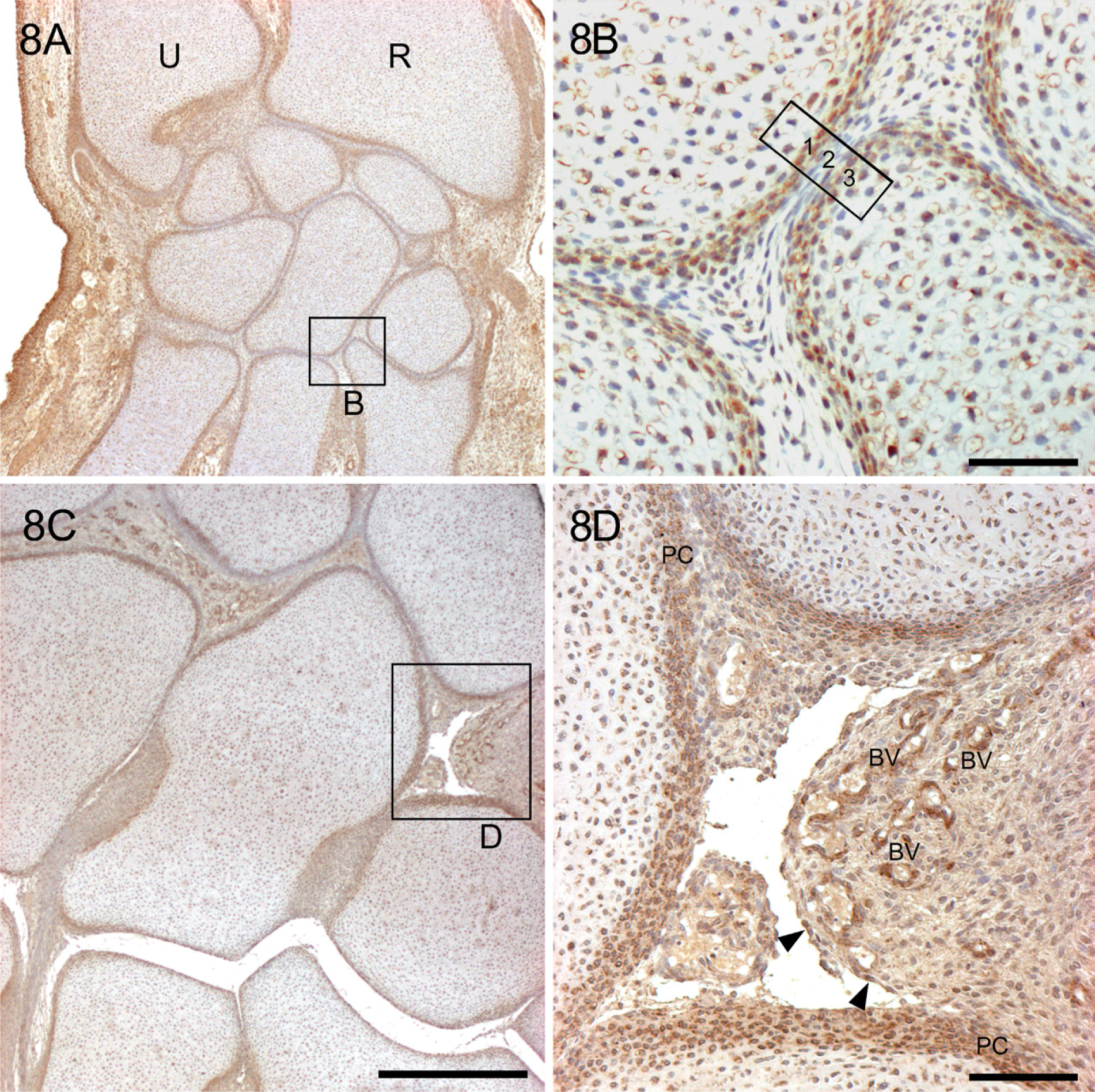
When the miniature cartilage models are outlined by YKL-40-positive chondroblasts within the dense mesenchyme of the skeletal blastema in the sixth week, a homogenous interzone appears in the position of the future joint. Starting from the seventh week, the densely stained chondrogenic inner layer of the perichondrium extends into the loose avascular mesenchymal YKL-40-negative interzone, rendering it three-layered: (1) future articular cartilage, (2) primordial synovial mesenchyme that continues peripherally in a vascular synovial mesenchyme, and (3) future articular cartilage. From the 8th week, cavitation begins in the peripheral part of the interzone in the elbow joint, in the knee joint in the 9th week, and in the radio-carpal and intercarpal joints in the 10th-12th week, from where it spreads toward the center of the interzone. This is shown in Figures 8A and 8B in the cartilaginous models of the carpal bones from a 10-week-old fetus (CRL: 45 mm). The outer fibrous layers of the perichondrium of the adjacent miniature cartilages join and form the basis for the future articular capsule. The cells in the periphery of the cartilage templates condense to form a bilaminar perichondrium, the outer layer of which becomes fibrous, whereas the deep layer develops as a layer of closely packed, rounded chondroblasts, which are responsible for the appositional growth of the templates. These cells have strong YKL-40 expression. However, the demarcation of the chondrogenic layer of the perichondrium from the newly formed cartilage—the chondrocytes—may be quite indefinite, thus rendering a wide zone of gentle transition. When the cartilaginous models are “laid down,” the mesenchyme between adjacent interfaces forms a three-layered interzone (Figure 8B). In the center, the cells become flattened and have no YKL-40 protein expression, whereas at the periphery, they are continuous with the perichondrium of the cartilaginous models and have a strong YKL-40 protein expression. As the templates grow, the central part of the interzone becomes compressed, whereas one or more cavities appear in the peripheral, circumferential part.
The YKL-40-positive deep layer of the perichondrium (PC) penetrates into the distal femoral epiphyseal cartilage in a 12-week-old fetus (CRL: 90 mm) and forms numerous cartilage canals (CC) indicated by arrows. Sections were stained with the MAb 201F9. Framed area in
YKL-40 expression in a hand from a 10-week-old fetus (CRL: 45 mm) before formation of joint cavities (
Joint cavities appear in the hands of 10- to 12-week-old fetuses. Figures 8C and 8D show the section of a hand from a 12-week-old fetus. At that stage, streams of articular chondrocytes with YKL-40 expression seem to enter the forming joint cavities from the highly vascularized synovial mesenchyme, which extends into the joint space from the capsule and has strong YKL-40 expression (Figure 8D). Synoviocytes are particularly strongly reacting, but macrophages and endothelial and vascular smooth muscle cells in the synovial mesenchyme are also strongly positive for the YKL-40 protein. At later stages of joint development, strongly stained synovial mesenchyme is also present. However, articular cartilage chondrocytes on the adjacent epiphyseal surfaces are weakly positive.
Synchondrial joints (e.g., disci intervertebrales) develop within the homogenous interzone, resulting in the formation of a three-layered interzone. The interzone mesenchyme differentiates into two peripheral layers of strongly YKL-40-stained cartilaginous endplates of the intervertebral disc (Figure 5A, arrowheads) and a more weakly stained nucleus pulposus, particularly after disintegration of the notochordal tissue. YKL-40 staining is also distributed in the extracellular matrix of the intervertebral discs. There is strong YKL-40 expression in the posterior annulus fibrosus and the adjoining posterior longitudinal ligament (Figure 5A). Intraarticular ligaments and menisci show moderate YKL-40 expression in early stages of development, whereas collateral ligaments are YKL-40 negative in the outer layers of the capsule. At later stages, YKL-40 is virtually absent from ligaments and menisci. The early differentiating adipocytes of the developing adipose tissue associated with the large joints show high YKL-40 expression.
Discussion
This study reports that YKL-40 protein expression is widely distributed at specific time points in most developing tissues of the musculoskeletal system in human embryonic and early fetal life. We studied YKL-40 protein expression by immunohistochemistry in 15 human embryos and 68 human fetuses between the 6th and 14th ovulation weeks. Our findings suggest that YKL-40 expression is associated with cell proliferation, differentiation, and tissue morphogenesis during development of the human musculoskeletal system. Initially YKL-40 mRNA and protein expression are found in tissues derived from the ecto-, meso-, and endoderm. Studies are presently ongoing regarding YKL-40 mRNA and protein expression during organogenesis and in the early developing human central nervous system.
YKL-40 protein expression is high in cell types characterized by rapid proliferation. In the youngest embryos, sixth ovulation week, a strong cytoplasmic staining for YKL-40 is found in all major ecto-, meso-, and endodermal derivatives. The notochord in the thoracic and abdominal region, prechondroblasts corresponding to the future vertebral chondrogenic center surrounding the notochord and the early myoblasts in the medial lip of the developing myotome, show very strong YKL-40 expression. In the 7- to 8-week-old embryos, the chondroblasts in the primary chondrification center and inner layer of the perichondrium, the chondrogenic layer, have strong YKL-40 expression. These cells are responsible for the continued interstitial and appositional growth of the former embryonic cartilage templates and are modeling small cartilage scale models of the future bones. Other cells with high proliferation rate such as epidermal cells, muscle cells, and the epithelial lining of the gastrointestinal tract have strong YKL-40 protein expression. YKL-40 may act as an autocrine and paracrine growth factor for these cells, because it stimulates proliferation of chondrocytes, synovial cells, and fibroblasts in vitro (De Ceuninck et al. 2001; Recklies et al. 2002).
In addition to a role in cell proliferation, these results suggest that YKL-40 has a role in cell differentiation. High YKL-40 protein expression is found in different cell types during somite differentiation in 6-week-old embryos, particularly in the dermomyotome. Differentiating mesenchymal cells surrounding the notochord and later differentiating prechondroblasts in the vertebral chondrogenic center surrounding the notochord have strong YKL-40 expression. In the long bones of 9- to 14-week-old fetuses, chondrocytes in the prehypertrophic and hypertrophic zone have strong YKL-40 protein expression, whereas chondrocytes in the proliferative and resting zone are weakly positive for YKL-40.
Hypertrophic chondrocytes in the growth plates are thought to undergo apoptosis immediately before ossification and represent the terminal stage of differentiation in the chondrogenic lineage (Forriol and Shapiro 2005). The hypertrophic chondrocyte is a master regulatory cell that prepares the matrix for calcification and vascularization and cause growth by cell expansion (Forriol and Shapiro 2005). Similarly to YKL-40, a high expression of parathyroid hormone-related protein (PTHrP) is found in prehypertrophic chondrocytes in the growth plate, whereas chondrocytes in the hypertrophic zone with high YKL-40 expression have low or no PTHrP expression (Lee et al. 1996; Karaplis and Deckelbaum 1998). PTHrP stimulates proliferation and inhibits differentiation and apoptosis of chondrocytes (Karaplis and Deckelbaum 1998; Strewler 2000). In vitro, PTHrP increases YKL-40 secretion of chondrocytes derived from patients with rheumatoid arthritis but not from patients with osteoarthritis or healthy subjects (Petersson et al. 2006).
Microarray gene analyses of human fetal and adult chondrocytes cultured either under conditions that allow them to preserve their differentiated phenotype or lead to their dedifferentiation showed that YKL-40 is overexpressed >4-fold in dedifferentiated fetal and adult chondrocytes compared with differentiated chondrocytes (Benz et al. 2002; Stokes et al. 2002). Redifferentiation of dedifferentiated chondrocytes also increases YKL-40 expression (Imabayashi et al. 2003). In situ hybridization and immunohistochemical analysis of adult cartilage showed no YKL-40 mRNA or protein expression in chondrocytes in normal adult cartilage but a high expression in chondrocytes in the superficial and middle zone of arthritic cartilage (Volck et al. 1999; Connor et al. 2000; Johansen et al. 2001; Kawasaki et al. 2001; Volck et al. 2001). Cartilage explant cultures or freshly isolated chondrocytes from normal cartilage secrete low levels of YKL-40 during the first days of culture, but after a few days, YKL-40 secretion increases >300-fold (Johansen et al. 2001). Furthermore, stimulation of YKL-40 production by adult articular cartilage can be generated by the trauma of cartilage resection or removal of chondrocytes from their native extracellular matrix environment (Johansen et al. 2001). It has recently been shown that YKL-40 enhances SOX9 and type II collagen expression by primary chondrocytes from neonatal mouse rib cartilage, suggesting that YKL-40 promotes the maintenance or expression of a chondrocytic phenotype (Jacques et al. 2007).
YKL-40 is a major secreted protein of arthritic chondrocytes (Hakala et al. 1993; Johansen et al. 2001), synoviocytes (Nyirkos and Golds 1990; Dasuri et al. 2004), and macrophages in inflamed synovial membrane from arthritic joints (Baeten et al. 2000; Kawasaki et al. 2001; Volck et al. 2001). High YKL-40 concentrations are found in synovial fluid and serum of patients with severe rheumatoid arthritis and osteoarthritis (Johansen et al. 1996, 1999; Conrozier et al. 2000; Garnero et al. 2001; Kawasaki et al. 2001; Volck et al. 2001). It has been suggested that the expression of YKL-40 in injured or degenerated articular cartilage could be related to the initial repair response and increased matrix synthesis seen in arthritic joints (Jacques et al. 2007).
YKL-40 has also been associated with specific differentiation stages in other cell types. YKL-40 mRNA expression in vitro is absent in normal human monocytes but is strongly induced during late stages of differentiation (Krause et al. 1996; Rehli et al. 1997; Renkema et al. 1998). Osteoblasts from adult and fetal bone do not secrete YKL-40 in vitro (Johansen et al. 1992), but YKL-40 mRNA expression is found in end-stage osteoblasts in osteophytic tissue and in primary osteocytes and osteoblasts at sites of endochondral and intramembranous bone formation (Connor et al. 2000). YKL-40 mRNA expression is low to moderate in osteoid-forming and proliferating osteoblasts and undetectable in fully mature osteocytes and osteoclasts (Connor et al. 2000), indicating, in accordance with our studies, a maturation stage-dependent expression of YKL-40 in bone cells. Furthermore, YKL-40 is synthesized in vitro by vascular smooth muscle cells during the time of transition from a proliferating monolayer culture to a non-proliferating differentiated multilayer culture (Millis et al. 1985). YKL-40 secretion continues as the cells reorganize and form multicellular nodules in which cells re-express markers of differentiated vascular smooth muscle cells (Millis et al. 1985; Shackelton et al. 1995; Malinda et al. 1999; Nishikawa and Millis 2003).
YKL-40 may play a role in cell survival. The hypertrophic chondrocytes in the growing fetal long bone advance toward apoptosis, whereas the calcified cartilaginous matrix provides an oriented structure for the osteoblasts to deposit an osteoid matrix after the vessels and osteoprogenitor cells invade the lower hypertrophic cells (Forriol and Shapiro 2005). The high YKL-40 expression in these fetal hypertrophic chondrocytes may protect them from undergoing apoptosis.
Membrane receptors for YKL-40 are not identified. The activation of signal transduction pathways in the cytoplasm of fibroblasts suggests that YKL-40 in these cells interacts with signaling components on the plasma membrane. YKL-40 has proliferative effects on fibroblasts initiating MAPK and PI-3K signaling pathways, leading to phosphorylation of ERK1/2 and AKT (Recklies et al. 2002, 2005; Ling and Recklies, 2004), pathways associated with mitogenesis and cell survival. In this study, we found strong membrane-associated staining for YKL-40 in cells of the notochord and in the growing tips of myotubes, indicating that the cells either have a membrane-associated receptor or a ligand for YKL-40 on the cell membrane.
YKL-40 also binds to collagen types I, II, and III (Bigg et al. 2006), and we found YKL-40 staining of the extracellular matrix in regions of differential appositional growth of the thoracic vertebral columns of 8-week-oldembryos. Other tissues had no YKL-40 staining in the extracellular matrix, suggesting that conformational changes in YKL-40 on binding to ligands in the extracellular matrix may render the epitopes inaccessible to the antibody or that the amount of YKL-40 is too low to be detected by immunohistochemistry.
We suggest that YKL-40 plays a major role in morphogenesis. The skeleton develops from a condensed mesenchymal structure not sharply demarcated, the skeletal blastema, which forms a continuous precartilagenous stage. In most cases, the skeletal blastema undergoes chondrification to form hyaline cartilage models of the bones. These cartilaginous models later ossify by a combination of perichondral and endochondral ossification. Some bones (e.g., the shaft of the clavicle and flat bones of the skull) develop by intramembranous ossification, which represents a direct transformation from condensed mesenchyme to bone (Forriol and Shapiro 2005). YKL-40 protein expression is particularly intense in the early developing human skeletal system in areas with marked morphogenetic properties, for example, during (1) segmentation of the skeletal blastema and formation of the apical epidermal ridge of the embryonic limb bud; (2) chondroblast shaping of the miniature cartilage models and their control of continued growth; (3) formation of joint cavities; and (4) shaping of the miniature bone models from hyaline cartilage models or directly by membranous ossification. Strong YKL-40 expression is found in the notochord, which forms a cellular rod along the longitudinal axis of the embryo, ventral to the central nervous system, and provides position and fate information to all surrounding tissues. The notochord is instrumental in specifying a variety of cell types in forming somites and thus in the transformation of the somites into the axial skeleton and most skeletal muscles (Stemple 2005).
It remains an open question as to the exact functions of YKL-40 in the development of the musculoskeletal system, but YKL-40 is expressed in critical periods. We suggest that YKL-40 plays a role in cell proliferation, differentiation, and survival and in morphogenesis.
Footnotes
Acknowledgments
This work was supported by grants from the Danish Rheumatism Association, Direktør Jens Aage Sørensen og Hustru Edith Ingeborg Sørensens Mindefond, and Vera og Carl Johan Michaelsens legat. The expert technical assistance of Sussi Forchhammer, Department of Cellular and Molecular Medicine, Developmental Unit, The Panum Institute, University of Copenhagen, Copenhagen, Denmark, through all stages of the project is gratefully acknowledged. Keld B. Ottosen, The Panum Institute, is thanked for the final layout of the figures.