
Abstract
Serine proteases and their cognate serpin-class inhibitors are involved in the controlled proteolytic events during follicular development, ovulation, formation, and maintenance of the corpus luteum (CL). In this study, we investigated the expression patterns of prostasin serine protease and protease nexin-1 (PN-1), a serine protease inhibitor also called serpin-E2, in rhesus monkey ovaries during the menstrual cycle and early pregnancy, by using in situ hybridization and immunohistochemistry. Expression of prostasin was localized in oocyte, granulosa cells, and/or theca cells of early antral follicles and antral follicles, with high levels observed in preovulatory follicles. Prostasin was also localized at high levels of abundance in the CL during the menstrual cycle and early pregnancy. During the menstrual cycle, PN-1 was coordinately localized with prostasin in oocytes, granulosa cells, and theca cells of antral follicles and preovulatory follicles and in the CL. In addition, the PN-1 expression level in macaque CL during early pregnancy increased as pregnancy proceeded. We propose that prostasin may be involved in follicular development, ovulation, and CL formation, whereas PN-1 may be present to regulate the proteolysis in these processes.
D
Prostasin serine protease, with a molecular mass of 40 kDa, has a glycosylphosphatidyl inositol membrane anchor and can also be secreted (Yu et al. 1994; Chen et al. 2001b). It is considered a trypsin-like serine protease because it displays arginine amidase activity on substrates at the carboxyl-terminal side of Arg residues (Yu et al. 1994). Prostasin is present at high levels inhuman seminal fluid, urine, and the prostate gland (Yu et al. 1994; Chen et al. 2001a) and at moderate levels in the colon, lung, kidney, pancreas, salivary gland, liver, and bronchi (Yu et al. 1994). It is an in vitro invasion suppressor of prostate cancer and breast cancer (Chen et al. 2001a; Chen and Chai 2002; Takahashi et al. 2003) and a regulator of epithelial sodium channel activity in the kidney (Narikiyo et al. 2002; Iwashita et al. 2003; Tuyen et al. 2005).
A prostasin-binding protein abundant in the seminal vesicles has recently been identified as protease nexin-1 (PN-1), which inhibits the activity of prostasin and can bind to membrane-anchored prostasin (Chen et al. 2004). PN-1, also called serpin-E2, is a serine protease inhibitor belonging to the serpin superfamily. It can modulate the proteolytic activities of several serine proteases, such as thrombin, plasmin, trypsin, uPA, and tPA (Scott et al. 1985; Gettins 2002). Although PN-1 is widely considered to be important in the processes of follicular development, ovulation, and formation of the CL (Hagglund et al. 1996; Kim et al. 2001; Hasan et al. 2002; Bedard et al. 2003; Cao et al. 2004), the expression or function of PN-1 in the rhesus monkey ovary has remained unknown.
In this study, as a first step toward understanding the potential roles of prostasin and PN-1 in the primate ovary, we investigated the distribution patterns of prostasin and PN-1 transcripts and proteins in the rhesus monkey ovary during the menstrual cycle and early pregnancy, by using in situ hybridization and immunohistochemistry.
Materials and Methods
Animals and Tissue Preparation
Animal care and treatment were in agreement with the Guidelines on the Care and Use of Laboratory Animals of the Ethical Committee, State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences. Rhesus monkeys (Macaca mulatta) were housed in the Medical Primate Center, Institute of Medical Biology, Chinese Academy of Medical Sciences as detailed previously (Wang et al. 2001). Adult female rhesus monkeys with a history of regular menstrual cycles and pregnancy were allowed to mate with the males for 2 days from the anticipated time of ovulation. Sperm plug examination, ultrasonography, and monkey chorionic gonadotropin detection in the urine were performed to determine pregnancy. The anticipated day of ovulation was designated as day 0 of pregnancy. On days 6 (n=3), 11 (n=3), 20 (n=3), and 28 (n=3) of the normal menstrual cycle, and on estimated days 12 (n=3), 18 (n=3), and 26 (n=3) of pregnancy, monkeys were laparotomized under ketamine hydrochloride anesthesia. Ovaries were removed, fixed in Bouin solution for 20 hr, and embedded in paraffin.
Probes Synthesis
A human prostasin cDNA fragment (corresponding to bases 799-1072 of L41351 in GenBank) was subcloned from full-length cDNA (Chen et al. 2001b) into the pSP73 vector (Promega; Madison, WI) using the XhoI and BamHI sites. The sense and antisense RNA probes were transcribed with T7 and SP6 RNA polymerases, respectively, using a digoxigenin (DIG) RNA labeling kit (Roche Diagnostics; Indianapolis, IN) following the manufacturer's instructions.
A full-length human PN-1 cDNA was amplified by PCR as described previously (Chen et al. 2004) and cloned into pGEM-T Easy vector (Promega). A SacI-HindIII fragment (corresponding to bases 804-1084 of A03911 in GenBank) was subcloned into the pGEM-3Z vector (Promega). The sense and antisense RNA probes were synthesized with T7 and SP6 RNA polymerases, respectively, using a DIG RNA labeling kit (Roche Diagnostics).
The concentrations of the riboprobes were determined using MetaView image analyzing system version 4.50 (Universal Imaging Corp.; Downingtown, PA) on spot tests following the manufacturer's instructions, with a DIG-labeled RNA control provided in the labeling kit.
In Situ Hybridization
In situhybridization was carried out as described previously (Wang et al. 2004). In brief, successive paraffin sections (5-μm thickness) were deparaffinized extensively and rehydrated in a decreasing series of alcohol solutions. After brief rinsing with 1× PBS twice, the sections were treated with proteinase K solution (at a final concentration of 10 μg/ml in 50 mM EDTA and 100mM Tris, pH 7.5) at 37C for 10 min. The sections were rinsed in 1× PBS, fixed in 4% paraformaldehyde for 15 min, and dehydrated in an increasing series of alcohol solutions. The slides were treated in a prehybridization solution [50% deionized formamide, 10% dextran sulfate, 0.5% SDS, 1× Denhardt, 10 mM Tris, pH 8.0, 10 mM dithiothreitol, 2× SSC (1× SSC is 0.15 M NaCl, 0.015 M sodium citrate), 250 μg/ml salmon sperm DNA] at 50C for 2 hr and hybridized with denatured probes at a final concentration of 400 ng/ml in the prehybridization solution at 50C for 18 hr. After serial washes at 42C in 23 SSC for 30 min, in 1× SSC for 30 min, in 0.1× SSC for 30 min, and at room temperature in buffer I (100 mM Tris, 150 mM NaCl, pH 7.5) for 5 min, the sections were blocked in a blocking solution (1% blocking reagent in buffer I; Roche Diagnostics) for 1 hr. The slides were incubated with anti-DIG-alkaline phosphatase (diluted 1:500 in the blocking solution) at room temperature for 2 hr and further rinsed in buffer I three times for 10 min. Color was developed with nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate (NBT/BCIP; Roche Diagnostics) in buffer II (100 mM Tris, 100mM NaCl, 50 mM MgCl2, pH 9.5). Finally, the slides were rinsed in 95% ethanol for 30 min and mounted for photography. The results were recorded with a SPOT digital camera system (Diagnostic Instruments; Sterling Heights, MI). Sense probes were used on adjacent slides in each experiment as control for background-level staining.
Immunohistochemistry
The anti-human prostasin polyclonal antibody used in this study was described previously (Chen et al. 2001b). A recombinant human PN-1 protein expressed in Escherichia coli as a fusion protein with the schistosomal glutathione-S-transferase was purified as described (Chen et al. 2004) and used as the antigen for raising a rabbit anti-human PN-1 antibody. The antibody was produced by Cocalico Biologicals (Reamstown, PA) under contract. A biotin-streptavidin-peroxidase method (Vectastain SP kit; Vector Laboratories, Burlingame, CA) was used to visualize prostasin and PN-1 immunostaining in rhesus monkey ovary samples as described previously for monkey uterus and placenta (Lin et al. 2006). Briefly, the paraffin-embedded sections (5-μm thickness) were deparaffinized and rehydrated completely, followed by antigen retrieval in citrate acid buffer (10 mM of citrate sodium, 10 mM of citric acid, pH 6.0) in a microwave oven at 92-98C for 15 min. The sections were cooled to room temperature and treated with 3% H2O2 in methanol for 15 min to inactivate endogenous peroxidase. After an incubation in 10% blocking serum in TBST (10 mM Tris-HCl, pH 7.6, and 150 mM NaCl, containing 0.1% Triton X-100) for 30 min, the sections were sequentially immunoblotted with rabbit anti-prostasin or anti-PN-1 antibody (at a dilution of 1:500 in the blocking buffer) for 2 hr, incubated with biotinylated anti-rabbit IgG for 30-60 min, and horseradish peroxidase (HRP)-labeled streptavidin for 1 hr. Washes with 1× PBS were performed between steps. The color reaction was performed with diaminobenzidine kit (Zhongshan; Beijing, China). The sections were counterstained with Harris hematoxylin and mounted for photography. Negative controls were slides treated side-by-side without the primary antibody. In this study, preabsorbed antibodies were not used as negative control.
Statistical Analysis
Signal intensities of prostasin and PN-1 mRNA and proteins detected by in situ hybridization and immunohistochemistry were quantified by computer-aided laser scanning densitometry (Personal Densitometer SI; Molecular Dynamics, Sunnyvale, CA). To make the statistical significance of the quantitative difference credible, three slides from each of three monkeys of each time point were examined, and 30 spots were randomly selected from each compartment on a slide. Values are presented as means ± SEM of each compartment, and the gray level of intercellular substance was considered as background. Statistical analysis was performed using the SPSS for Windows Package, Release 10.0 (SPSS; Chicago, IL). One-way ANOVA was used to determine differences among the three groups (early antral follicles, antral follicles and preovulatory follicles during the menstrual cycle, or CL on D12, D18, and D26 of pregnancy). A t-test was used to determine the difference between the signals in the compartment and its corresponding background (intercellular substance). p<0.05 was considered to be a statistically significant difference. Expression levels at p<0.05 were considered as moderate, and expression levels at p<0.01 were considered as strong or high.
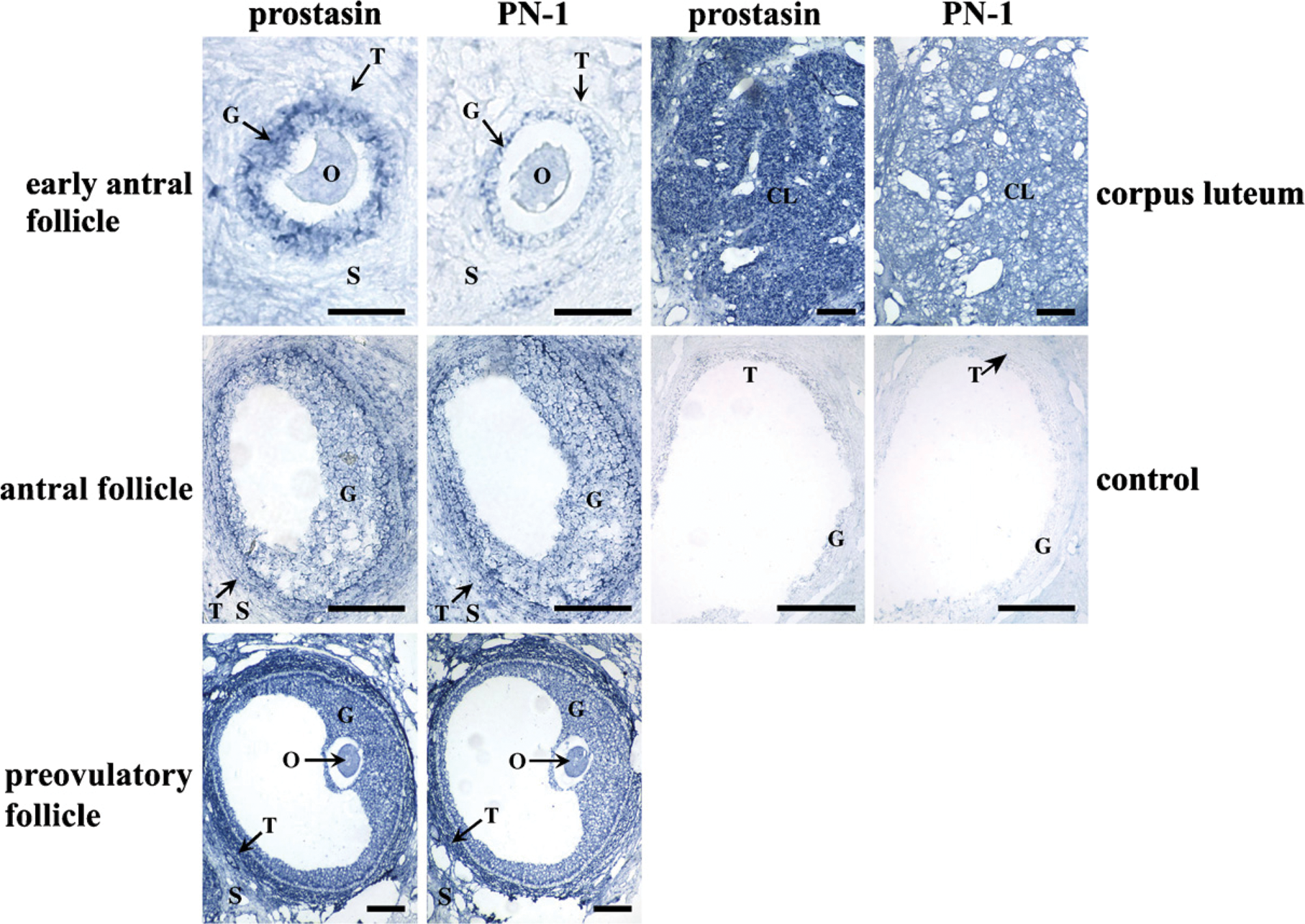
Localization of prostasin and protease nexin-1 (PN-1) mRNA in macaque ovary during menstrual cycle by in situ hybridization. control, control sections hybridized with sense probe; CL, corpus luteum; G, granulosa cells; O, oocyte; S, stroma; T, theca cells. Bar = 200 μm.
Results
Localization of Prostasin and PN-1 mRNA in Macaque Ovarian Tissue During the Menstrual Cycle
In situ hybridization was performed to study the spatiotemporal expression patterns of prostasin and PN-1 mRNA in the cyclic macaque ovary. Prostasin mRNA was concentrated in oocytes and granulosa cells of early antral follicles at high levels (p<0.01; Figure 1; Table 1), whereas signals in theca cells were not different from background staining (p>0.05; Figure 1; Table 1). In the early antral follicles, the PN-1 transcript was expressed in oocytes and granulosa cells at a moderate level (p<0.05; Figure 1; Table 1), and signal in theca cells was the same as the background (p>0.05). In the granulosa cells and theca cells of developing antral follicles, both prostasin and PN-1 were strongly expressed (p<0.01; Figure 1; Table 1). Prostasin and PN-1 were also expressed in the stroma at this stage. In preovulatory follicles, prostasin and PN-1 were localized in oocytes, granulosa cells, theca cells, and the stroma at high levels (p<0.01; Figure 1; Table 1), and the signals at this stage were the strongest compared with those at other stages of folliculogenesis. In the CL, prostasin was expressed in the CL at high levels and in the stroma at moderate levels. A strong PN-1 signal was observed both in the CL and stroma (p<0.01; Figure 1; Table 1). Sense-strand probes for prostasin and PN-1 yielded no specific signal in any tissues studied (Figure 1).
Densitometric quantification of prostasin and PN-1 transcripts in different histological compartments of the macaque ovary during the menstrual cycle
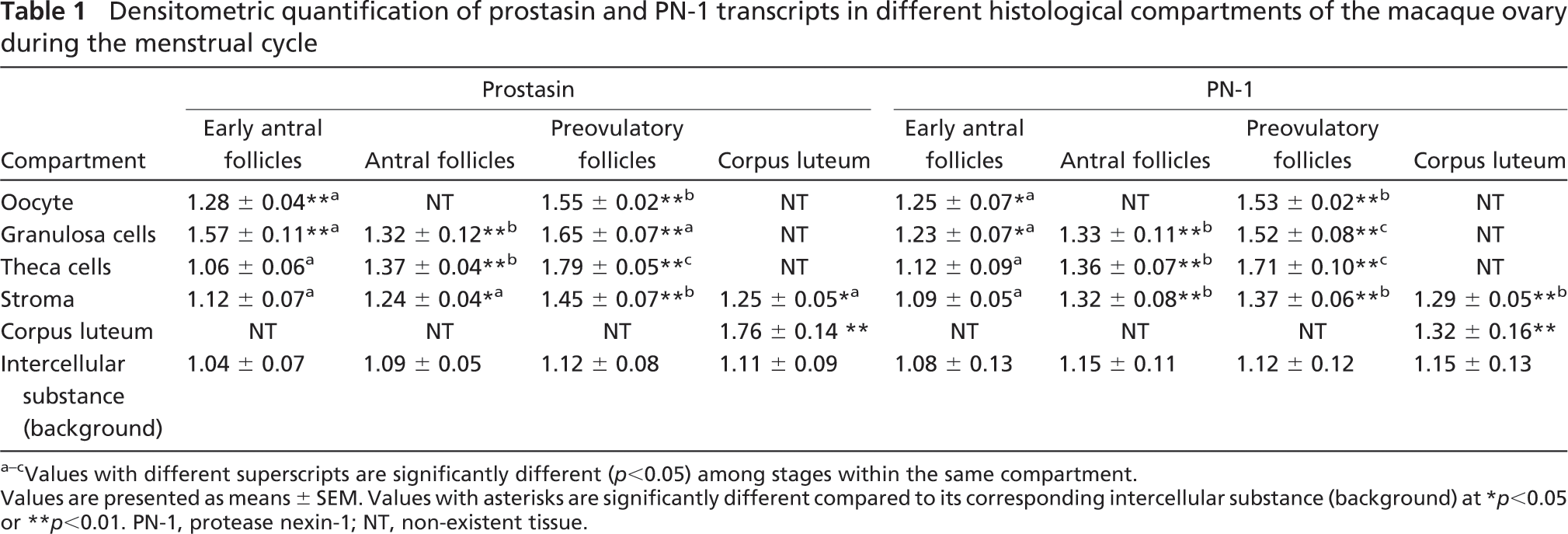
Values with different superscripts are significantly different (p<0.05) among stages within the same compartment.
Values are presented as means ± SEM. Values with asterisks are significantly different compared to its corresponding intercellular substance (background) at ∗ p ∗lt;0.05 or ∗∗ p<0.01. PN-1, protease nexin-1; NT, non-existent tissue.
Immunohistochemical Staining of Prostasin and PN-1 in Rhesus Monkey Ovaries During the Menstrual Cycle
To localize prostasin and PN-1 proteins in the cyclic ovary, immunohistochemical staining was performed. The results showed that the expression levels of prostasin and PN-1 proteins in granulosa cells and theca cells increased from background levels (.>.05) to high levels (p<0.01) along with follicle development from early antral follicles to preovulatory follicles. However, prostasin and PN-1 proteins were highly expressed in oocytes from early antral follicles to preovulatory follicles (p<0.01; Figure 2; Table 2) and also were expressed in the stroma at moderate or high levels. In the CL, prostasin protein was highly expressed in the CL (p<0.01) and moderately expressed in the stroma (p<0.05; Figure 2; Table 2). PN-1 protein was only moderately expressed in the CL. The negative control treated with normal rabbit IgG produced negligible background (Figure 2).
Localization of Prostasin and PN-1 mRNA in Macaque Ovarian Tissue During Early Pregnancy
As shown in Figure 3 and Table 3, relatively strong signals of prostasin transcripts were localized in the CL in the macaque ovary on days 12, 18, and 26 of early pregnancy (p<0.01). Distinct from the expression pattern of prostasin, PN-1 was moderately expressed in the CL on day 12 of pregnancy (p<0.05), strongly expressed on day 18, and peaked on day 26 (p<0.01; Figure 3; Table 3). The intensity of PN-1 mRNA in the CL significantly increased over the three stages of pregnancy (p<0.05; Table 3). No background staining was detected in control tissues hybridized with sense probes (Figure 3).
Immunohistochemical Staining of Prostasin and PN-1 in Rhesus Monkey Ovaries During Early Pregnancy
As shown in Figure 4 and Table 4, high levels of prostasin protein signals were observed in the CL in the monkey ovary on days 12, 18, and 26 of early pregnancy (p<0.01; Figure 4; Table 4). PN-1 protein was expressed in the CL at a relatively low concentration on day 12 of pregnancy (p<0.05; Figure 4; Table 4). The signals of PN-1 peaked on day 18 and kept a high level of expression on day 26 (p<0.01; Figure 4; Table 4). Control sections stained with normal rabbit IgG produced no positive signal (Figure 4).
Discussion
In this study, we reported the expression of prostasin during macaque follicular development and CL formation in the menstrual cycle and early pregnancy. Ovarian follicles and the CL undergo striking tissue remodeling involving multiple proteolytic events, during which serine proteases of plasminogen activation cascade have been shown to play crucial roles (Hagglund et al. 1996; Ny et al. 2002; Cao et al. 2004; Liu 2004). Our results showed that prostasin serine protease was expressed in oocytes, granulosa cells, theca cells, and/or the stroma of growing follicles during early antral and antral stages, with the highest level of expression in preovulatory dominant follicles, supporting a hypothesis that prostasin contributes to follicular growth and ovulation. Prostasin was also localized at high levels in the CL after ovulation, suggesting that it may be involved in matrix degradation and remodeling during CL formation. These speculated functions of prostasin would be similar to the roles of other serine proteases, such as uPA in early growing follicles (Ny et al. 2002; Liu 2004), tPA (Ny et al. 2002; Liu 2004) and kallikrein (Clements et al. 1997) in preovulatory follicles, and uPA (Liu 2004) and tPA (Liu et al. 1996) in CL formation. In addition, during early pregnancy, the high level of expression of prostasin mRNA and protein in the CL may suggest that prostasin has a functional role in CL formation and maintenance during early pregnancy.
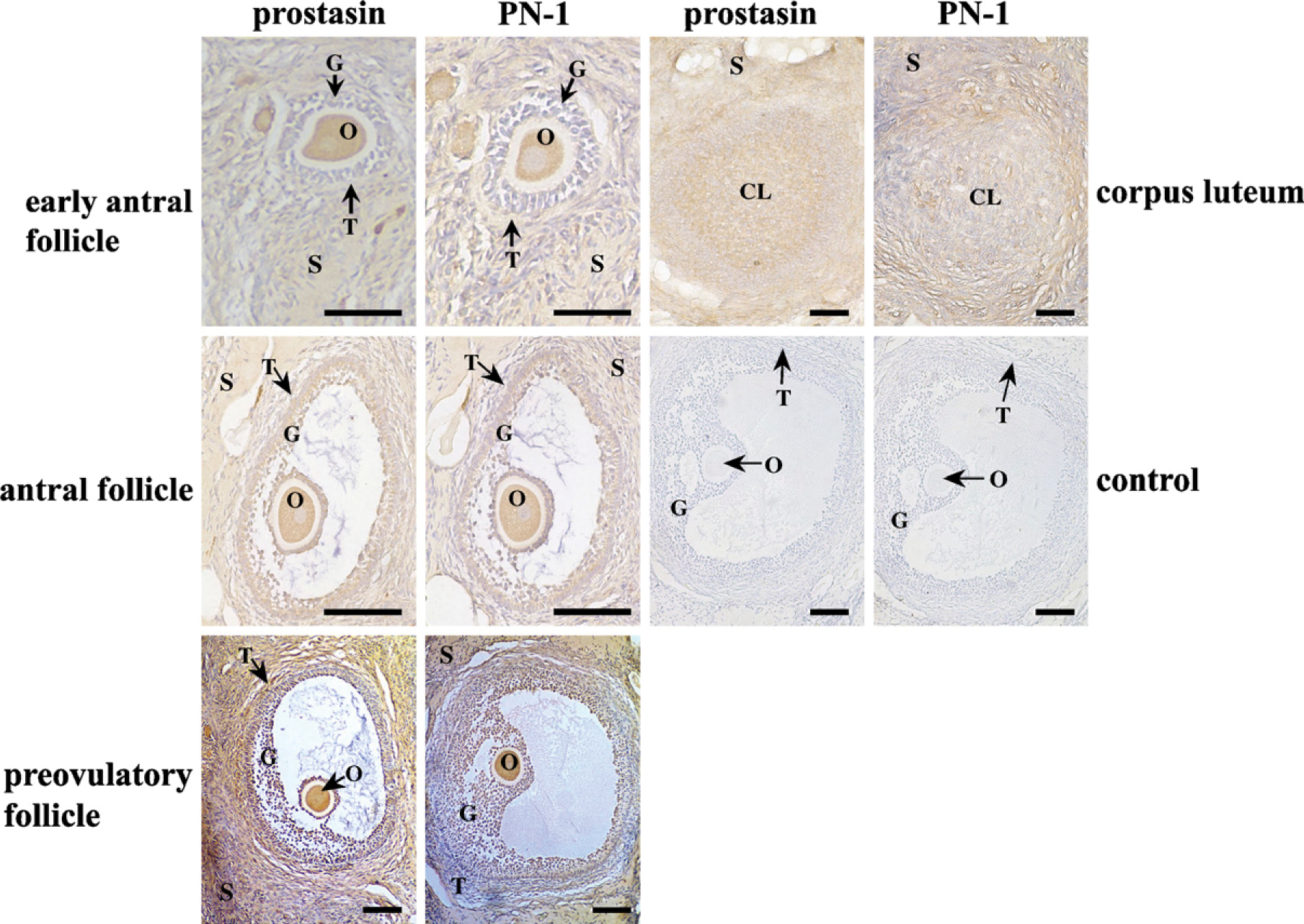
Immunohistochemical staining of prostasin and PN-1 proteins in macaque ovaries during the menstrual cycle. Control, control sections immunostained with normal rabbit IgG. (See Figure 1 legend for other abbreviations.) Bar = 200 μm
Densitometric quantification of prostasin and PN-1 protein in different histological compartments of the macaque ovary during the menstrual cycle
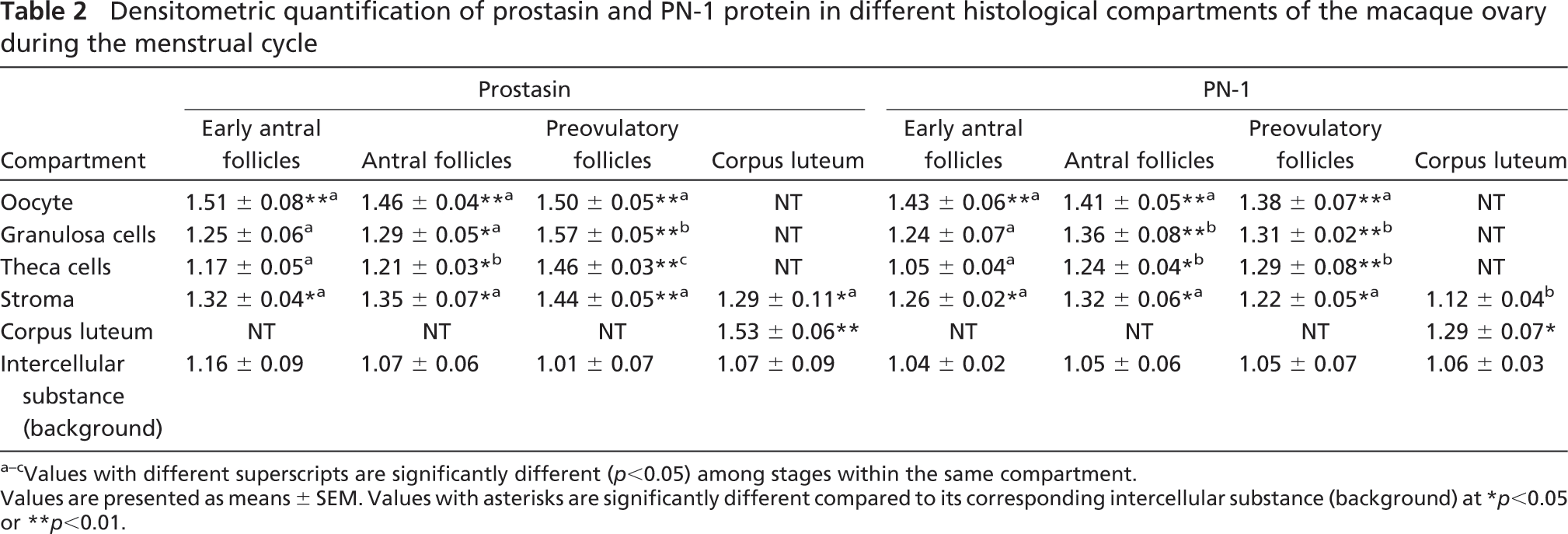
Values with different superscripts are significantly different (p<0.05) among stages within the same compartment.
Values are presented as means ± SEM. Values with asterisks are significantly different compared to its corresponding intercellular substance (background) at ∗ p<0.05 or ∗∗ p<0.01.
Many lines of evidence exist to show that PN-1 is involved in controlled proteolysis during ovarian tissue remodeling. PN-1 is expressed at the highest level in granulosa cells of growing dominant bovine follicles (Bedard et al. 2003) and is decreased after human chorionic gonadotropin injection, contributing to ovulation (Bedard et al. 2003). In rats and mice, strong PN-1 expression is also observed in granulosa cells of developing follicles from the secondary stage until ovulation (Hasan et al. 2002), and granulosa cells of rat large preovulatory follicles express an abundance of PN-1 (Hagglund et al. 1996; Hasan et al. 2002), which decreases in its level after hCG injection (Hasan et al. 2002) or in postovulatory follicles (Hagglund et al. 1996). In this study, we showed that, during the macaque menstrual cycle, PN-1 was expressed in oocytes, granulosa cells, theca cells, and the stroma of developing follicles, with the highest level in preovulatory dominant follicles. Collectively, these observations are consistent with the role of PN-1 in controlled proteolysis contributing to follicular development and ovulation. The increasing expression of PN-1 in the monkey CL, suggesting its role in CL formation, is in agreement with the localization of PN-1 protein in the bovine CL (Bedard et al. 2003). However, PN-1 was previously not detected in the rat CL (Kim et al. 2001; Hasan et al. 2002), indicating a species difference in the spatiotemporal pattern of PN-1 expression in CL.
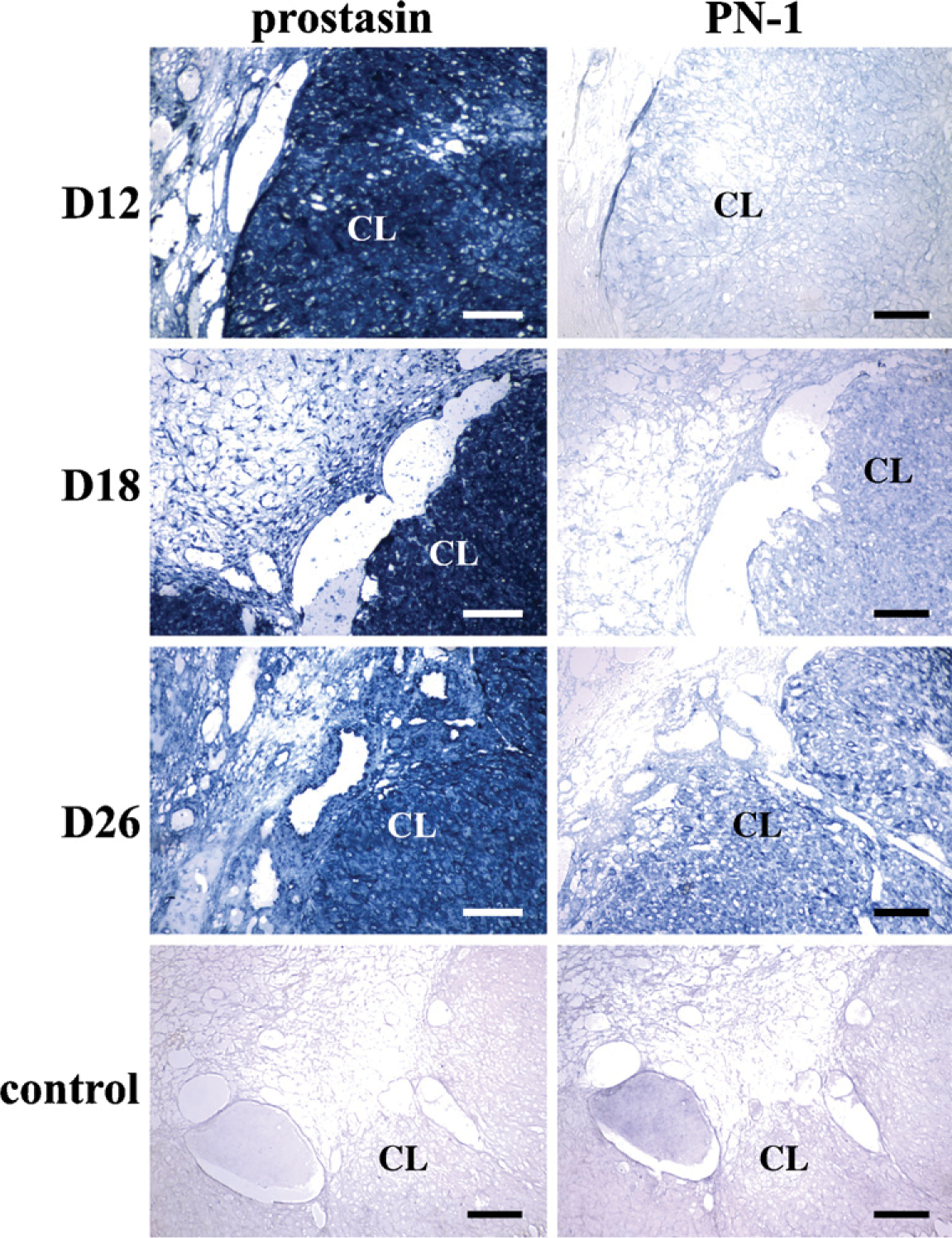
Localization of prostasin and PN-1 mRNA in macaque ovaries during early pregnancy by in situ hybridization. Various stages are shown: day 12 (D12), day 18 (D18), and day 26 (D26). Control, control sections hybridized with sense probe. (See Figure 1 legend for other abbreviations.) Bar = 200 μm.
Densitometric quantification of prostasin and PN-1 mRNA in different histological compartments of the macaque ovary on D12, D18, and D26 of pregnancy
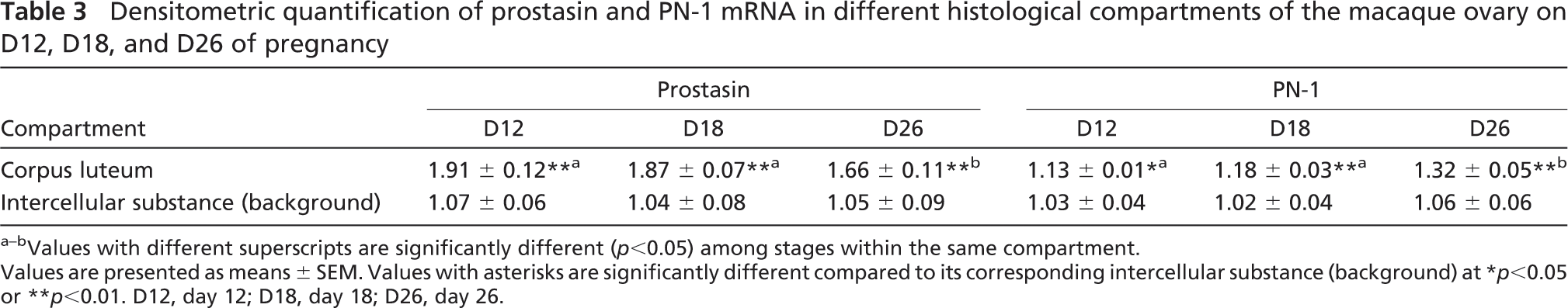
Values with different superscripts are significantly different (p<0.05) among stages within the same compartment.
Values are presented as means ± SEM. Values with asterisks are significantly different compared to its corresponding intercellular substance (background) at ∗p<0.05 or ∗∗p<0.01. D12, day 12; D18, day 18; D26, day 26.
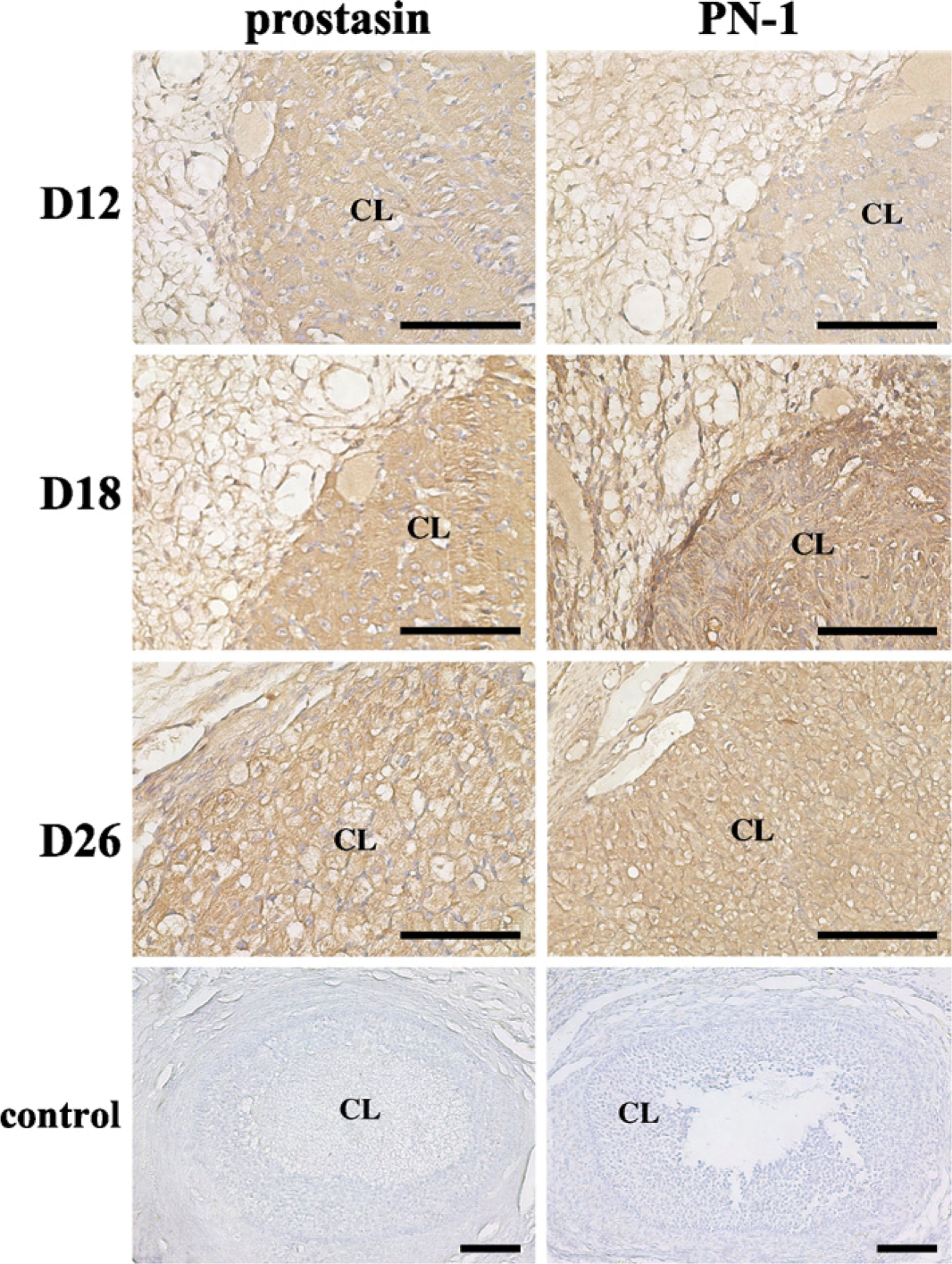
Immunohistochemical staining of prostasin and PN-1 proteins in macaque ovaries during early pregnancy. Various stages are shown: day 12 (D12), day 18 (D18), and day 26(D26). Control, control sections immunostained with normal rabbit IgG. (See Figure 1 legend for other abbreviations.) Bar = 200 μm.
Study on the distribution pattern of PN-1 in the macaque CL during early pregnancy led to the revelation of a progressive increase of PN-1 expression as pregnancy proceeded. A report on rats (Kim et al. 2001) provided evidence to show that PN-1 mRNA in the ovary was present at the highest level on day 6.5 postcoitus (implantation period) and was reduced on day 13.5 postcoitus (postimplantation). Taken together, these results suggest that, in early pregnancy, PN-1 is progressively involved in the control of matrix degradation during the process of CL formation and maintenance.
Densitometric quantification of prostasin and PN-1 protein in different histological compartments of the macaque ovary on D12, D18, and D26 of pregnancy
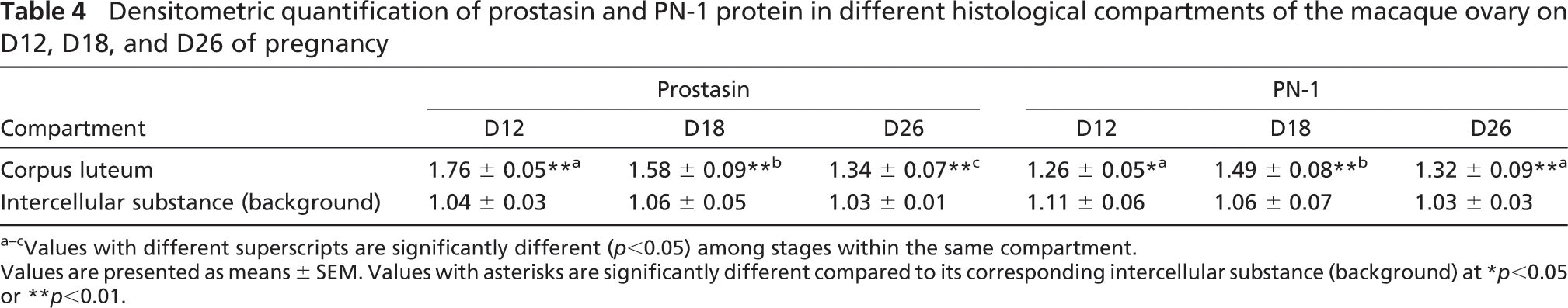
Values with different superscripts are significantly different (p<0.05) among stages within the same compartment.
Values are presented as means ± SEM. Values with asterisks are significantly different compared to its corresponding intercellular substance (background) at ∗p<0.05 or ∗∗p<0.01.
Because of the fact that proteolysis during follicular development, ovulation, and CL development has to be spatiotemporally restricted, the fine-tuned balance between the levels of protease and inhibitor is important to the control of tissue growth and remodeling. Earlier studies have showed that mouse granulosa cells express low levels of PN-1 and high levels of uPA, whereas cumulus cells express high levels of PN-1 and low levels of uPA, and these heterogeneous and complementary expression patterns for PN-1 and uPA provide the basis for regulation of proteolytic activity around the cumulus- oocyte complex (Hagglund et al. 1996). From recent in vitro studies in cultured small follicular cells, tPA activity was seen to increase with time of culture for antral but not basal cells, whereas PN-1 secretion increases in basal but not antral cells. In cultured granulosa cells from large follicles, increased tPA activity occurs as PN-1 secretion decreases. It was suggested from these data that there is a follicular stage-dependent regulation of tPA and PN-1 production in tissue remodeling of the granulosa cell layer (Cao et al. 2004). In this study, we showed that PN-1 was coordinately localized with prostasin in granulosa cells of developing follicles from the antral stage to the preovulatory stage and in the CL, suggesting that PN-1 may also be involved in the same controlled tissue remodeling during follicular development and CL formation by inhibiting the activity of another serine protease, prostasin.
In conclusion, we showed that prostasin serine protease is expressed in a pattern that is consistent with a role in matrix degradation and remodeling during follicular development, ovulation, and CL formation. PN-1 may be involved in controlled proteolysis in these processes by regulating the activity of prostasin, as suggested by its specific pattern of expression.
Footnotes
Acknowledgments
This study was supported by the Knowledge Innovation Program of Chinese Academy of Sciences (KSCX3-IOZ-07), the Department of Defense Prostate Cancer Research Program (to KXC, Grant DAMD17-02-1-0032), and the National Institutes of Health (to L-MC, Grant HD 40241).