
Abstract
This study was designed to learn more about the changes in expression of rat anterior pituitary (AP) leptin during the estrous cycle. QRT-PCR assays of cycling rat AP leptin mRNA showed 2-fold increases from metestrus to diestrus followed by an 86% decrease on the morning of proestrus. Percentages of leptin cells increased in proestrus and pregnancy to 55-60% of AP cells. Dual labeling for leptin proteins and growth hormone (GH) or gonadotropins showed that the rise in leptin protein-bearing cells from diestrus to proestrus was mainly in GH cells. Only 10-20% of leptin cells in male or cycling female rats coexpress gonadotropins. In contrast, 50-73% of leptin cells from pregnant or lactating females coexpress gonadotropins and only 19% coexpress GH, indicating plasticity in the distribution of leptin. Leptin cells expressed GnRH receptors, and estrogen and GnRH together increased the coexpression of leptin mRNA and gonadotropins. GnRH increased cellular leptin proteins three to four times and mRNA 9.8 times in proestrous rats and stimulated leptin secretion in cultures from diestrous, proestrous, and pregnant rats. These regulatory influences, and the high expression of AP leptin during proestrus and pregnancy, suggest a supportive role for leptin during key events involved with reproduction.
Keywords
L
However, leptin has also recently been recognized as a multifunctional paracrine regulator in a number of organs including the anterior pituitary (AP) (Jin et al. 1999, 2000; Morash et al. 1999; Vidal et al. 2000; Popovic et al. 2001; Lloyd et al. 2001; Sone and Osamura 2001; Sone et al. 2001; McDuffie et al. 2004). In humans, leptin proteins are found in subsets of corticotropes, somatotropes, gonadotropes, or thyrotropes (Jin et al. 1999; Vidal et al. 2000). Dual labeling at the electron microscopic level shows that leptin proteins are stored in secretory granules (Vidal et al. 2000). Recent studies in our laboratory detected the expression of leptin mRNA and proteins by somatotropes (McDuffie et al. 2004). However, the percentages of leptin protein-bearing cells varied with the reproductive state. Therefore, we hypothesized that gonadotropes might also be a source of leptin. We also postulated that regulators of gonadotropes might be involved in the production of anterior pituitary leptin.
One important candidate regulator proposed for testing was gonadotropin-releasing hormone (GnRH). This neuropeptide is produced by neurons scattered in the preoptic and anterior regions of the hypothalamus, stretching back to the arcuate nucleus (Conn et al. 1987; Clayton 1989; Conn 1994). GnRH is secreted in pulses. GnRH pulse amplitude and frequency change as the rats approach midcycle. Slower pulses of GnRH, seen earlier in the cycle, favor follicle-stimulating hormone (FSH) secretion. Higher amplitude pulses during proestrus favor luteinizing hormone (LH) secretion over FSH secretion and eventually lead to the LH surge. (Belchetz et al. 1978; Savoy-Moore et al. 1980; Wildt et al. 1981; Levine and Ramirez 1982; Loumaye and Catt 1982; Crowley et al. 1985; Haisenleder et al. 1991; Kaiser et al. 1997; Burger et al. 2002; Bédécarrats and Kaiser 2003). To enhance their response to GnRH, estrogens stimulate the production of GnRH receptors, which reach a peak late in diestrus or early in proestrus (Lloyd and Childs 1988; Childs et al. 1994b). These known cyclic changes in GnRH receptors were used in the design of tests of GnRH effects on pituitary leptin.
In the present group of studies, we continued our analysis of changes in the expression of leptin with different reproductive states, adding tests of leptin expression by pituitaries from rats during the afternoon of proestrus as well as tests of pituitaries from pregnant or lactating rats. We tested the efficacy of GnRH in the regulation of AP leptin expression, comparing exposures of 1 hr and 3 hr in cells from rats in proestrus. The findings presented in this report show that the expression of leptin proteins and mRNA in normal male and cycling female rats predominates in somatotropes. However, coexpression of leptin mRNA and proteins can also be found in subpopulations of gonadotropes, which predominate in pregnant or lactating females. The study will also present in vitro data demonstrating that leptin-bearing cells express GnRH receptors, and that GnRH stimulates the expression of leptin mRNA, proteins, and secretion.
Materials and Methods
Animals
Male and female Sprague Dawley rats weighing 200-250 g were obtained from Harlan Sprague Dawley (Indianapolis, IN) and were acclimated and housed three to four per cage with a 12 hr on, 12 hr off light/dark cycle for 7-10 days before use. Vaginal smears were used to determine the stage of the cycle, as previously described (McDuffie et al. 2004). The animal use protocol was approved annually by the Animal Care and Use Committee, with regular updates as specified in the guidelines.
To obtain pregnant females, proestrous females were placed individually in the same cage with a male for 1 night. The smear was checked for sperm the next morning. We then separated the female, if pregnant, and sacrificed her on the fourth day at 10:00
Pituitaries from all animals were taken either at 10:00
Stimulation of Dispersed Pituitary Cells
For secretion and cytochemical studies, freshly dispersed pituitary cells were used for several reasons. First, the use of whole cells for the detection of coexpression of two products prevented artifacts often seen in sections, which make it impossible to differentiate between a neighboring process filled with label and a patch of label at the periphery of a cell. Sections also may miss detection of dual label in areas in the cell that are above or below the plane of the section. Living, dispersed cells were also needed to detect GnRH receptors by biotinylated analogs in sectioned material (Childs et al. 1983a,b).
Tests of expression of leptin and other pituitary hormones show that there are no changes in percentages of labeled cells when the culture is extended for as long as 48 hr. Percentages of LH and GH cells are similar to those freshly dispersed cultures as they are in sections; however, labeling is more intense in the whole cells and it is easier to detect labeled small cells. The enzymatic dispersion process and the subsequent 1-2 hr of culture were not deleterious to the detection of product. In cases where we use a longer incubation in secretagogue, estrogen, or vehicle, percentages of leptin-bearing cells remain comparable to those in freshly dispersed cultures.
After 1-2 hr of plating, freshly dispersed pituitary cells from proestrous female or male rats were treated for 1 hr or 3 hr with vehicle or 10 pM-1000 pM GnRH. The vehicle was defined as serum-free media (McDuffie et al. 2004). Cells were fixed and then prepared for in situ hybridization or immunocytochemistry (McDuffie et al. 2004). Media were collected for leptin enzyme immunosorbent assay (EIA).
Immunocytochemistry
Immunolabeling for leptin was done as previously described (McDuffie et al. 2004) with a working dilution of anti-leptin (1:37,500; Sigma, St Louis, MO). Dual labeling for leptin and GH or LHβ was done by first detecting leptin with the streptavidin-biotin-peroxidase protocol. Then GH, FSHβ, or LHβ was detected by the ImmPRESS protocol according to kit instructions (Vector Laboratories; Burlingame, CA), as described previously (McDuffie et al. 2004; Childs et al. 2005). Dilution of anti-rat GH was 1:200,400 and dilution of anti-rat LHβ or anti-human FSHβ was 1:30,000-1:40,000. Controls are illustrated in the previous reports for leptin antigens (McDuffie et al. 2004). Controls for the new protocols that detected GH antigens or mRNA are reported in Childs et al. (2005) or Iruthayanathan et al. (2005). Those for the anti-LHβ or anti-FSHβ antisera were described in Childs et al. (1994a).
Affinity Cytochemistry for GnRH Receptors
To detect GnRH receptors on leptin-bearing cells, pituitary cells from diestrous rats were stimulated for 10 min with 1 nM biotinylated GnRH, which was detected as previously described (Childs et al. 1983a,b,1994b; Lloyd and Childs 1988). This was followed by immunolabeling for leptin antigens with streptavidin-peroxidase and amber-orange DAB as previously described (Childs et al. 1983b; McDuffie et al. 2004). Controls included competition with unlabeled GnRH for receptor sites, which eliminated black labeling for GnRH and absorption of the anti-leptin with leptin, which eliminated amber labeling for leptin.
Once GnRH binding was detected on cells with leptin proteins, further tests of diestrous rats were conducted in which cells were treated overnight with vehicle or 100 pM estradiol (Childs et al. 2005) and then exposed to vehicle or 1 nM GnRH for 1 hr or biotinylated GnRH for 10 min. These conditions had previously been shown to increase the numbers of target cells for GnRH (Lloyd and Childs 1988). Cells were then fixed and prepared for detection of biotinylated GnRH or leptin mRNA and LHβ or FSHβ.
In Situ Hybridization
In situ hybridization was carried out as described previously (McDuffie et al. 2004), with modifications (Childs et al. 2005). Leptin mRNA was hybridized with the 48-bp biotinylated oligonucleotide probe complementary to nucleotides 342-389 located within the coding sequence for rat leptin (accession number NM_013076). This probe and the control sequences were produced by
Analysis of Labeling and Statistics
Cytochemical labeling was analyzed by either cell counts or by Bioquant Nova Image Analysis equipment (Bioquant Image Analysis Corp.; Nashville, TN) with an algorithm that integrated label density and area and changes in both numbers of labeled cells as well as the amount of label per cell. This protocol is described in a recent report (Iruthayanathan et al. 2005). The approach for the statistical analysis including the power analysis is also described in recent reports (McDuffie et al. 2004; Childs et al. 2005; Iruthayanathan et al. 2005).
EIA
The leptin mouse/rat EIA kit by American Laboratory Products Company (ALPCO Diagnostics; Salem, NH) was used to detect serum leptin proteins. EIA was performed following kit instructions. Inter- and intraassay variation coefficients were <4.7% and <4.4%, respectively. The kit included the production of sample dilutions of leptin for assay, which was found to be linear over the standard range. Limit of sensitivity of the kit was 10-20 pg/ml.
Controls for the detection of leptin mRNA are shown. (
RNA Extraction, cDNA Synthesis, and QRT-PCR
Whole pituitaries from cycling female rats were used for these studies. We analyzed leptin mRNA expression in three to five rats/stage or time in the cycle. After sacrifice by decapitation, the whole pituitary was placed in RLT buffer (Qiagen; Valencia, CA) containing β-mercaptoethanol (as per manufacturer's protocol). The ratio was 100:1-RLT:β-mercaptoethanol. RNA was then extracted as described in previous studies, including the DNase steps that removed genomic DNA (Iruthayanathan et al. 2005).
iScript cDNA synthesis (Bio-Rad Laboratories; Hercules, CA) was used to reverse transcribe total RNA in an MJ PTC 150 Minicycler (MJ Research; Cambridge, MA) in a 20-μml reaction mixture containing 4 μl 5 × iScript, 1 μl reverse transcriptase, and 15 μl RNA per manufacturer's protocol (25C at 5 min, 50C at 30 min, and 85C at 5 min). Tests of serial dilutions for the cDNA samples showed reproducible assays of leptin mRNA with dilutions spanning 1:10-1:100. Aliquots were frozen at −80C and diluted 1:10 and 1:100 for the QRT-PCR assay for leptin mRNA.
Standards for each gene for QRT-PCR were prepared according to the method of Zhou et al. (2003) and amplified as described in our previous study (Iruthayanathan et al. 2005). A nested primer strategy was used to amplify leptin cDNA to provide a high enough concentration for the standards. This involved the use of two sets of primers, which increased the yield of amplicon. The pituitary leptin gene was cloned and used to produce cDNA for the template, which was then amplified by PCR in two rounds. The first round of PCR was done with forward and reverse primers F1 and R1, which amplified a 300-bp region from nucleotide 105 to 404 in NM_013076.1. The second round of PCR used primers F2 and R2, which amplified regions 179-249 nested in the first amplicon. The product was sent for sequencing and found to be identical to rat leptin. To make the standards, each cDNA fragment was diluted to 4.15 amol/μl and frozen. For the QRT-PCR assays, eight 10-fold serial dilutions were made for use as standards.
QRT-PCR assays were run in a Roche Light Cycler 1.2 (Roche Applied Sciences; Indianapolis, IN). QRT-PCR was carried out as in our previous study with the FAST-START DNA Master SYBR Green I enzyme mix (Roche Applied Sciences). The housekeeping gene used to normalize the readings was hypoxanthine guanine phosphoribosyl-transferase (HPRT) as described in our previous publication (Iruthayanathan et al. 2005). HPRT did not change with the stage of the reproductive cycle.
During the course of running the leptin assays, we experienced some difficulties with the SYBR green detection system, as it often resulted in a product that included primer dimers, rendering the final product levels uninterpretable. Therefore, we switched to the Roche Applied Sciences “Universal Probe System”, which was run according to the kit instructions. The Universal probe designed for the detection of leptin was #13 (Cat. #04685121001). Universal Probe #13 reacts with nucleotides 220-227, aggcagag in the leptin cDNA template. The forward primer designed by Roche for the amplification of leptin was ccaggatcaatgacatttcaca (nucleotides 179-200) and the reverse primer was aatgaagtccaaaccggtga (nucleotides 230-249) in NM_013076.1. The amplicon is a 71-bp sequence that includes one 1564-bp intron-spanning region at bp 203-204. We used the same standards that were produced for the SYBR Green protocol.
The Lightcycler Taqman Master Kit (Roche Applied Sciences) was prepared as described in the instructions (Cat. #04535286001), and then a master mix was prepared that included the following components (multiplied by a factor that varied with the numbers of tubes). For each tube there was 10.4 μl of nuclease-free water, 0.2 μl of Universal probe #13 (10 μM stock), 0.2 μl each of forward and reverse primers (20 μM used to make a final concentration of 200 nM), and 4 μl of the prepared TaqMan master. The master mix was added (15 μl) to each of the glass tubes, and then 5 μl of standard (101-105), 1:10 diluted sample cDNAs or Tris was added to individual tubes. All samples were run in duplicate. The Tris or water control served as a negative control. After the tubes were set up, they were centrifuged for 10 sec and then placed in the Light Cycler that was programmed as follows: preincubation—95C, 10 min, 45 cycles: denaturation—95C, 10 sec; annealing—60C, 30 sec; and extension—72C, 1 sec. After 45-55 cycles they were cooled at 45C for 1 min. This protocol allowed us to successfully detect product in all samples. It avoids the formation of primer dimers, thus having the advantage of being more specific. With these primers, only leptin amplicons are detected by Universal probe #13.
Results
Changes in Leptin Expression With the Reproductive State
We previously reported changes in leptin proteins (McDuffie et al. 2004) with the stage of the cycle. The continuing analysis in this study added more data detecting both mRNA and proteins, including that from rats taken at 2:00
There is a gradual rise in percentages of AP cells with leptin proteins from a low of 21.0 ± 4.0% on the morning of estrus to a peak of 55.0 ± 3.0% of AP cells on the afternoon of proestrus. Figures 2C-2E illustrate fields labeled for leptin proteins, comparing labeling on the morning and evening of proestrus and showing reduced labeling on the morning of estrus. Peak expression of leptin during proestrus is higher than that in all other groups (p<0.001) including male rats, which had 39.6 ± 1.0% of AP cells with leptin proteins.
Leptin mRNA is expressed in 32-37% of pituitary cells in male or in estrous, metestrous, or diestrous female rats (Figure 2). Thus, early in the cycle (estrus and metestrus), there are more cells with leptin mRNA than leptin proteins. There are no significant differences among these groups. However, there is a significant decline (p<0.002) in the percentages of cells with leptin mRNA to 20.0 ± 3.0% on the morning of proestrus. This reduction on the morning of proestrus was confirmed by the QRT-PCR assays (Figure 2B), which showed a significant 86% decline from diestrous afternoon to proestrous morning (p<0.007). QRT-PCR assays show that leptin mRNA is maintained at relatively low levels through estrus. Then, levels rise during metestrus and diestrus. The 2-fold rise in diestrus morning and afternoon is to values higher than all other groups (p<0.015). There are no differences between the two diestrous values.
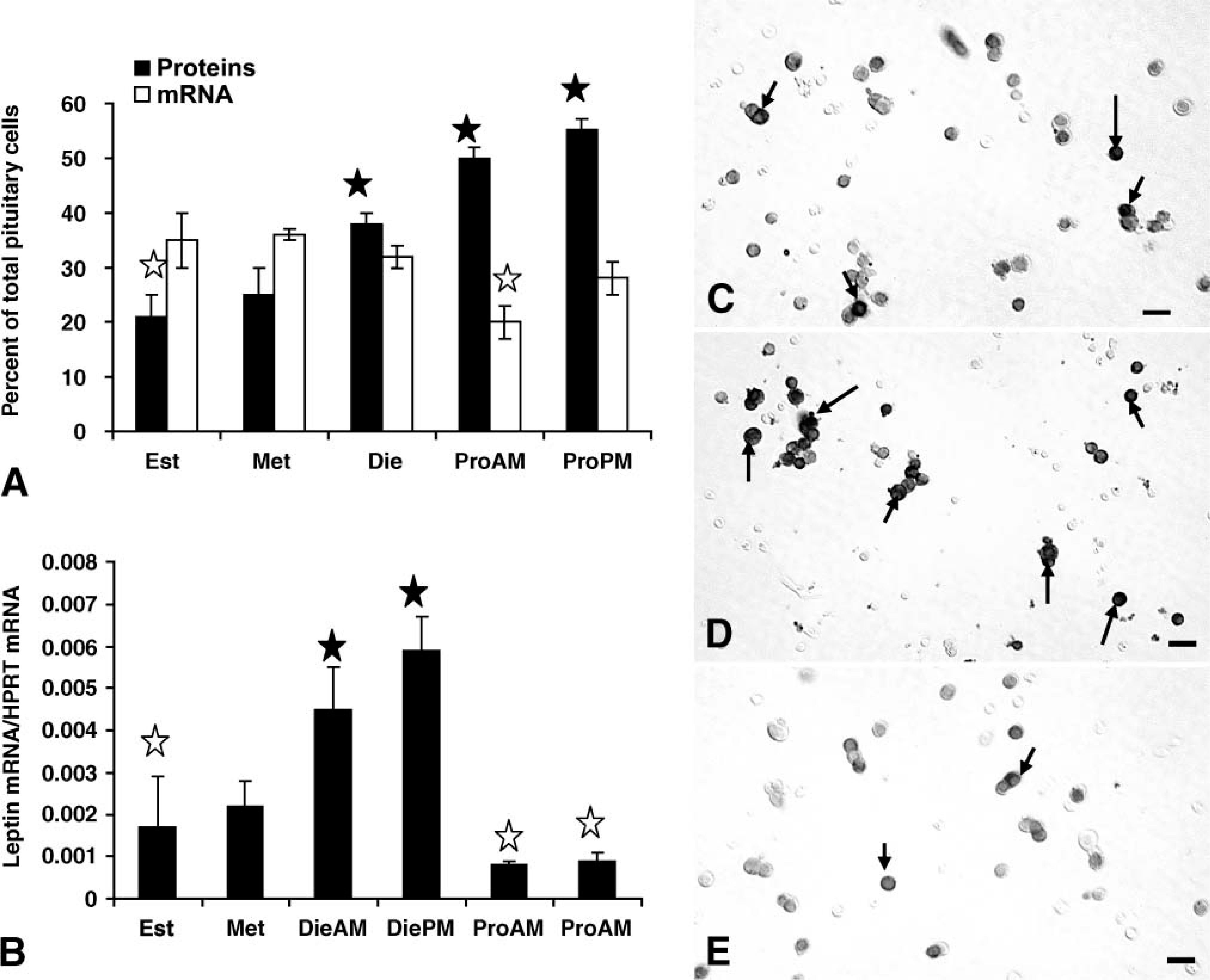
Expression of pituitary leptin proteins and mRNAin different stages of the cycle. (
Increase in mRNA from metestrus to diestrus detected by the QRT-PCR assays was not detected by a change in percentages of leptin-bearing cells. However, densitometric analysis showed that the total area of label for leptin mRNA increased from 844.0 ± 114.0 μm2 to 1246.0 ± 15.0 μm2 (p<0.01) during this period.
To learn if other reproductive states were associated with changes in pituitary leptin, cell populations from pregnant or lactating rats were studied. Figure 3 shows that populations from pregnant rats contained relatively the highest percentages of AP cells with leptin proteins (60.0 ± 2.0%) or mRNA (44.0 ± 2.0%), when compared with all other groups. Percentages of AP cells with leptin proteins are higher than those from males and all cycling groups except females taken on the afternoon of proestrus. The values for mRNA-bearing cells are higher than those from all other groups (p<0.001).
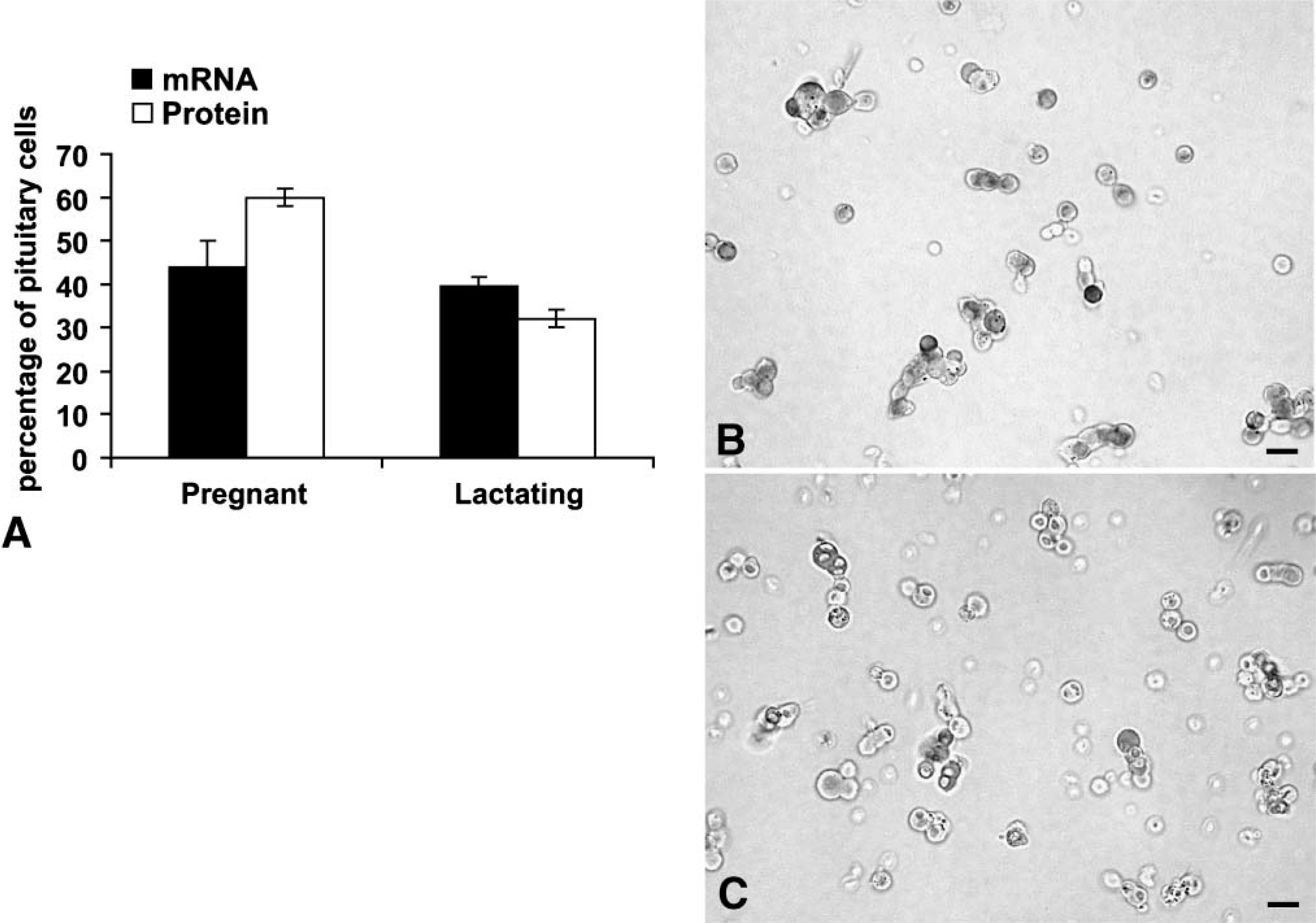
(
AP populations from females taken on the third day of lactation had mid-range levels of leptin proteins when compared with other physiological states. There were 32.0 ± 3.0% cells with leptin proteins and 40.0 ± 2.0% cells with leptin mRNA (Figure 3). Percentages of AP cells with leptin proteins in lactating rats are higher than those from estrous rats (p<0.02) and lower than those from proestrous (p<0.047) or pregnant rats (p<0.001). Percentages of AP cells with leptin mRNA are higher than all groups except pregnant rats. Photographs of fields from these rats are also illustrated in Figure 3.
Cell Types That Coexpress Leptin Proteins
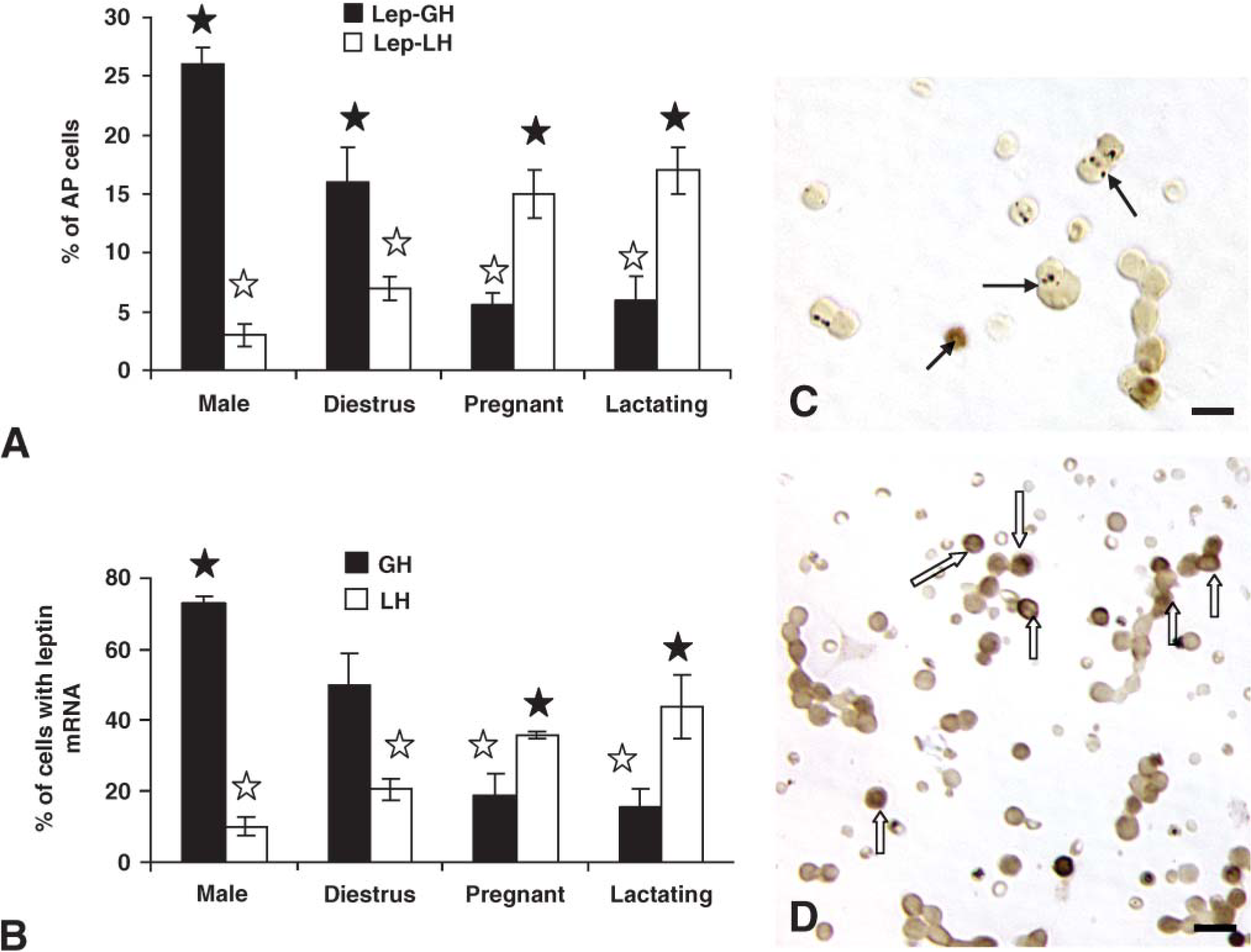
Our previous study reported that most leptin-bearing cells were somatotropes. However, the timing and direction of changes in expression during the cycle (Figure 2) suggested that gonadotropes may be involved. Dual immunolabeling was therefore done on some of these experimental groups to test this hypothesis. The main question was focused on whether or not somatotropes or gonadotropes, or both, contributed to the rise in the percentage of leptin-bearing cells during proestrus and pregnancy.
When coexpression of leptin proteins and GH proteins was tested, there was a significant increase (p<0.001) in the percentages of dual-labeled AP cells from 25.0 ± 3.0% on the morning of diestrus to 37.0 ± 4.0% of the population on the morning of proestrus (Figure 4). This increment (12 percentage points) matches that seen when the total percentages of AP cells with leptin were calculated (Figure 2). The overall percentage of AP cells with GH proteins did not change significantly. It was 36.0 ± 6.0% on the morning of diestrus and 41.0 ± 4.0% on the morning of proestrus.
In contrast, the percentage of AP cells that coexpressed leptin and gonadotropins did not change from diestrus to proestrus. Diestrous populations had 6.5 ± 1.0% or 7.0 ± 1.0% cells with leptin and LH or FSH, respectively, and proestrous populations had 8.0 ± 2.0% AP cells with leptin and LH or FSH (Figure 4). Previous studies have shown that bihormonal (cells with both LH and FSH) gonadotropes predominate in the population of diestrous and proestrous rats, representing >70% of the gonadotrope population (Childs et al. 1987,1994a), which suggests that leptin is being expressed by cells that are mostly bihormonal.
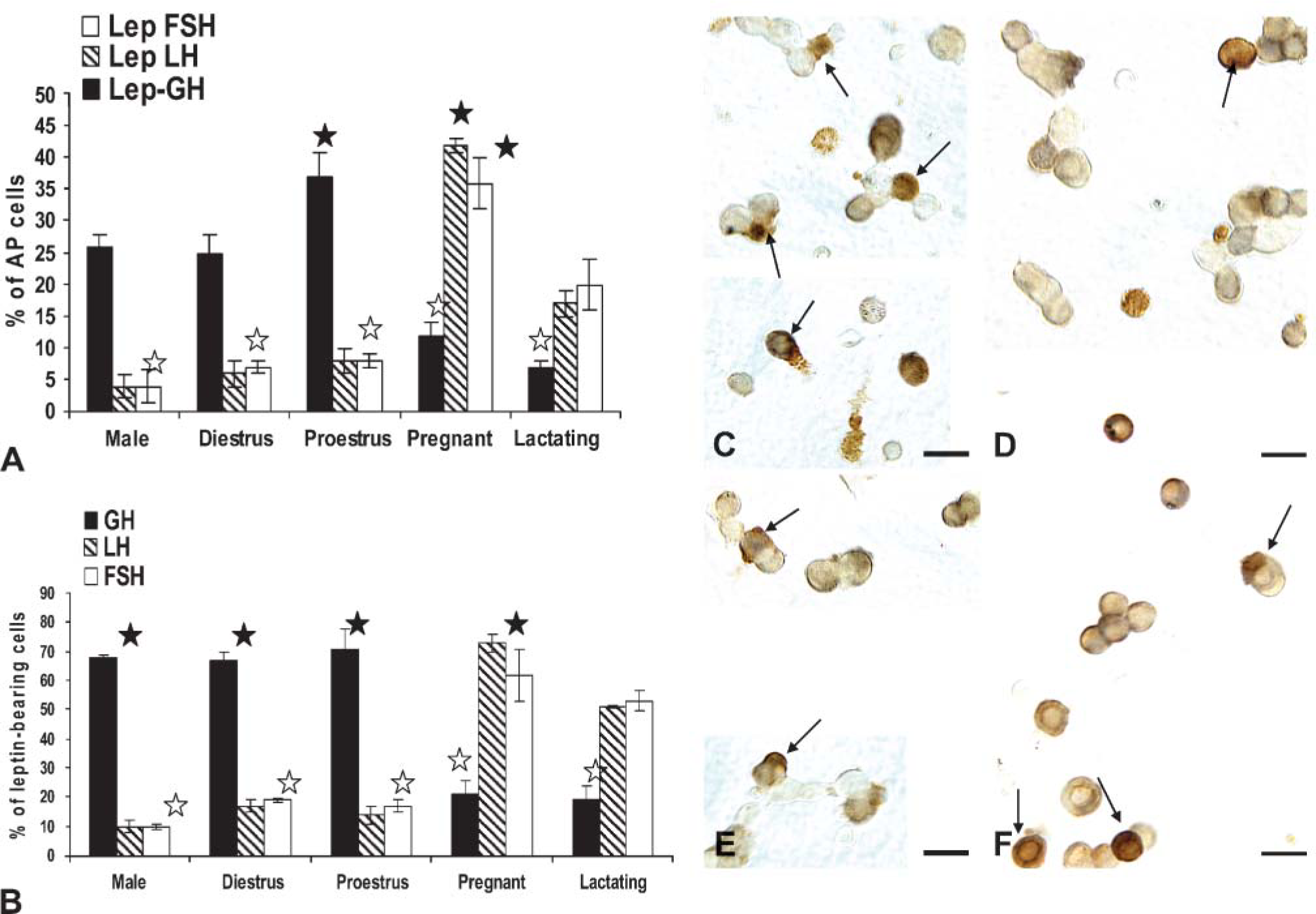
Top graph (
Cultures from male rats exhibited a profile similar to that of diestrous rats with 26.0 ± 2.0% of AP cells coexpressing leptin and GH. Only 4.0% of AP cells coexpressed leptin and LH or FSH in the male, values which were also not different from diestrous rats. Those for AP cells coexpressing leptin and LH were significantly lower than percentages seen on the morning of proestrus (p=0.045).
Significant plasticity in expression was seen in populations from pregnant or lactating rats (Figure 4A) as leptin-bearing cells shifted from being predominantly somatotropes to predominantly gonadotropes. There were no significant changes in the percentages of cells with GH proteins in pregnant or lactating rats. There was a significant reduction (p<0.001) in the percentages of AP cells that coexpressed GH and leptin proteins to 12.0 ± 2.0% in the populations from pregnant females and 6.6 ± 1.0% in those from lactating females. In pregnant rat cells, there was a significant rise in percentage of AP cells that coexpressed LH or FSH and leptin to 42.0 ± 1.0% or 36.0 ± 4.0%, respectively. Cells from lactating females also had significantly more AP cells with LH and leptin (18.7 ± 0.4%) or FSH and leptin (17.0 ± 1.0%) than all other groups except pregnant rats.
The analysis also focused on the proportion of leptin-bearing cells that expressed each of the pituitary hormones tested. These values were also used to predict if other cells contributed to leptin-bearing cells and to validate the data in Figure 4A. Validation was done by manually multiplying the percentages of leptin cells that contained each of the hormones by the overall percentages of AP cells that contained leptin, as reported in Figure 2 and Figure 3. These calculated percentages were within 0.5-1.0 percentage points of those derived from the cell counts in Figure 4A.
GH stores are found in 67-71% of leptin protein-bearing cells in males or in diestrous or proestrous females, values that are not significantly different from one another (Figure 4B). In contrast, only 19-20% of leptin cells coexpress GH proteins in pregnant or lactating animals, which is significantly lower than values from the other groups (p<0.001).
LH is found in 14-17% and FSH is found in 17-19% of leptin cells in diestrous and proestrous rat populations, values that are not different from one another. As stated above, it is likely that leptin may be expressed in gonadotropes, at least half of which are bihormonal (store both LH and FSH). In populations from pregnant rats, most leptin-bearing cells express LH (73.0 ± 2.0% of leptin cells) and/or FSH (62.0 ± 6.0% of leptin cells). These data show clear overlap in the percentages of leptin-bearing cells with gonadotropins, supporting the hypothesis that leptin is expressed in part by bihormonal gonadotropes.
In male rats, only 10% of leptin cells coexpress LHβ or FSHβ proteins. LH values are significantly lower (p< 0.006) than all female groups, except those from proestrus morning. The percentage of leptin cells with FSH in the male is lower than all female groups (Student's t-test).
Figure 4 also depicts the dual labeling for leptin proteins and GH (Figures 4C and 4D) or LHβ proteins (Figures 4E and 4F) in proestrous (Figures 4C and 4E) or pregnant (Figures 4D and 4F) rats. The fields dual labeled for leptin and GH show the contrast between the numerous dual-labeled cells in the proestrous female rat (Figure 4C) with only one in the field from the pregnant rat (Figure 4D). Similarly, few LH cells coexpress leptin in the field from the proestrous rat (Figure 4E), and there are numerous dual-labeled LH-leptin cells in the field from the pregnant rat (Figure 4F)
Cell Types That Coexpress Leptin mRNA
Dual labeling for leptin mRNA and LH or GH proteins was also done on some of these animals, focusing on diestrus because it was a peak time of expression of mRNA. Figure 2 showed that during most stages of the cycle, except diestrus, leptin mRNA was found in 32-36% of pituitary cells. In diestrous cell populations, 19.0 ± 2.0% of AP cells coexpressed leptin mRNA and GH, which is 51% of leptin mRNA-bearing cells (Figure 5). Among LHβ gonadotropes, the percentages were similar to those seen when leptin protein-bearing cells were counted (7.25 ± 1.0% of AP with leptin mRNA and LHβ). Similar results were seen for FSHβ labeling (data not shown).
The loss in cells coexpressing GH and leptin mRNA was again evident in pregnant and lactating females; values are significantly lower than values in diestrous animals (p<0.001) (Figure 5A). In contrast, more AP cells coexpressed leptin mRNA and LHβ proteins (1617%), values which were significantly higher than those from diestrous rats (p<0.001).
In cells from male rats, 26.0 ± 1.0% of AP cells express leptin mRNA and GH proteins, which is comparable to the levels seen in the dual-immunolabeled fields and greater than values in the diestrous female or those from pregnant or lactating animals (p<0.001) (Figure 5A). When LH gonadotropes were analyzed, the percentages of AP cells with leptin mRNA and LHβ in the male were comparable to those seen after dual immunolabeling.
Percentages of leptin mRNA-bearing cells that contain GH are higher in the male than all of the female groups (73.0 ± 2.0%; p<0.001) (Figure 5B). Percentages of leptin mRNA-bearing cells that contain LHβ are only 10.0 ± 2.0% in cells from male rats and 20.0 ± 2.0% in those from diestrous females, which is significantly higher than values in the male (p<0.01; Student's t-test).
As in the case of dual immunolabeling, pregnant and lactating rats had more leptin-bearing cells with LHβ proteins (36.0 ± 1.0 or 44.0 ± 9.0% of leptin cells in pregnant or lactating groups, respectively). These values were not different from one another but were significantly higher than those from all other groups. Figures 5C and 5D also illustrate the dual labeling for leptin mRNA and GH or LHβ in pregnant rats, showing the low expression of leptin mRNA in GH cells (Figure 5C) and the high expression in cells with LHβ antigens (Figure 5D).
GnRH and Estrogen Effects on Leptin Expression
The timing of the rise in leptin protein expression during the cycle from diestrus to proestrus coincided with the rise in estrogen, which stimulates the production of GnRH receptors by gonadotropes (Lloyd and Childs 1988) and somatotropes (Childs et al. 1994a). We hypothesized that leptin expression might be regulated in vivo by GnRH pulses. To determine if GnRH could bind directly to leptin-bearing cells, we exposed freshly dispersed pituitary cells from three groups of diestrous rats (three rats/group) to 1 nM biotinylated GnRH for 10 min. After biotinylated GnRH was detected by avidin-biotin complexes (McDuffie et al. 2004), dual labeling was used to identify leptin in these cells. Figures 6A and 6B illustrate dual labeling for bio-GnRH and leptin proteins. The biotinylated neuropeptide is seen as dark patches on or near the surface of the cell, and leptin labeling is seen inside the cells. In diestrous rats, cells with GnRH receptors and leptin proteins are 11.5 ± 2.0%. This represented 30.0 ± 3.0% of leptin-bearing cells and 73.0 ± 3.0% of GnRH target cells.
Top graph (
Estrogen is a well-established modulator of GnRH receptors, and our previous studies have shown that 100 pM increases the percentage of GnRH target cells when given overnight to diestrous rats (Lloyd and Childs 1988). These studies used this experimental approach on an additional three groups of diestrous rats to learn if estrogen increased the number of GnRH receptors on leptin-bearing cells. Figure 6C shows that whereas estrogen does increase the overall percentage of GnRH target cells as in our previous studies (Lloyd and Childs 1988), it does not significantly increase the number of leptin-bearing cells that bind GnRH, which remain at 13.25 ± 2.0% of AP cells.
The next study was designed to learn if estrogen and GnRH could increase leptin expression by gonadotropes. Three groups of cells pooled from three diestrous rats/group were treated with and without 100 pM estradiol overnight and then given vehicle or 1 nM GnRH for 1 hr the next morning. They were then fixed and labeled for leptin mRNA followed by immunolabeling for LHβ or FSHβ. Neither estrogen nor GnRH alone stimulated more gonadotropes to express leptin mRNA (data not shown). However, Figure 6D shows that when added, estrogen and GnRH stimulated a significant increase in AP cells that coexpress LHβ and leptin mRNA from 7.0 ± 2.0% to 11.0 ± 3.0% (p< 0.03) of AP cells. Similarly, estrogen and GnRH together stimulated an even greater increase in the percentages of cells with leptin mRNA and FSHβ from 8.0 ± 3.0% to 15.0 ± 6.0% (p<0.02).
To test if the overnight incubation in estrogen may have caused losses in expression of leptin mRNA, these data were compared with those from freshly dispersed cultures (7.0 ± 1.0% of AP cells from diestrous rats express leptin mRNA and LHβ, as shown in Figure 5A). Similarly, 7.25 ± 2.8% of AP cells coexpress leptin mRNA and FSHβ in these same cultures. Both sets of findings point to no losses in leptin mRNA during the overnight incubation in estrogen.
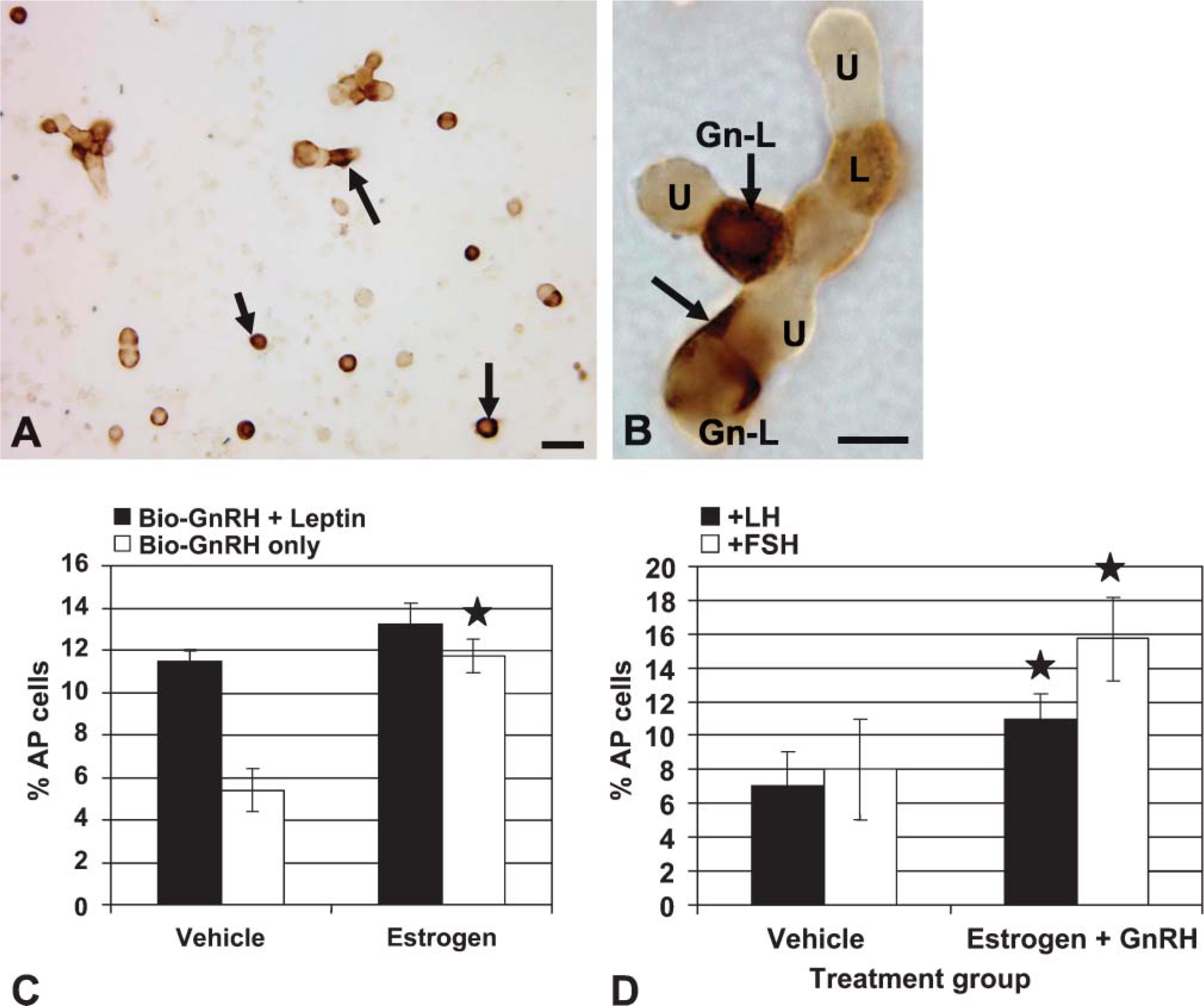
Top photograph depicts dual labeling for biotinylated analog of GnRH (black) followed by immunolabeling for leptin proteins (orange). Arrows in
GnRH Stimulation of Cellular and Secreted Leptin
Because GnRH receptors are at a peak late in diestrus, extending to the morning of proestrus (Lloyd and Childs 1988; Childs et al. 1994b), cells from proestrous morning female rats were studied to learn more about the specific effects of GnRH on leptin mRNA and protein expression.
Figure 7A illustrates the study of freshly dispersed cells from rats taken on the morning of proestrus. There is a significant increase in the average integrated optical density (IOD) of labeling after 1 hr in 100 pM GnRH (p<0.001), which plateaus at 500 pM. However, there is a significant decrease in IOD of labeling for leptin proteins after 1 nM GnRH (p<0.03) when compared with that following 500 pM. The value for 1 nM GnRH is still higher than the IOD for the vehicle-treated group (p=0.009). Similar studies were done of cells treated for 3 hr with GnRH, and there were no differences in expression of leptin proteins or mRNA.
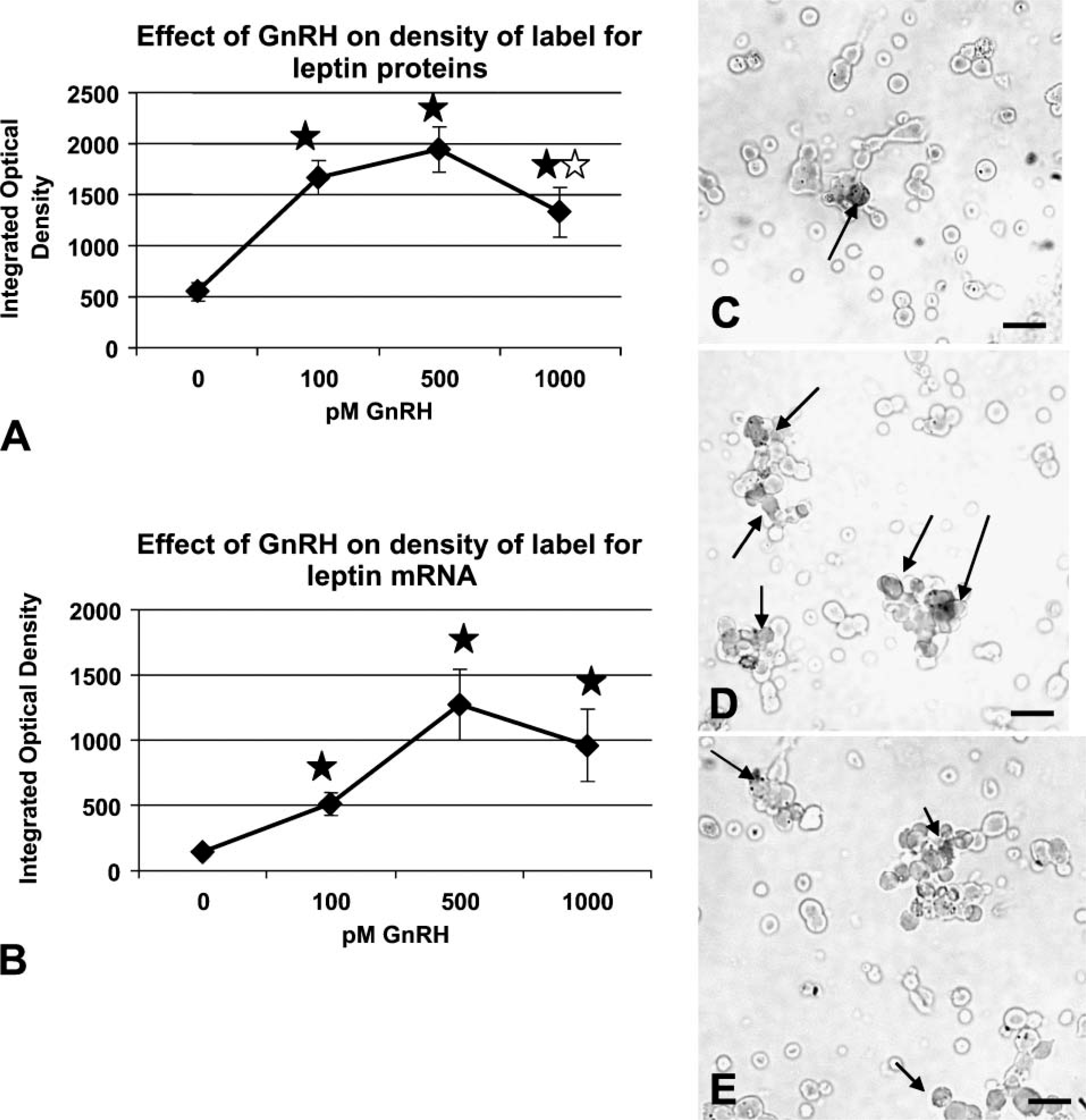
Cellular leptin proteins (
Figure 7B shows a similar response to GnRH when IOD of label for mRNA was detected. Note that the average IOD in the vehicle control is ∼5-fold lower than that for the proteins (Figure 7A). This reflects the lower expression of mRNA on the morning of proestrus seen in Figure 2. The IOD for leptin mRNA label is significantly increased in all three concentrations of GnRH; the increase with 100 pM is significant by Student's t-test (p<0.001), and there is a further increase to reach a peak with 500 pM (p<0.008). The IOD following 1 nM is not different from that with 500 pM.
GnRH also stimulated secretion of leptin from cultures of pituitary cells taken from diestrous, proestrous, or pregnant females. Figure 8 compares basal and GnRH-stimulated secretion in these groups. Basal secretion is significantly higher when one compares media from pregnant rat AP cells with that from diestrous rats (p=0.012). GnRH-stimulated secretion is increased over basal in each of the groups (diestrous p=0.016; proestrus p<0.015, and pregnant p<0.01). Cells from pregnant rats show the highest responses to GnRH when compared with all others.
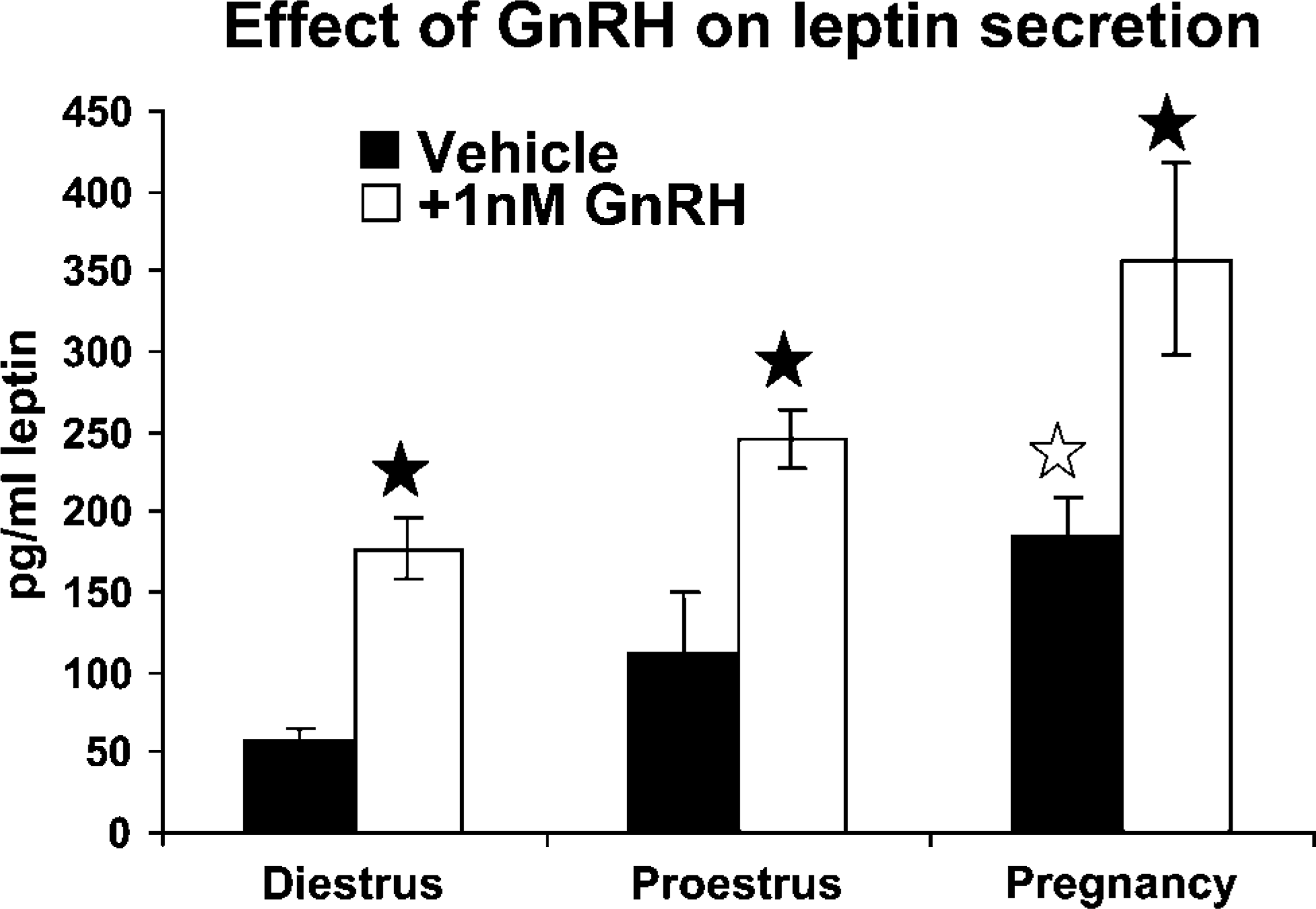
Cells from diestrous, proestrous morning, and pregnant females were treated for 1 hr with 1 nM GnRH. Media were assayed by enzyme immunosorbent assay for leptin. Basal secretion from pregnant rat cultures is higher than that from diestrous rat cultures (open star). GnRH treatment stimulated higher media levels in all experimental groups (filled stars). GnRH-treated cultures from pregnant rats have higher media levels than all other groups. See text for statistics.
Discussion
This study was designed to learn more about changes in pituitary leptin with different reproductive states. Our earlier study had shown leptin mRNA and protein expression in somatotropes and changes in protein expression with the estrous cycle (McDuffie et al. 2004). In this study we added tests of rats in proestrous afternoon and showed that this period was distinguished by continued high expression of AP leptin proteins comparable to that seen in the morning. Previous cell counts of gonadotropes did not detect major overall changes during the estrous cycle that would account for the changes in the percentages of leptin-bearing cells (Childs et al. 1992a,b). We also added tests of pregnant and lactating rats, showing increased expression of leptin in both groups. At this point, we cannot rule out the possibility of mitotic activity contributing to these changes.
As in the previous study (McDuffie et al. 2004), over two thirds of leptin-bearing cells in cycling females and normal males coexpress growth hormone. This study adds data showing that >10% of leptin-bearing cells are gonadotropes in male rats, but 14-19% are gonadotropes in female rats in diestrus or proestrus. A more striking shift is seen in AP populations from pregnant females in which 62-73% of leptin-bearing cells express FSH or LH proteins and only 19% express GH. The reason for this plasticity in expression is unclear at this point; however, some of the leptin-regulation studies discussed below may provide clues.
Leptin Regulation During Estrous Cycle
Leptin has been shown to regulate the secretion of gonadotropins, both in vivo and in vitro (Yu et al. 1997a,b; Gonzalez et al. 1999,2000; Walczewska et al. 1999; De Biasi et al. 2001). However, whereas serum leptin (presumably from adipocytes) changes during the reproductive cycle, peak levels are not seen until the luteal phase, after the LH surge (Hardie et al. 1997; Shimizu et al. 1997; Missinis et al. 1998; Riad-Gabriel et al. 1998; Geisthovel et al. 2004), suggesting postovulatory functions for serum or adipocyte leptin. This pattern does not suggest that leptin plays a major role in regulating the preovulatory LH surge, unless the rising serum leptin has a permissive effect.
In contrast, pituitary leptin appears to be differentially regulated in cycling females so that percentages of AP cells that express leptin proteins reach a peak just before the LH surge on the afternoon of proestrus. Then, the leptin-bearing cells lose stores of leptin by the morning of estrus and become invisible to immunolabeling. This suggests that the stores may have been secreted, although we cannot rule out degradation as a cause of the reduction. Pituitary leptin secretion could thus contribute to the higher serum levels seen after ovulation. Leptin-bearing cells could still be detected by their content of mRNA during estrus. Thus, the cells themselves did not disappear.
The rise in leptin protein-bearing cells seen just before the LH surge indicates that it may play an important role with respect to ovulation. A more specific paracrine role is suggested by the fact that leptin is known to be a secretagogue for LH, both in vivo and in vitro (Yu et al. 1997a,b; Finn et al. 1998; Gonzalez et al. 1999,2000; Walczewska et al. 1999; De Biasi et al. 2001). Whereas it is tempting to speculate that the rise and fall in pituitary leptin from proestrus to estrus helps support the LH surge, at this point the evidence must be considered circumstantial. Future studies would be needed to prove this paracrine function for pituitary leptin.
Leptin mRNA expression was in 32-37% of the cells early in the cycle (from estrus to diestrus). Quantification by QRT-PCR showed a dramatic rise in transcripts on diestrus morning and afternoon, 12 hr before peak expression of leptin proteins is detected. This rise during diestrus thus supports the increased proteins needed for proestrous activity. As we studied the changes percentages of cells with leptin mRNA, we noted a significant decline on the morning of proestrus, which was confirmed by the QRT-PCR assays. This rapid decline is intriguing and suggests that leptin transcripts are tightly regulated during the cycle, perhaps by rapid degradation. This pattern of expression indicates that the timing of this periovulatory expression of pituitary leptin may be important. Leptin is an anorexigenic hormone, which has potent inhibitory effects on neurons that stimulate appetite (Zhang et al. 1994; Rowland et al. 1996; Schwartz et al. 1996; Vasselli 2001). In our previous studies we suggested that pituitary leptin might be regulated to ensure adequate nutrition for pregnancy or other reproductive activities (McDuffie et al. 2004). Of greater importance in this regard may be its role in facilitating the utilization of nutrients, like glucose (Schneider and Zhou 1999; Schneider et al. 2000,2002). Thus, because of its other functions, pituitary leptin may be tightly regulated to allow it to perform supportive roles for the reproductive system, which may include both glucose utilization and LH stimulation. However, its expression may be carefully timed to prevent anorexigenic effects that might compromise a pregnancy.
Changes in leptin expression during the cycle and after pregnancy suggested that reproductive hormones like estrogen or GnRH might be involved in its regulation. Our previous studies (McDuffie et al. 2004) found that estrogen alone did not stimulate leptin in cells from metestrous rats. However, overnight treatment with estrogen followed by exposure to 2 nM growth hormone-releasing hormone (GHRH) resulted in an increase in percentages of leptin-bearing cells in vitro. In a recent publication (Childs et al. 2005), we reported an increase in GHRH receptors on GH cells following exposure to the same concentrations and times in estradiol. Thus, the stimulation of leptin seen in our first study may have been mediated by estrogen's stimulatory effects on GHRH receptors (McDuffie et al. 2004).
In the present study, the same paradigm was used because of evidence that estrogen also stimulates GnRH receptors in metestrous or diestrous rats (Lloyd and Childs 1988). In addition, we have reported that there is an increased expression of GnRH receptors by gonadotropes and somatotropes from diestrous to proestrous morning (Childs et al. 1994b). The present findings showed that cells with leptin did bind biotinylated analogs of GnRH. Furthermore, estrogen exposure to cells from diestrous females increased cells with GnRH receptors. However, estrogen did not increase the percentage of AP cells that bound GnRH and contained leptin.
GnRH Regulation of Pituitary Leptin
The biotinylated analog of GnRH detected GnRH receptors on leptin-bearing cells, which suggests that this hormone may regulate leptin directly. The counts of cells with leptin and GnRH receptors appear to have detected two times more ‘gonadotropes’ (defined by their GnRH binding) with leptin than were detected with dual immunolabeling for LH or FSH, which showed only 6 ± 1% of diestrous AP cells with leptin and LHβ and 6.6 ± 0.6% with leptin and FSHβ. This is partly explained by the appearance, in diestrus and proestrus, of somatotropes that express GnRH receptors (Childs et al. 1994b) and LH and FSH mRNA (Childs et al. 1994a). These ‘somatogonadotropes’ represent 11-16% of the AP population during proestrus. Thus, the predominance of somatotropes in the leptin-bearing cell population suggests that the GnRH might be affecting leptin from this cell type or its somatogonadotrope subtype.
To learn if estrogen and GnRH affected the gonadotropin content of leptin mRNA-bearing cells, diestrous rats were stimulated for 24 hr with estrogen and then treated for 1 hr with GnRH. In groups treated with both hormones, there were significant increases in the percentages of AP cells with leptin mRNA and LHβ or FSHβ, which shows the potential of these reproductive hormones. Neither hormone was effective by itself. At this point, we cannot rule out the possibility that the increase could have been by mitotic activity, as GnRH is mitogenic for gonadotropes (Childs and Unabia 2001).
GnRH effects on expression of cellular leptin mRNA and proteins were studied in proestrous rats, which express maximal numbers of GnRH receptors (Lloyd and Childs 1988). The average IOD array was used to integrate information from changes in density and area of label in leptin-bearing pituitary cells. There was an increase in IOD of label for leptin proteins, and mRNA in the cell populations was seen only in groups treated for 1 hr with physiological concentrations of GnRH (< 1 nM), indicating a sensitive, relatively short-term response. Groups treated for 3 hr did not respond, again suggesting that responses to GnRH may be timed, in vivo, to match its pulses. We also compared the IOD from immunolabeling with that from in situ hybridization and confirmed the significant reduction in expression of leptin mRNA seen on the morning of proestrus by the QRT-PCR assay. It is worthwhile to note, however, that the cells were still able to respond to GnRH by the production of more transcripts.
The final set of studies compared basal and GnRH-mediated secretion in cells from diestrous, proestrous, and pregnant female rats. If one compares the percentages of leptin-bearing cells in different physiological states with their secretory activity, there was also good correlation between the abundance of leptin protein-bearing cells in the population and their basal and GnRH-stimulated responses.
The possibility that leptin secretion is regulated by a neuroendocrine route suggests that this protein may have a secretory pathway distinct from that of adipocyte leptin. Insulin-mediated leptin secretion from adipocytes is considered to be via constitutive pathways, although a subpopulation of leptin vesicles may be secreted by a regulated pathway (Bradley et al. 2001). Vidal et al. (2000) detected leptin coexpression with pituitary hormones at the electron microscopic level, and their photographs depict labeling for leptin in the same granules that store LH or GH. This suggests that the leptin secretory cycle may be similar to that of gonadotropins or GH. Collectively, our experiments with GnRH and the morphological data from Vidal et al. (2000) support regulatory pathways for leptin similar to those that regulate pituitary hormones.
Plasticity in the Site(s) of Production of Pituitary Leptin
Studies of coexpression of leptin and gonadotropins or GH suggest that more leptin-bearing cells are GH cells in cycling female rats and in the male. However, in pregnancy, most leptin-bearing cells are gonadotropes. Future studies would be needed to follow the progression in leptin expression by gonadotropes with early and later pregnancy. In addition, future dual-labeling studies are needed to know if other cells contribute to the leptin-bearing population during these reproductive states.
Changes in the expression of GH proteins by AP cells does not vary significantly with the stage of the cycle (Childs et al. 2000). In this study, the counts showed 36% GH cells in diestrus and 41% in proestrus. These data agree with those from previous studies (Childs et al. 2000). Thus, the 12 percentage-point increase in leptin expression from diestrus to proestrus could be accounted for by the increase in the subset of AP cells that express GH and leptin proteins. LH- or FSH-protein-bearing cells also do not change after diestrus (Childs et al. 1987,1992a,b), and their expression of leptin also remains at 6-8% of the population.
In male rats or cycling female rats, adding the percentages of leptin cells that contain each hormone brings the values to between 87 and 103% of leptin-bearing cells. One must recognize, however, that in cycling female rats, 50-70% of gonadotropes are bihormonal (Childs et al. 1987,1994a). Furthermore, in both males and cycling females, 30-60% of GH cells coexpress one of the gonadotropins (Childs et al. 1994a). Thus, leptin could be produced by cells that also produce gonadotropins and/or GH.
Overlap in storage may also apply to the populations from lactating or pregnant rats in which adding the percentages brings the total to 130% or 156% of leptin-bearing cells. Overlap in storage of gonadotropins with or without GH has not yet been studied or described for these experimental groups; however, other ‘stimulated states’ such as castration show an increase in the proportion of bihormonal gonadotropes to nearly 100% of gonadotropes (Childs 1995,2006).
Leptin production by somatogonadotropes might help explain the plasticity evident during pregnancy, especially if the shift reflects a reduction in GH expression by this subset. Future studies of GH expression after pregnancy are needed to learn more about this phenomenon. The average percentages of GH cells in pregnant rat cultures are not lower than those normally seen in diestrous female rats.
In light of the probable overlap in storage of gonadotropins and GH in leptin-bearing cells, future studies will be needed to learn if other pituitary cell types change expression of leptin with the reproductive cycle. In our previous studies (McDuffie et al. 2004), we reviewed the literature in which dual labeling for leptin and other hormones was reported. We are the only group to report expression of leptin mRNA. However, previous investigators have reported leptin protein distribution in rodents to vary from just thyroid- stimulating hormone (TSH) cells (Jin et al. 2000) to TSH and gonadotropes, but not somatotropes (Sone et al. 2001). Overall percentages of leptin-bearing cells were lower than those reported in our studies. Studies in humans found leptin in most cell types including 70% of corticotropes, 21% of GH cells, 33% of FSH cells, 29% of LH cells, and 32% of TSH cells. Leptin was also found in 64% of folliculostellate cells but in <3% of prolactin cells. Considering the plasticity of leptin and other hormone expression in the pituitary, one would need to know more about the percentages of each of these cell types in the human and the physiological state of the donors at death to fully compare their work with the present studies. However, the broad distribution suggests that other cell types have the potential to produce leptin and could contribute to the changes in leptin-bearing cells.
To conclude, this study has demonstrated gender and cyclic differences in leptin expression, which could be regulated by GnRH supported by estrogen feedback. In females, the highest basal and GnRH-mediated leptin secretory activity was found in pituitary cells from proestrous or pregnant rats. This proves that pituitary leptin can be secreted and strengthens evidence for a role for pituitary leptin during these physiological states. Collectively, this study and our previous study (McDuffie et al. 2004) suggest that leptin expression, including secretion, may be regulated by the same neuroendocrine hormones that regulate somatotropes and gonadotropes.
Footnotes
Acknowledgements
Funding for this study was provided to G.V. Childs by the National Science Foundation (Grant IBN 0240907), National Institutes of Health (NIH; Grant R03 HD-44875), and the National Center for Research Resources (NCRR, NIH; Grant 1 P20 RR-020146). In addition, funding for overseas training was provided to Dr. Kudo by the Ministry of Education, Culture, Sports, Science and Technology of Japan for Private Universities. Contents of the study are the sole responsibility of the authors and do not necessarily represent the official views of NCRR or NIH.
The authors appreciate the help of Dr. Alex Pierson, Roche Life Sciences, with the Universal Probe system for leptin mRNA. The authors thank A.F. Parlow and the Hormone Distribution Office, National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK, NIH) for the antisera to rat growth hormone and Dr. J.G. Pierce for the anti-bovine LHβ.