
Abstract
We examined whether an enhanced green fluorescent protein (EGFP)-tagged chromogranin A (CgA) gene construct could serve as a marker protein to follow the synthesis of CgA and the process of granulogenesis in non-neuroendocrine (NE) cells. We transfected a CgA-EGFP expression vector into non-NE COS-7 cells and investigated the localization of a chimeric CgA-EGFP protein using confocal laser scanning microscopy (CLSM). The fluorescent signal of CgA-EGFP was distributed granularly in the cytoplasm. An immunocytochemical study using anti-CgA antibody with a quantum dot (Qd)525 shows colocalization of fluorescent signal of chimeric CgA-EGFP and CgA-Qd525 signals in granular structures, particularly at the periphery of the cytoplasm. We interpreted granules that were immunoreactive to CgA in electron micrographs as secretory. Spectral analysis of EGFP fluorescence revealed distinct EGFP signals without CgA colocalization. This is the first report to show that a granular structure can be induced by transfecting the EGFP-tagged human CgA gene into non-NE cells. The EGFP-tagged CgA gene could be a useful tool to investigate processes of the regulatory pathway. A more precise analysis of the fluorescence signal of EGFP by combination with the Qd system or by spectral analysis with CLSM can provide insight into biological phenomena.
Keywords
N
The accepted model of SG biogenesis comprises four distinct steps ultimately leading to the formation of MSGs. These are (1) aggregation of the regulated secretory protein (RSP) and its sorting to the membrane in the TGN, (2) budding from the TGN, (3) homotypic fusion of ISGs, and (4) remodeling of the ISG membrane and contents (Tooze et al. 2001).
Chromogranin A (CgA) is a major soluble protein of the secretory vesicles of NE cells that interacts with several other integral SG membrane proteins at pH 5.5 and dissociates from them at pH 7.5 (Yoo 1993,1994). Expression of chromogranin/secretogranin proteins is widespread in NE cells (Fischer-Colbrie et al. 1985; Simon and Aunis 1989). Among the SG-related proteins, CgA is regarded as a major marker of the NE system (Linnoila et al. 1988; Lloyd et al. 1988,1989; Eriksson et al. 1990). Chromogranins (Cgs) play not only granulogenic roles but also induce SG formation even in non-NE cells (Kim et al. 2001; Rustom et al. 2002; Huh et al. 2003) or inhibit cell growth of non-NE cells (folliculostellate cells of the pituitary gland) (Stilling et al. 2005). CgA might serve as a model for studies of proteins involved in the regulated pathway of secretion. Human CgA has a molecular mass of 49 kDa and comprises 439 amino acids. Its mRNA encodes a protein of 457 amino acids, the first 18 of which constitute the signal sequence. Overexpression of CgA induces densecore granules that are immunoreactive for CgA in non-endocrine fibroblast CV-1 cells (Kim et al. 2001).
Green fluorescent protein (GFP) has been widely applied as a tag to localize protein generated by gene transfection. It can monitor the transfection of cells as a reporter for promoter activity or localize fusion proteins in the cytosol. Fusing a RSP to a GFP variant to form a chimera should allow the study of granulogenesis in real time, i.e., granule maturation and speed of granular transport (Kaether and Gerdes 1995, 1997; Burke et al. 1997; Lang et al. 1997).
This study determines whether an enhanced GFP (EGFP)-tagged CgA gene construct could serve as a marker protein to follow the synthesis of CgA in non-NE cells via granulogenesis. We focused on granulogenesis induced in non-NE cells after transfection with a CgA-EGFP construct.
Materials and Methods
Cells
Monkey COS-7 cells were obtained from the RIKEN Cell Bank (RCB0539; Ibaraki, Japan) and maintained in DMEM (Invitrogen; Carlsbad, CA) supplemented with 10% FBS (Invitrogen) and 100 U/ml penicillin-streptomycin (Invitrogen). Cells were incubated at 37C in a humidified atmosphere (5% CO2/95% air).
Transfection of Expression Vector pCMV-CgA-EGFP
The expression vector pCMV-CgA-EGFP for chimeric protein CgA-EGFP was designed as previously described (Taupenot et al. 2002). Briefly, full-length DNA of human CgA (Helman et al. 1988) including the 18 amino acid signal peptide (MRSAAVLALLLCAGQVTA) was cloned into the pCMV expression vector (Taupenot et al. 2002). Amplified full-length human CgA (SgP-CgA) was purified and subcloned into the Xhol/KpnI site of the pCMV-EGFP (N2) vector (Clontech Laboratories; Mountain View, CA) (Figure 1). The predicted molecular mass for SgP-CgA-EGFP was 75.8 kDa as calculated molecular mass and 105 kDa as apparent molecular mass by SDS-PAGE.
COS-7 cells in Permanox chamber slides (Nalge Nunc International; Rochester, NY) or glass-bottomed culture dishes (FastGeneTM; NIPPON Genetics Co. Ltd., Tokyo, Japan) were transiently transfected with DNA (pCMV-CgA-EGFP or pCMV-EGFP) using FuGENE HD (Roche Applied Science; Indianapolis, IN) according to the manufacturer's instructions. Cells were then harvested 48 hr later and fixed.
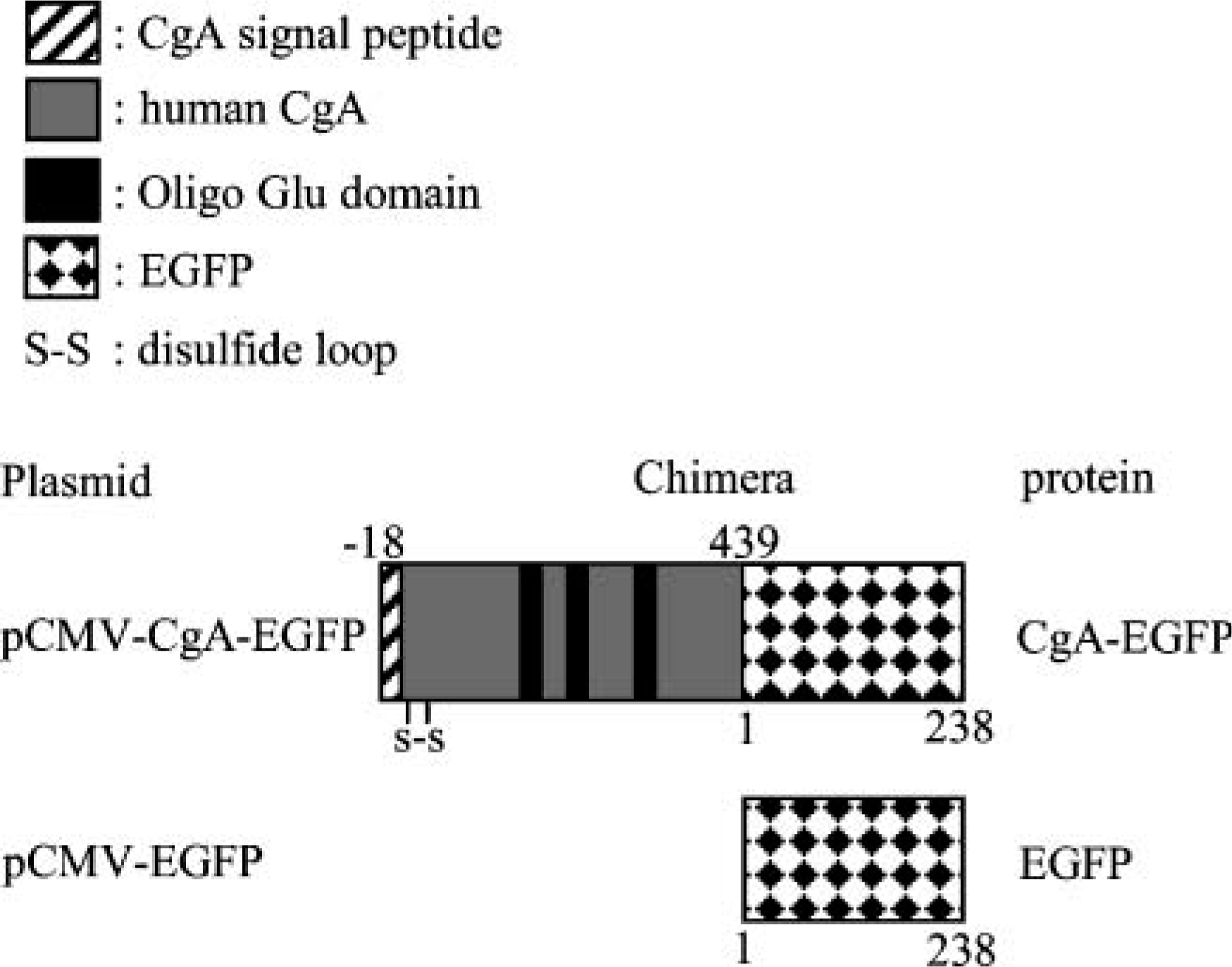
Construction of expression vector pCMV-CgA-EGFP containing full-length human chromogranin A (CgA) cDNA. Enhanced green fluorescent protein (EGFP) was fused to the C-terminus of human CgA. Numbered amino acid residue indicates position in mature protein after signal peptide cleavage.
Immunoblot Analysis
Extracted cellular proteins were resolved by SDS-PAGE on 10% polyacrylamide gels and transferred to nitrocellulose membranes (Schleicher and Schuell BioScience; Keene, NH). Nonspecific binding was blocked by incubation for 1 hr in PBS buffer containing 5% non-fat dry milk and 2% non-immune sheep serum. Membranes were then reacted with monoclonal anti-CgA, LK2H10 (Immunon; Pittsburgh, PA), or monoclonal anti-GFP, 11E5 (Molecular Probes, Invitrogen) for 1 hr and washed for 15 min with 0.05% Tween 20 in PBS. Blots were subsequently incubated for 1 hr with horseradish peroxidase-conjugated secondary antibodies (GE Healthcare UK Ltd.; Buckinghamshire, UK) in blocking buffer. Immunoreactive proteins were visualized using the ECL Plus Western Blotting Detection System as chemiluminescence (GE Healthcare U.K. Ltd.).
Electron and Immunoelectron Microscopy
Cells cultured in plastic chamber slides (Lab-Tek; Nalge Nunc International) were fixed with 2.5% glutaraldehyde in 0.1 M phosphate buffer, pH 7.4, at room temperature for 30 min, washed with buffer, and immersed in 0.1 M phosphate buffer containing 1% osmium tetroxide for 30 min. Slides were washed with buffer, dehydrated in ethanol, and embedded in epoxy resin. Ultrathin sections were attached to nickel grids that were counterstained with uranyl acetate and lead citrate. After confirming SG by electron microscopy (EM), the ultrathin sections on nickel grids were etched by incubating in 10% H2O2 for 10 min, washed with three successive droplets of 1% BSA/PBS buffer, and then incubated overnight at 4C in a humidified chamber with droplets of anti-CgA antibody (diluted 1:50, LK2H10; Immunon). After washing as described above, grids were incubated for 1 hr at room temperature with droplets of BSA/PBS buffer containing secondary antibody labeled with gold particles (5-nm average diameter). The grids were washed once with BSA/PBS and three times with distilled water and finally stained with uranyl acetate and lead citrate.
Chimeric Photoprotein Fluorescence
COS-7 cells cultured and in glass-bottomed culture dishes (FastGeneTM; NIPPON Genetics) were transfected for 48 hr with expression plasmids encoding the indicated EGFP fusion protein, fixed in 4% paraformaldehyde, and examined by confocal laser scanning microscopy (CLSM) (LSM510META; Carl Zeiss, Jena, Germany).
Immunocytochemistry
For immunocytochemistry, COS-7 cells cultured on glass-bottomed culture dishes were transfected and fixed in 4% paraformaldehyde. Cells were permeabilized with 0.4% Triton X-100 (Sigma; St Louis, MO), and nonspecific binding was blocked with 1% normal horse serum in PBS for 30 min at room temperature. A monoclonal antibody to detect human CgA (dilution 1:50, LK2H10; Immunon) and a polyclonal antibody to detect lysosomal marker, LAMP-1 (dilution 1:200, H-228; Santa Cruz Biotechnology, Santa Cruz, CA) were applied to the cells. After a PBS wash, quantum dot (Qd) (525 nm)-labeled second antibody for CgA (dilution 1:250; Invitrogen) and Qd-labeled (605 nm) second antibody (dilution 1:250; Invitrogen) for LAMP-1 were added to the cells. Slides were mounted to improve the signal-to-background ratio before visualization by an incubation for 20-30 min with ToTo-3 (dimeric cyanine nucleic acid dyes; Invitrogen) in 1,4-diazabicyclo[2.2.2]octane (DABCO; Sigma). ToTo-3 dye was diluted to 2.4 nM in PBS (200 μL) and directly applied to the slides. The slides were stored protected from light at 4C.
Spectral Analysis
Specimens were examined by computer-assisted image spectrum analysis using the LSM-510 META (Carl Zeiss) in lambda stack mode. The META analysis was arranged to ensure optimal specimen illumination and efficient collection of emitted light, which allows digital separation of the component dyes. First, a lambda stack was acquired to determine the spectral signature of each specimen by recording the spectral distribution of fluorescence emissions in 10.7-nm increments. Because spectrally resolved images are simultaneously recorded, this step is completed very quickly, which is optimal for delicate specimens, and fast dynamic processes can be reliably captured. EGFP, Qd525, and Qd605 were excited at 488 nm using an argon laser, and ToTo-3 was excited using a 633 HeNe laser.
Results
Protein Products of CgA-EGFP in Transfected COS-7 Cells Detected by Immunoblotting
Cellular expression of the chimeric protein CgA-EGFP was confirmed by immunoblotting lysates from transfected COS-7 cells with antibodies to human CgA and GFP. Figure 2 shows products of ∼105 kDa on SDS-PAGE that are consistent with the predicted mass of the full-length human CgA-GFP chimera (∼75 kDa for human CgA plus 27 kDa for GFP) (Taupenot et al. 2002). CgA from humans and other species has abnormal SDS-PAGE mobility, and its apparent molecular mass (70-75 kDa) is substantially higher than the predicted value of ∼50 kDa (Helman et al. 1988; Taupenot et al. 1995), which is a result of the highly anionic pI of CgA. A GFP-immunoreactive band was detected in extracts of pCMV-EGFP transfected COS-7 cells at ∼25 kDa (not shown). Immunoreactivity to CgA-EGFP was absent in pCMV-EGFP transfected and non-transfected cells.
Distribution of Chimeric CgA-EGFP Protein by CLSM
We designed an expression plasmid encoding the enhanced fluorescence variant of wild-type GFP fused at the N-terminus of EGFP, to the C-terminus of full-length human CgA (pCMV-CgA-EGFP; Figure 1). The single, red-shifted excitation peak of EGFP (mutations Ser65 to Thr65 and Phe64 to Leu64) fluoresces ∼35-fold more intensely than wild-type GFP when excited at 488 nm. Intracellular distribution of CgA-EGFP chimeric protein was assessed by CLSM in COS-7 cells fixed with paraformaldehyde (Figure 3). Non-fused EGFP was diffusely distributed in the cytosol of pCMV-EGFP transfected COS-7 cells (data not shown).
Signals of chimeric CgA-EGFP protein were diffuse and/or accumulating and aggregating in the cytoplasm and perinuclear area, consistent with the Golgi field (Figure 3A, green signals). Fluorescence corresponding to EGFP or chimeric CgA-EGFP was not detected in non-transfected cells (data not shown). We further investigated the intracellular localization of CgA-EGFP by immunocytochemistry using a monoclonal antibody (LK2H10) against human CgA (Figure 3B, red signals). Figure 3B shows a non-uniform punctate distribution of anti-CgA-Qd525- immunocytochemically stained signals (red) throughout but more at the peripheral region of the cytoplasm. The CgA-Qd525 signal was excluded from the nucleus. Green signals of chimeric CgA-EGFP protein and red CgA-Qd525 signals that colocalized in a granular fashion merged into yellow (Figure 3D).
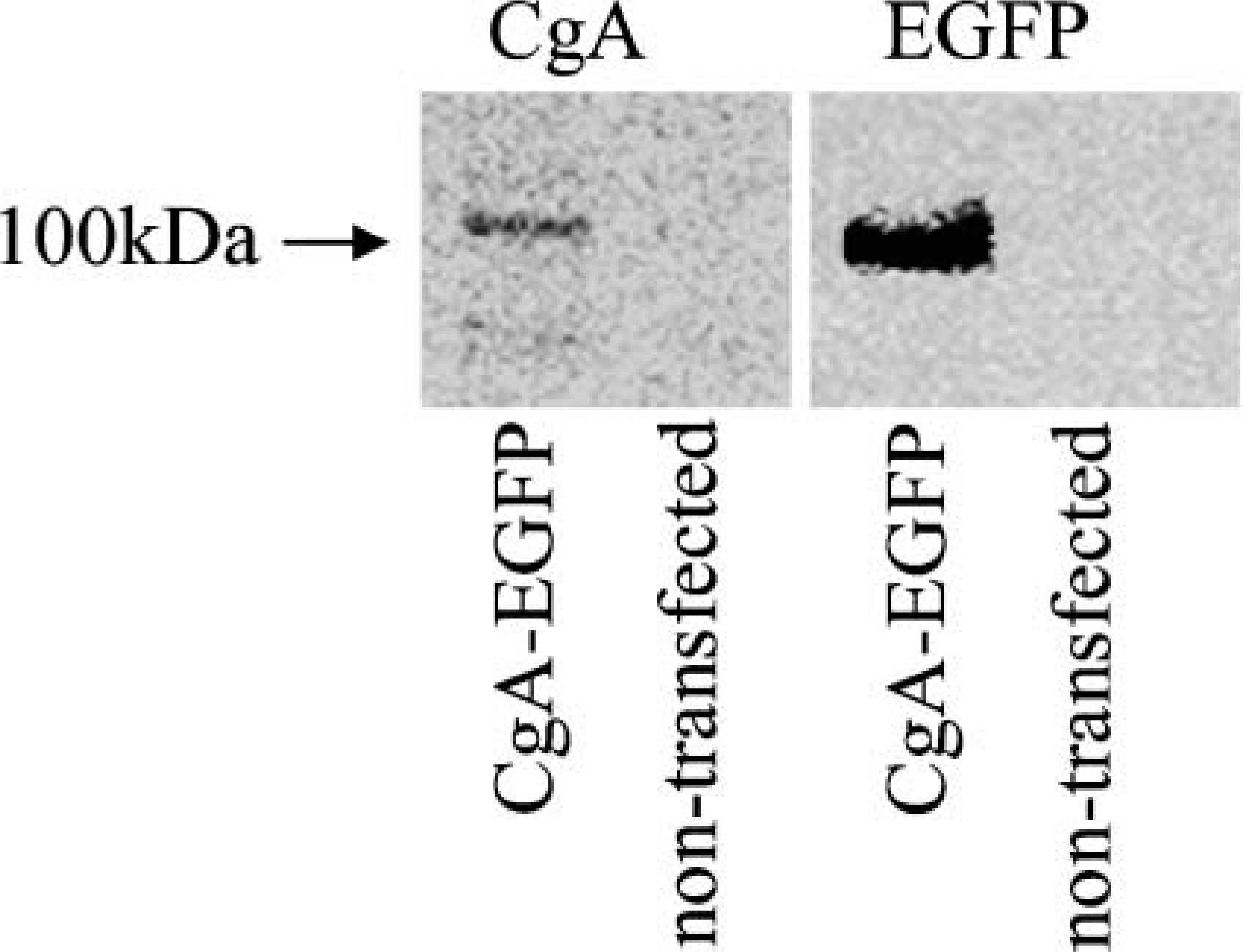
Expression of human CgA in transiently transfected COS-7 cells. Total protein extracts from the COS-7 cells transfected with pCMV-CgA-EGFP and non-transfected EGFP were resolved on 10% SDS gels and probed with anti-CgA or anti-GFP antibodies. Consistent with earlier studies (Taupenot et al. 2002), chimeric protein resolved at ∼105 kDa as reported.
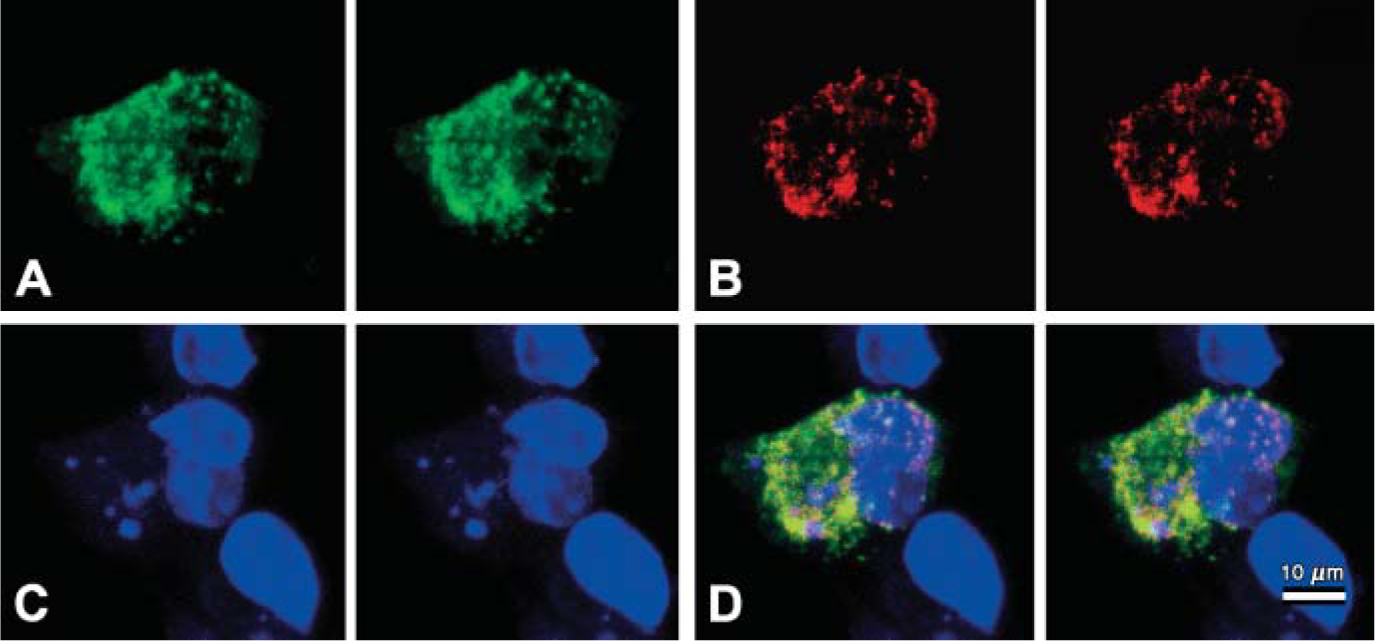
Three-dimensional META analyses in chimeric protein SgP-CgA-GFP expressed Cos 7 cells by confocal laser-scanning microscope (CLSM). Stereo pair images of CgA-EGFP signal (green color)
With CLSM we could clearly demonstrate that the signals of CgA-EGFP (Figure 4A, green in color) and LAMP-1 (Figure 4B, red in color) are mainly localized in different granules (Figure 4D).
SG Formation Identified by EM
Despite the absence of endogenous SGs in non-transfected COS-7 cells (Figure 5), COS-7 cells transfected with pCMV-CgA-EGFP contained granule-like structures (Figure 5). These structures varied in size from ∼150 to 250 nm, which was larger than the dense-core SGs in PC12 cells (Taupenot et al. 2003) or in non-NE cells (Kim et al. 2001). Granules were composed of an outer limiting membrane with core structures that were interpreted as SGs. In contrast, COS-7 cells transfected with pCMV-EGFP did not produce granular structures (data not shown). Immuno-EM analysis of human CgA in COS-7 cells transfected with pCMV-CgA-EGFP revealed the presence of several dense-core granules that were specifically stained for human CgA (LK2H10), indicating that human CgA was packaged into membrane-bound structures (Figure 5B, inset). Human CgA labeled with 5-nm gold particles was absent in mitochondria, underscoring the presence of human CgA formed de novo in the dense core granules of the cells.
Distinct Distribution of CgA and EGFP Determined by CLSM Spectrum Analysis
Three-dimensional confocal META analyses revealed individual signals from CgA-EGFP, EGFP, and CgA-QD525 in COS-7 cells transfected with pCMV-CgA-EGFP as an unmixed image. Figure 6 shows individual signals of CgA-EGFP, EGFP, and CgA-QD525 discriminated by META spectral analysis. Emission spectra of the reference signals from chimeric protein CgA-EGFP (Figure 6), EGFP (Figure 6), CgA-Qd525 (Figure 6), and ToTo-3 counterstain (Figure 6J) were defined, and all fluorescence emission spectra were merged in Figure 6K. The input images were then calculated and individual signals were separated in the liner-unmixing mode (Figures 6D-6G; CgA-EGFP chimeric protein signal, green; EGFP, yellow; CgA-Qd525, red; ToTo-3 nuclear, blue).
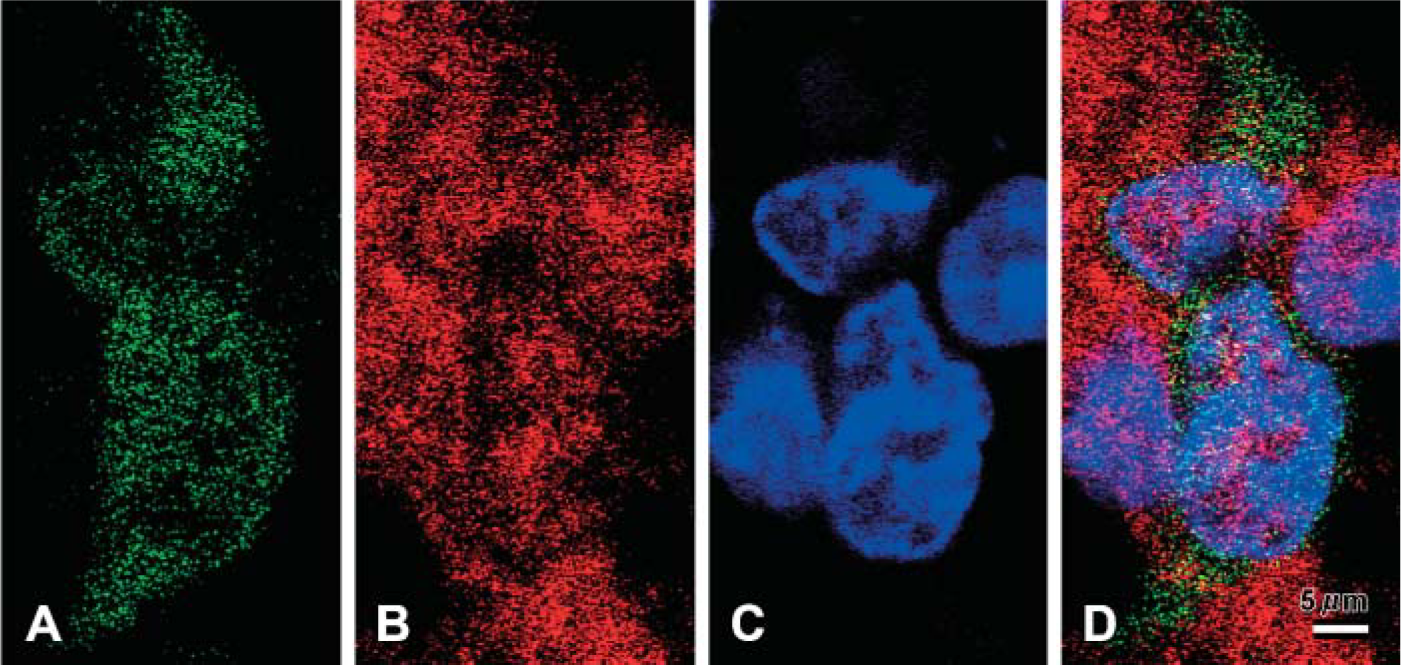
Three-dimensional simultaneous detection of CgA-EGFP and LAMP1-Qd605 in SgP-CgA-EGFP-expressed Cos-7 cells by META spectrum analysis in CLSM.
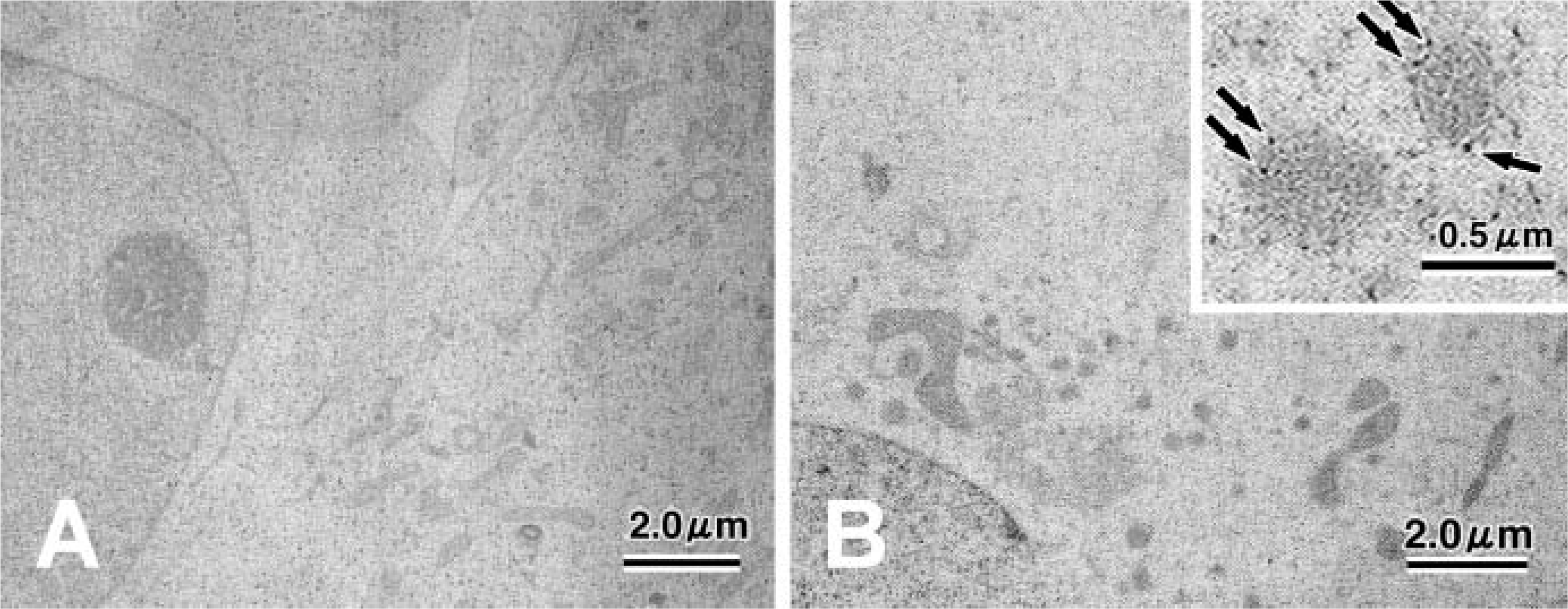
Electron and immunoelectron microscopic appearance of COS-7 cells. Non-transfected COS-7 cells do not appear to contain dense-core granules (
These results show that chimeric CgA-EGFP protein, unfused EGFP, and CgA immunoreactivity emitted distinct signals that were differently distributed for a short period. Accumulated and aggregated CgA-EGFP fluorescence signals were detected in the cytoplasm and perinuclear Golgi field (Figure 6D). Granule-like signals of EGFP were distributed mainly in the cytoplasm (Figure 6E). Fine, dotted CgA signals were mainly localized in the peripheral region excluding the Golgi field (Figure 6F). Nuclei were counterstained with ToTo-3 (Figure 6G). CgA-EGFP chimeric protein signals (green) and EGFP signals (yellow) were differently distributed. EGFP signals were mainly in the perinuclear area, whereas speckled CgA-EGFP signals were aggregated in the periphery of the cytoplasm.
Discussion
This is the first report to demonstrate granular structure produced by transfecting the EGFP-tagged human CgA gene into non-NE COS-7 cells. The role of the granin family in granulogenesis in NE and non-NE cells has been demonstrated by transfection with the CgA and CgB genes (Kim et al. 2001; Huh et al. 2003). To understand the functional role of granins and intracytoplasmic localization, GFP-tagged Cgs can be powerful tools. For example, GFP-tagged human CgB has been transfected into non-NE cells, which has led to granule formation (Kaether and Gerdes 1995; Wacker et al. 1997; Tvarusko et al. 1999). We therefore investigated the role of CgA in the process of granulogenesis in vitro.
Another striking finding was that CgA and EGFP are not always identically distributed in the cytoplasm. In cells transfected with EGFP-tagged CgA, the EGFP signal indicates CgA itself, so CgA localization has not been further examined. In the present study we investigated the localization of EGFP and CgA using an additional cytochemical technique and found that the CgA distribution did not always coincide with the EGFP signal. We found that fluorescence signals for chimeric CgA-EGFP or EGFP protein were distributed in the cytoplasm and mainly in the Golgi fields. In contrast, CgA-Qd525 signals were immunocytochemically localized mainly to the periphery of the cytoplasm (Figure 3). One explanation for this is that the fusion protein could be processed in the TGN, and only CgA lacking EGFP was packed in the mature granules.
The process of granule formation has been investigated in detail by EM. Morphology and final size of granules are diverse and depend on the cell type and granule contents. MSGs in PC12 cells are ∼120 nm, whereas insulin granules in β cells are 200 nm in diameter. The biogenesis of SGs has been thoroughly investigated in NE cells. In PC12 cells, ISGs bud from the Golgi apparatus as small granules. The diameter of ISGs in NE cells is ∼80 nm, which is the same as that of coated vesicles (Tooze et al. 1991), suggesting that coat proteins are involved in ISG budding from the TGN. Small ISGs of a similar size and character then undergo homotypic fusion (Urbe et al. 1998), which is considered necessary for SG maturation in NE cells. The present study found that SGs were between 150 and 250 nm in diameter and were consistent with the size of mature granules.
Core density of the granular structures also indicates SG maturity. Previous EM studies have shown that granules have low electron density in NE, endocrine, and exocrine cells (Arvan and Castle 1992; Tooze et al. 2001). Such low-density organelles are thought to be ISGs undergoing maturation. SGs identified in the present study were not dense compared with those of typical MSGs. We speculated that COS-7 cells transfected with CgA-EGFP cannot produce compounds such as peptide hormones in SGs; thus, SGs remained immature.
The role of CgA has been under discussion. Kim et al. (2001) reported the role of CgA as a key protein for on/off switch regulating granulogenesis by reduction and overexpression analyses of CgA. Day and Gorr (2003), however, pointed out the facts that SGs could exist in the absence of CgA and that CgA could be synthesized in the absence of SGs. They proposed the role of CgA on the condensation of SG cargo rather than a mastergene or universal on/off switch for granulogenesis. They also detected that the granular structures shown as SGs by Kim et al. (2001) were distinct from SGs, lysosomes, or endosomes. Morphological differentiation between SGs and CgA-containing lysosomes is critical in the present study. Although immuno-EM study for lysosomal protein could not be performed, we demonstrated that CgA and LAMP-1 were detected in different granules with a few exceptional colocalizations. There are a few granules containing CgA and lysosomes, but larger numbers of CgA were contained in non-lysosomal vesicles. Thus, we consider that this evidence sufficiently excludes the possibility of colocalization of CgA in the lysosome. To investigate whether or not CgA is a master gene for granulogenesis, transcription of downstream genes including secretory proteins or functional analyses is to be further examined. Nonetheless, results of the present study support the hypothesis that CgA acts to assemble or aggregate the CgA-contributing granular structure formation (Day and Gorr 2003).
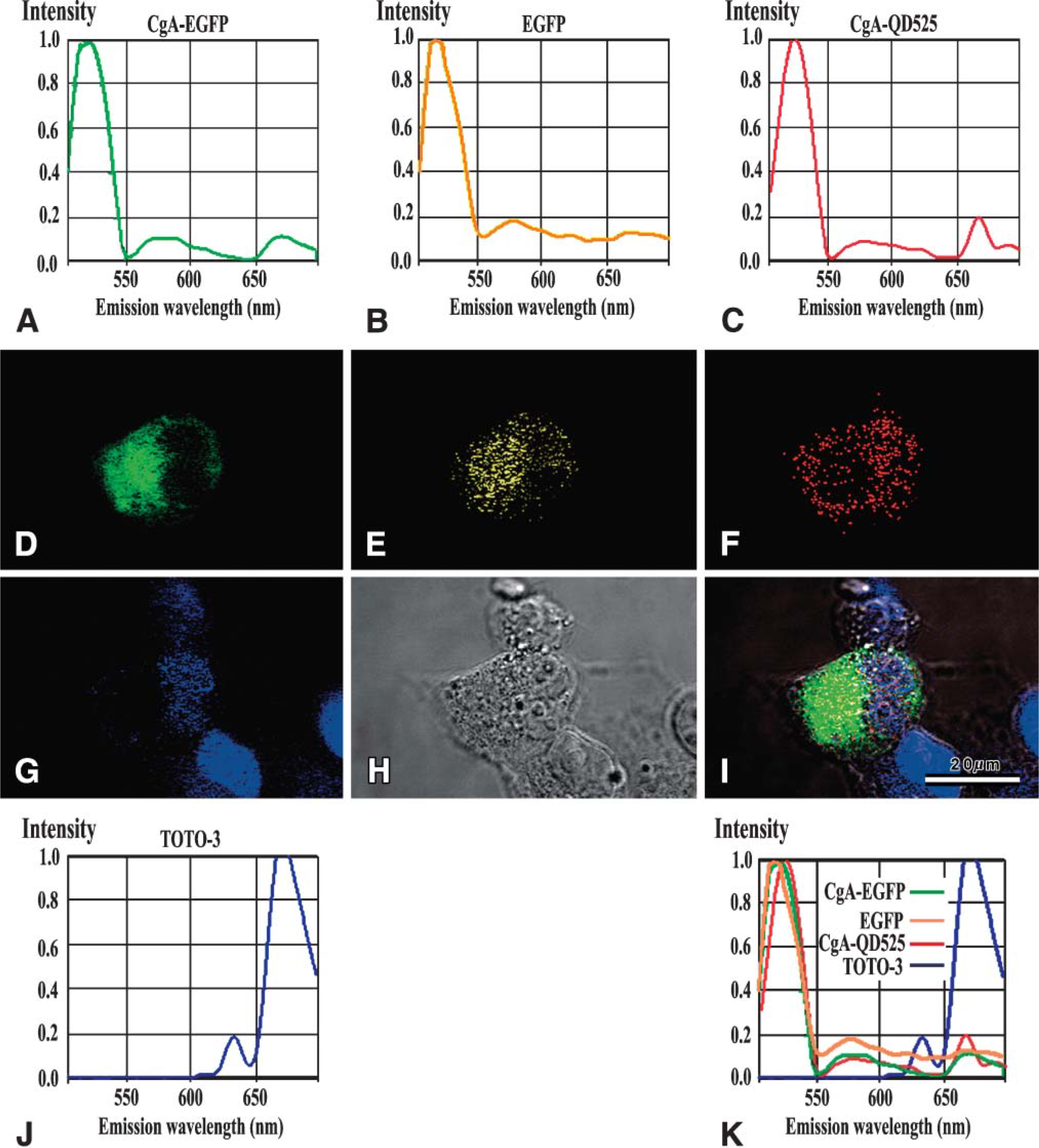
Three-dimensional signal detection of CgA-EGFP, EGFP, and CgA-Qd525 by META spectrum analysis. Spectral signatures of each fluorescence emission in META are shown in
In conclusion, EGFP-tagged CgA is a powerful tool to investigate how CgA contributes to granulogenesis.