
Abstract
Smad ubiquitin regulatory factor 2 (Smurf2) is an E3 ubiquitin ligase that is involved in the Smad-mediated TGF-β signaling. TGF-β has been shown to play an important role during normal embryo implantation, but whether Smurf2 is involved in this process has not been reported. This study was first conducted to investigate the expression of Smurf2 transcript and protein in different compartments of the rhesus monkey uteri and placenta during early pregnancy. The results showed that both the cloned partial sequence of Smurf2 gene and the corresponding amino acid residues shared 99% identity with those of human homologs. On day 12 (D12) of pregnancy, strong signals of Smurf2 mRNA were found in basalis glandular epithelium and luminal epithelium, and moderate expressions were detected in functionalis glandular epithelium. During early villi stage and villi placental stage, Smurf2 mRNAs were mainly localized in the placenta villi, trophoblastic column, trophoblastic shell, and basalis glandular epithelium. There appeared strong staining signals in the arterioles on D26 of pregnancy, but faint staining signals on D18 of pregnancy. No specific staining of Smurf2 mRNA was observed in stromal cells and myometrium. The expression pattern of Smurf2 protein was generally similar to that of its mRNA. These results provide the first evidence that Smurf2 may play specific roles in glandular secretion, trophoblastic cell invasion, and placentation through mediating the expression of the related proteins of TGF-β signaling pathway during early pregnancy.
M
Ubiquitin-mediated proteolysis regulates the activities of diverse receptor systems. Protein ubiquitination is carried out by a multienzyme cascade, E1 ubiquitin-activating enzymes, E2 ubiquitin-conjugating enzymes, and E3 ubiquitin ligases. The E3 ubiquitin ligases recruit the specific substrate to the ubiquitination machinery and subsequently the target protein is degraded by the 26s proteasomes (Hershko and Ciechanover 1998). TGF-β signaling is closely controlled by the ubiquitin-proteasome pathway. It has been demonstrated that the ubiquitin-proteasome pathway mediates the degradation of Smads (Lo and Massague 1999; Zhu et al. 1999). Smurf1 (abbreviation of Smad ubiquitin regulatory factor 1) was identified as a HECT class ubiquitin ligase that can target bone morphogenetic protein-specific Smad1 and Smad5 for degradation (Zhu et al. 1999). Smurf2, a closely related homolog of Smurf1, contains a phospholipid/calciumbinding C2 domain, three WW domains, and a HECT ubiquitin ligase catalytic domain (Kavsak et al. 2000; Lin et al. 2000; Bonni et al. 2001; Zhang et al. 2001) and has been shown to play an important role during TGF-β signaling (Zhu et al. 1999; Kavsak et al. 2000; Lin et al. 2000; Ebisawa et al. 2001; Zhang et al. 2001). Smurf2 showed a strong interaction with Smad1 and Smad2 and selectively regulated the expression of Smad2 (Lin et al. 2000) through a ubiquitination-and proteasome-dependent degradation process. In addition, Smurf2 can interact with a Smad transcriptional corepressor SnoN (ski-related novel gene N) that targets them for ubiquitination and degradation (Bonni et al. 2001).
Smurf2 was identified in a variety of tissues and cell lines such as testis, placenta, and ovarian cancer cell lines and exerts various effects on many biological processes such as development, senescence, tumor formation, and disease progression (Kavsak et al. 2000; Zhang et al. 2001; Fukuchi et al. 2002; Zhang and Cohen 2004; Kuratomi et al. 2005). However, little is known regarding the role of Smurf2 during mammalian early pregnancy. In addition, numerous studies indicate that the ubiquitin-proteasome pathway and TGF-β are involved in the development of the decidua and are also necessary for embryo implantation, normal placental growth, and development during pregnancy (Austin et al. 1996; Harbers et al. 1996; Bebington et al. 2001; Wang et al. 2004a; Jones et al. 2006). It is possible, therefore, that Smurf2 as an E3 ubiqiutin ligase may be involved in some biological events during early pregnancy via regulating TGF-β signaling.
For the first time, we studied the spatiotemporal expression of Smurf2 in the endometrium and placenta during early pregnancy by using the rhesus monkey early pregnancy model.
Materials and Methods
Animals and Tissue Collection
Uteri of 14 rhesus monkeys (Macaca mulatta) with records of normal menstrual cycles and pregnancy were obtained from the Center of Medical Primates, Institute of Medical Biology, Chinese Academy of Medical Sciences. All experimental protocols were in accordance with the policy on the Care and Use of Animals of the Ethical Committee, State Key Laboratory of Reproductive Biology, Institute of Zoology, Chinese Academy of Sciences. The length of gestation of the rhesus monkey is ∼165 days. Female rhesus monkeys were caged with proven fertile male monkeys for 2 days from the anticipated day of ovulation [about day 12 (D12) of the menstrual cycle], considered as D0 of pregnancy. Pregnancy prediction was determined by the presence of vaginal sperm plug, monkey chorionic gonadotropin (mCG) detection in the urine, and ultrasound examination (Wang et al. 2001). On estimated D12 (n=4), D18 (n=4), D26 (n=4), and D35 (n=2) of pregnancy, monkeys were anesthetized by ketamine hydrochloride and the uteri removed surgically. Uteri were fixed in Bouin's solution overnight and embedded in paraffin.
Rhesus Monkey Smurf2 Gene Fragment Cloning
Total RNAs were prepared from uteri (n=3) using Trizol reagent (Gibco BRL; Gaithersburg, MD) according to the manufacturer's instructions. Two μg of total RNA was used to synthesize the first-strand cDNA with SuperScript II reverse transcriptase (Gibco) and oligo dT (SBS Genetech Co., Ltd.; Beijing, China). PCR reaction (total volume 25 μl) was carried out using 2 μl of the first-strand cDNA as templates and Taq DNA polymerase (TaKaRa; Dalian, China). Sense and antisense primers for Smurf2 are 5î AGGCTCAATTCTTGGCTCTG 3î and 5î ATGAAGAGCGACGAGCAAGA 3î, respectively, giving a 227-base pair (bp) PCR product. PCR reaction was done with an initial denaturation step at 95C for 5 min, 94C for 45 sec, 57C for 45 sec, and 72C for 45 sec, followed by 30 cycles and a final extension step at 72C for 7 min. The amplified Smurf2 cDNA fragment containing the expected size was cut and purified using the Nucleo Trap Gel Extraction Kit (Clontech; Palo Alto, CA). cDNA was cloned into the pGEM-T vector (Promega; Madison, WI) according to manufacturer's instructions and sequenced commercially (Sangon Corp.; Shanghai, China).
Preparation and Labeling of cRNA Probes
Generation and labeling of probes were described in our previous work (Yang et al. 2006). Briefly, the 227-bp fragment from the rhesus monkey Smurf2 sequence was cloned and inserted into the pGEM-T vector as described above. To prepare the antisense and sense digoxigenin (DIG)-labeled Smurf2 probes, the plasmid was then linearized using bipartite primers consisting of either a modified T7 RNA promoter and the downstream gene-specific sequence (antisense) or a modified SP6 RNA promoter and the upstream genespecific sequence (sense). PCR reactions were carried out as mentioned above. Transcripts at expected size from the pGEM-T-mSmurf2 plasmid were purified using Nucleo-Trap Gel Extraction Kit (Clontech). Antisense and sense probes were synthesized using T7 and SP6 RNA polymerases (Promega), respectively.
In Situ Hybridization
In situ hybridization (ISH) protocol was detailed previously (Wang et al. 2004b). Briefly, paraffin sections were cut serially at a thickness of 5 μm and mounted on APES-coated (Zhongshan Corp.; Beijing, China) slides. After being dewaxed completely, sections were digested with proteinase K solution (10 μg/ml) for 10 min at 37C and fixed in 4% paraformaldehyde for 5 min at 4C followed by prehybridization in prehybridization solution containing 50% deionized formamide, 10% dextran sulfate (w/v), 0.5% SDS, 1X Denhardt, 10 mmol/liter Tris-Cl (pH 8.0), 10 mmol/liter DTT, 2X SSC (1X SSC is 0.15 M NaCl, 0.015 M sodium citrate), and 250 μg/ml sheared salmon sperm DNA for 2 hr at 50C and hybridization with 400 ng/ml denatured probes in prehybridization solution overnight at 50C. Slides were then serially washed in 2X SSC, 1X SSC, and 0.1X SSC at 42C, equilibrated in buffer A (100 mM Tris, 150 mM NaCl, pH 7.5) for 5 min and blocked with blocking solution (buffer A containing 1% blocking reagent, Roche; Mannheim, Germany) followed by incubation with anti-DIG-alkaline phosphatase (diluted 1:500 in blocking solution) at room temperature for 2 hr. The slides were rinsed in buffer A three times for 10 min each. Color development was performed using nitroblue tetrazolium/5-bromo-4-chloro-3-indolyl phosphate (NBT-BCIP; Boehringer-Mannheim, Mannheim, Germany) in buffer B (100 mM Tris, 100 mM NaCl, 50 mM MgCl2,pH9.5). Levels of Smurf2 mRNA staining detected by ISH were subjectively assessed by three independent investigators. Senseprobe hybridization was used as a control for background level.
Immunohistochemistry
Immunohistochemical staining was performed using streptomycin avidin-peroxidase immunochemistry kit (Zhongshan Corp.). Five-μm-thick paraffin-embedded sections were de-paraffinized with xylen, and rehydrated through a graded series of ethylalcohol and PBS. To expose the antigen sufficiently, slides were heated in a microwave oven at 92-100C for 15 min in citrate buffer (10 mM, pH 6.0), cooled to room temperature, quenched in 3% H2O2 for 10 min to block endogenous peroxidase, and washed in PBS. The slides were then incubated with normal blocking reagent for 20 min and an anti-Smurf2 polyclonal antibody (diluted 1:200, cat. #07-249; Upstate Biotechnology, Lake Placid, NY) for 2 hr at room temperature. After washing with PBS, sections were treated with biotinylated secondary antibody for 15 min and avidin-conjugated horseradish peroxidase for another 15 min. Color development was achieved by incubating in a diaminobenzidine (DAB; Zhongshan Corp.) solution. Slides were counterstained with hematoxylin and mounted with histomount mounting solution (Zymed; San Francisco, CA) for microscopic examination and photography. A negative control was prepared by substituting normal rabbit serum for the primary antibody. Staining intensity on the slides was subjectively assessed by three independent investigators.
Results
In Situ Localization of Smurf2 mRNA in the Endometrium and Placenta of Rhesus Monkey During Early Pregnancy
To examine the spatiotemporal expression of the Smurf2 mRNA in the pregnant monkey uterus using ISH, we first obtained one partial fragment of Smurf2 from the monkey uterus using RT-PCR. The sequence of Smurf2 PCR products is shown in Figure 1A. This partial cDNA fragment of rhesus monkey containing 227 bp shared 99% sequence similarity with the corresponding region of human Smurf2 gene (nt 1972-2198, GenBank accession no. AY014180). The corresponding nucleotide sequence encodes 75 amino acid residues (amino acids 1-75) with 99% sequence identity with its human homolog (amino acids 595-669), which is part of the C-terminal Hect domain. Only one amino acid is different between the human and monkey in protein sequences (Figure 1).
Results from ISH are shown in Figure 2 and Table 1. Smurf2 mRNA was spatiotemporally expressed in the endometrium and placenta of rhesus monkey during early pregnancy. On D12 of pregnancy, Smurf2 mRNAs were localized mainly in the glandular epithelium and luminal epithelium, but the staining signals in functionalis glandular epithelium were lower than those in basalis glandular epithelium (Figures 22-A2). During early villi stage (D18 of pregnancy, Figures 2D-2F) and villi placenta stage (D26 of pregnancy, Figures 2G-2I; D35 of pregnancy, Figures 2J and 2K), Smurf2 mRNAs were highly expressed in the placental villi, trophoblastic column, and basalis glandular epithelium and moderately detected in the trophoblastic shell (Figures 2E, 2H, and 2K). There was a faint staining signal in the arteriole on D18 of pregnancy (Figure 2F), whereas a strong staining signal appeared on D26 of pregnancy (Figure 2I). Very faint or undetectable staining was observed in the myometrium and stromal cells of all samples examined. Control slides hybridized with sense probe showed nonspecific signals (Figure 2L).

Comparison of the 227-nucleotide fragment and the corresponding amino acid residue sequence between human and rhesus monkey. (
Semiquantification of Smurf2 mRNA at different locations of rhesus monkey endometrium and placenta on days 12 (D12), D18, D26, and D35 of pregnancy
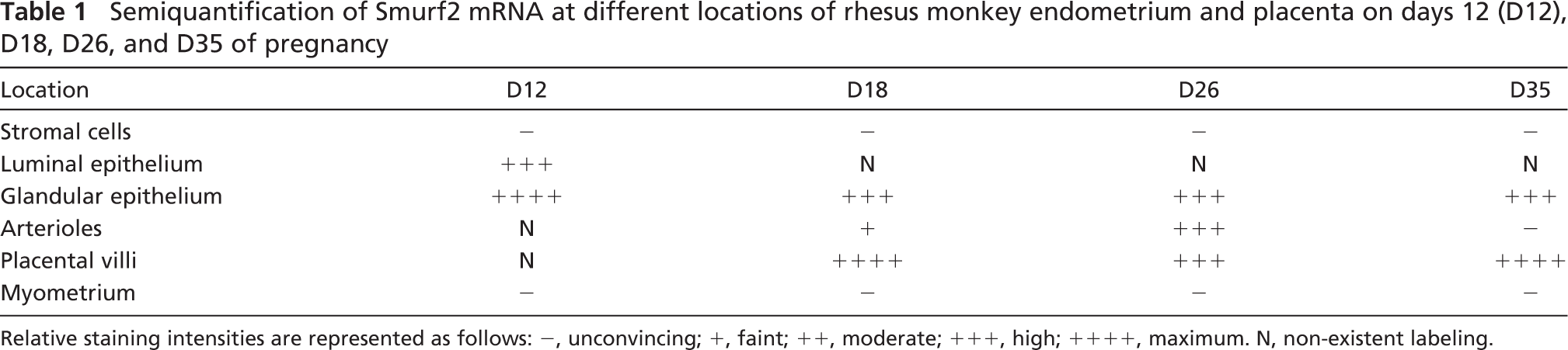
Relative staining intensities are represented as follows: −, unconvincing; +, faint; ++, moderate; +++, high; ++++, maximum. N, non-existent labeling.
Immunohistochemical Localization of Smurf2 Protein in the Endometrium and Placenta of Rhesus Monkey During Early Pregnancy
The partial nucleotide sequence of Smurf2 shared high similarity between rhesus monkey and human as shown in Figure 1B. To further understand the possible role of Smurf2 protein in placentation and endometrial remodeling during early pregnancy, the commercial antibody of Smurf2 that recognizes human Smurf2 was used to examine the localization of rhesus monkey Smurf2 protein by using immunohistochemistry, as shown in Figure 3 and Table 2. Similar to that of its mRNA, Smurf2 protein localization also showed a spatiotemporal distribution pattern in the macaque endometrium and placenta during early pregnancy, and the localization of Smurf2 protein was generally similar to that of its mRNA. On D12 (Figures 3A and 3B) of pregnancy, there were strong staining signals in the basalis glandular epithelium and moderate staining in the luminal epithelium. During early villi stage (D18 of pregnancy, Figures 3C-3F) and villi placenta stage (D26 of pregnancy, Figures 3G-3I; D35 of pregnancy, Figures 3J and 3K), staining signals of Smurf2 protein were found mainly in the placenta villi, trophoblastic column, trophoblastic shell, arteriole, and the basalis glandular epithelium (Figures 33-C3). Strong staining signals can be found in both the syncytotrophoblastic cells and cytotrophoblastic cells (Figures 3F and 3I). Myometrium and stromal cells showed faint or undetectable staining signal. In addition, there were no positive staining signals in the control sections, substituting the primary antibody with normal rabbit IgG (Figure 3L).
Discussion
Smurf2 is a member of the Hect family of E3 ubiquitin ligases that regulates various Smads and TGF-β receptor complex via a ubiquitin-proteasome pathway (Kavsak et al. 2000; Lin et al. 2000; Zhang et al. 2001). Smurf2 exerts various functions in many biological processes such as development, senescence, tumor formation, and disease progression (Kavsak et al. 2000; Zhang et al. 2001; Fukuchi et al. 2002; Zhang and Cohen 2004; Kuratomi et al. 2005). It was shown that Smurf2 mRNA was expressed in human placenta. However, localization of Smurf2 has not been elucidated during early pregnancy.
As many of the aspects of primate embryo development are similar to those of human embryos, the non-human primate represents an ideal model for understanding basic human biology in early pregnancy due to the limited access to early human pregnant specimens. Our present study showed that both the Smurf2 gene fragment and amino acid residues of the rhesus monkey had high sequence identity with the corresponding human homologs, and the transcript and protein of Smurf2 were found mainly in the endometrial epithelia and placenta during rhesus monkey early pregnancy. These results are helpful to explore the possible roles of Smurf2 during human early gestation.
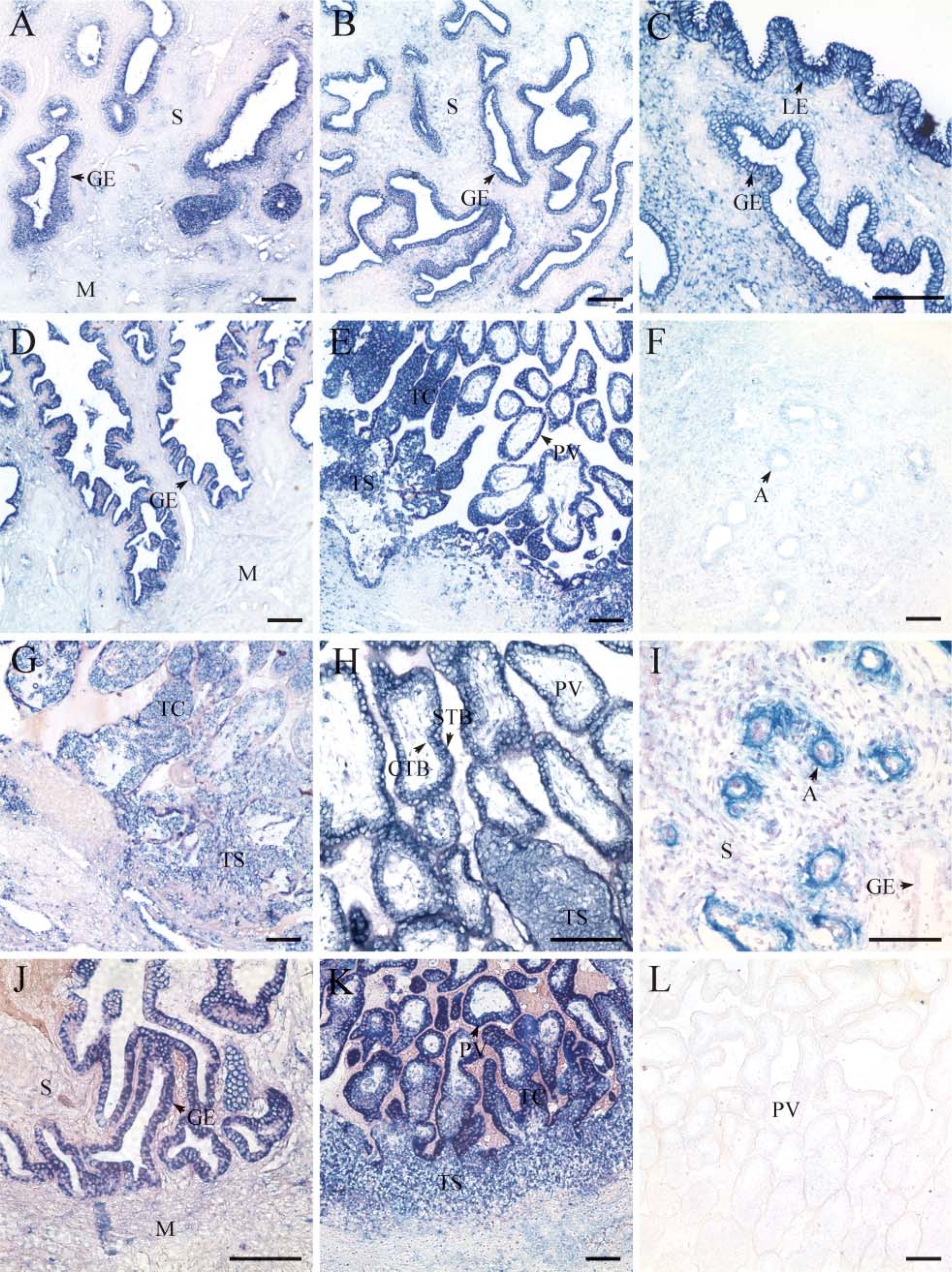
In situ hybridization showing spatial and temporal distribution pattern of Smurf2 mRNA in the rhesus monkey endometrium and placenta on days 12 (D12) (
Semiquantitative analysis of Smurf2 protein at different compartments of rhesus monkey endometrium and placenta on D12, D18, D26, and D35 of pregnancy

Relative staining intensities are represented as follows: −, unconvincing; +, faint; ++, moderate; +++, high; ++++, maximum. N, non-existent labeling.
Secretions of endometrial glands are very important for embryogenesis, embryo implantation, and placentation (Gray et al. 2001). Evidence from our and other laboratories has documented the epithelial localization of TGF-βs, TGF-β receptors, and Smads in pregnant endometrium in many species (Tamada et al. 1990; Das et al. 1992; Lennard et al. 1995; Lin et al. 2004,2006; Liu et al. 2004). The present data demonstrated that a high level of Smurf2 was also localized to the luminal epithelium and glandular epithelium adjacent to the myometrium during rhesus monkey early pregnancy. As Smurf2 can interact with Smads protein and is involved in the Smad-mediated TGF-β signaling (Kavsak et al. 2000; Lin et al. 2000; Bonni et al. 2001; Zhang et al. 2001), the epithelial localization of Smurf2, together with members of TGF-β signaling pathway, may suggest that Smurf2 functions to regulate TGF-β signaling through interacting with epithelial Smads and TGF-β receptors, thus participating in the establishment of uterine epithelial morphology and maintaining the normal function of epithelial cells during pregnancy.
The invasion of human placental trophoblastic cells shares many identities with that of cancer cells; however, unlike cancer cells, their proliferation, migration, and invasion are strictly controlled by TGF-β derived from decidua and trophoblast (Graham et al. 1992,1994; Irving and Lala 1995). TGF-β signaling pathway exerts its functions predominantly through Smad family. Smads are expressed in human trophoblastic cells and are active in response to TGF-β (Wu et al. 2001). Loss of TGF-β responsiveness is a phenotype shared by premalignant and malignant extravillous trophoblastic cells (Graham et al. 1994; Khoo et al. 1998). Smurf2 mRNA shows a high expression in human placenta (Kuratomi et al. 2005). In the present study we detected expression of Smurf2 in the placental compartments from early villi stage and villi placental stage. Our results showed that strong staining signals were observed in the trophoblastic columns and placental villi during monkey early pregnancy. These findings hint that Smurf2 might be involved in regulating the trophoblast invasion and placentation stringently through targeting the related proteins of the TGF-β signaling pathway and mediating their proteasomal degradation.
Smurf2 mRNAs were highly expressed in some arterioles on D26, and the protein of Smurf2 can be detected in the arterioles of all samples. The presence of Smurf2 in arterioles indicates that it may play a role in angiogenesis.
In summary, we report for the first time the spatiotemporal expression of Smurf2 in rhesus monkey endometrium and placenta during early pregnancy. The specific expression profile of Smurf2 suggests that Smurf2 may be involved in regulating glandular secretion, trophoblastic cell invasion, and placentation through mediating the expression of the related proteins of TGF-β signaling pathway during early pregnancy. To better understand the role of Smurf2 during early pregnancy, further investigation is necessary to elucidate the colocalization of Smurf2 and Smads.
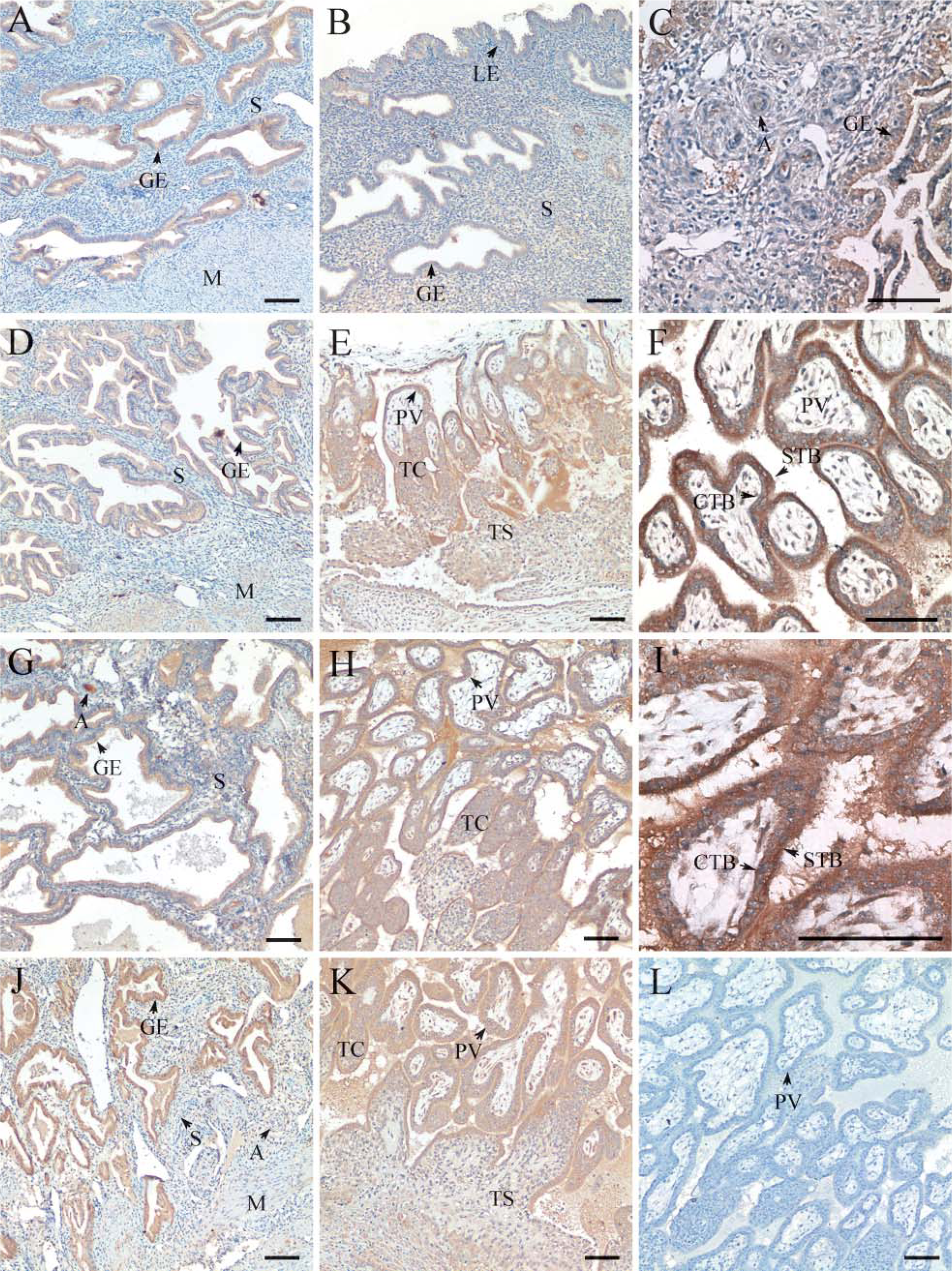
Immunohistochemical staining for Smurf2 protein in rhesus monkey endometrium and placenta on D12 (
Footnotes
Acknowledgements
This work was supported by the 973 Program (MOST; 2006CB504006 and 2006CB944008, to H-MW), the Knowledge Innovation Project of the Chinese Academy of Sciences (KSCX3-IOZ-07), and funds from the National Natural Science Foundation of China (30670262).