
Abstract
Juvenile hemochromatosis is a severe hereditary iron overload disease caused by mutations in the HJV (hemojuvelin) and HAMP (hepcidin) genes. Hepcidin is an important iron regulatory hormone, and hemojuvelin may regulate hepcidin synthesis via the multifunctional membrane receptor neogenin. We explored the expression of murine hemojuvelin and neogenin mRNAs and protein. Real-time RT-PCR analysis of 18 tissues from male and female mice was performed to examine the mRNA expression profiles. To further study protein expression and localization we used immunohistochemistry on several tissues from three mouse strains. Mouse Neo1 mRNA was detectable in the 18 tissues tested, the highest signals being evident in the ovary, uterus, and testis. Neogenin protein was observed in the brain, skeletal muscle, heart, liver, stomach, duodenum, ileum, colon, renal cortex, lung, testis, ovary, oviduct, and uterus. The spleen, thymus, and pancreas were negative for neogenin. The highest signals for Hjv mRNA were detectable in the skeletal muscle, heart, esophagus, and liver. The results indicate that Neo1 mRNA is widely expressed in both male and female mouse tissues with the highest signals detected in the reproductive system. Moreover, Hjv and Neo1 mRNAs are simultaneously expressed in skeletal muscle, heart, esophagus, and liver.
H
Hepcidin is an antimicrobial peptide and iron regulatory hormone. It binds to the iron exporter ferroportin and induces its internalization from the cell surface and subsequent degradation (Nemeth et al. 2004). Ferroportin is expressed in hepatocytes, reticuloendothelial system cells, and in the basolateral membrane of duodenal enterocytes (Abboud and Haile 2000). Thus, hepcidin negatively regulates the rate of intestinal iron absorption and, in general, of iron efflux into the plasma. The low production of hepcidin observed in HH patients (see above) can therefore explain the high levels of intestinal iron absorption and high transferrin saturation; hence, the overall HH phenotype. Moreover, HH-related HAMP downregulation indicates that HFE protein, transferrin receptor-2, and hemojuvelin are upstream regulators of HAMP, although the underlying pathways remain obscure.
Hemojuvelin (HJV, HFE2, or RGMc) is a glycosyl phosphatidylinositol-anchored protein for which five spliced variants have been predicted, encoding proteins of 200, 313, and 423 amino acids (Papanikolaou et al. 2004; Zhang et al. 2005). To date, there is no clear evidence for the production of the two shorter peptides, and new findings suggest that under certain conditions the full-length HJV is posttranslationally processed to yield a shorter N-terminal and a larger C-terminal product (Zhang et al. 2005). Of additional importance, a recent study has revealed that hemojuvelin can be found in the human body in two forms: membrane associated and soluble hemojuvelin proteins. These two hemojuvelin types appear to reciprocally regulate HAMP expression, in response to extracellular iron concentrations in human primary hepatocytes, by physically competing for a receptor-binding site (Lin et al. 2005). Interestingly, another recent report has shown evidence for a physical association between hemojuvelin and the membrane receptor neogenin [but not its homolog, Deleted in Colorectal Cancer (DCC)] in cultured embryonic kidney cells. This same study demonstrated that the hemojuvelinneogenin interaction is required for iron accumulation is these cells (Zhang et al. 2005). Based on these new findings, a logical hypothesis would be that hemojuvelin exerts its HAMP regulatory role through its receptor, neogenin.
Neogenin is a type I transmembrane receptor that is homologous to DCC, and both constitute a subfamily within the N-CAM family of cell adhesion molecules. The extracellular regions of neogenin and DCC contain four immunoglobulin domains with disulfide-bonded cysteines and six type-III fibronectin repetitions. The cytoplasmic domains of these two receptors have no homology to any other known proteins (Meyerhardt et al. 1997). Diverse functions of neogenin upon interaction with different ligands have been reported, such as the repulsive guidance of retinal axons and the regulation of neuronal survival via the interaction with chick RGM (repulsive guidance molecule) (Monnier et al. 2002; Matsunaga et al. 2004). Another function of neogenin is its role in myotube formation, mediated by its binding to netrin-3 (Kang et al. 2004).
It is significant that the binding ligands of neogenin are expressed in quite distinct locations (Schmidtmer and Engelkamp 2004; Barallobre et al. 2005), further suggesting that it may act as a multifunctional receptor whose role is determined by the presence of specific interacting molecules. In our present study in the mouse, we elucidate sites of simultaneous expression of Neo1 and Hjv mRNA. We also explore the spatial distribution of the murine neogenin and hemojuvelin proteins in different tissues.
Materials and Methods
RNA Extraction and First-strand cDNA Synthesis
Tissue samples for mRNA quantification were obtained from 10-week-old NMRI mice (four males and four females) with the approval of the Animal Care Committee of the University of Tampere, Tampere, Finland. The tissues extracted included the lung, brain, muscle, heart, spleen, thymus, pancreas, liver, esophagus, stomach, duodenum, jejunum, ileum, colon, kidney, testis, ovary, and uterus. All specimens used for quantitative real-time PCR were snap frozen upon extraction and stored at −80C until use. Total RNA was isolated using TRIZOL reagent (Invitrogen; Carlsbad, CA) according to the manufacturer's instructions. After digestion with RNase-free DNase I (Novagen; Madison, WI), the resulting RNA samples were further purified using phenol/chloroform, followed by precipitation with ice-cold ethanol. RNA concentration and purity were determined in each case by optical density measurements at 260 and 280 nm. RNA extracts from males and females were separately pooled to reduce the potential for individual variation. Three μg of each total RNA isolate was converted into first-strand cDNA with a First Strand cDNA Synthesis kit (Fermentas; Burlington, Canada) and random hexamer primers, according to the protocol recommended by the manufacturer.
Quantitative Real-time PCR
The relative levels of mouse Neo1 and Hjv transcripts in different tissues were assessed by real-time PCR using the Roche LightCycler detection system (Roche; Rotkreuz, Switzerland). The primer sets for the target genes used in these analyses were designed using Primer3
Primer sequences for internal control genes

Designed using Primer3 (
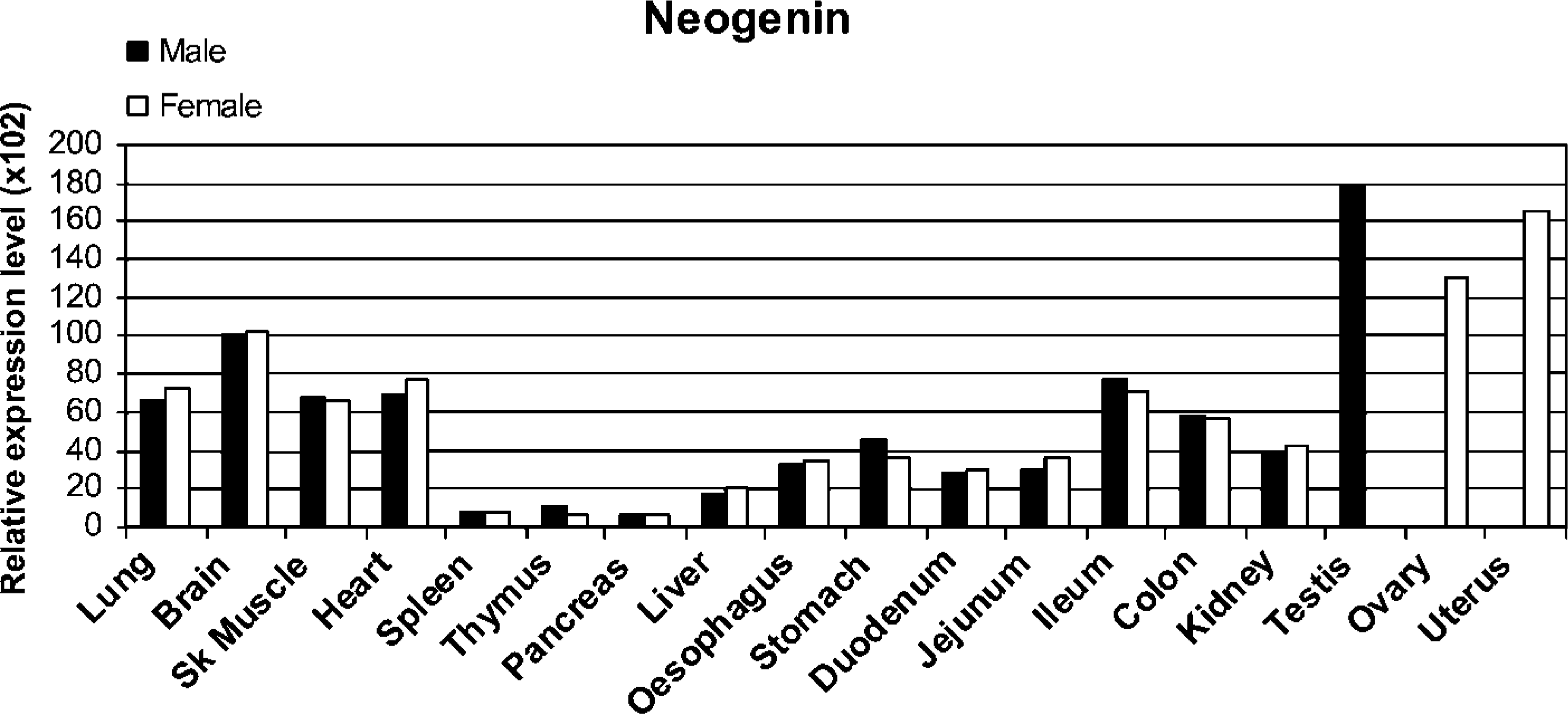
Real-time RT-PCR analysis of Neo1 in mouse tissues. Data shown are the relative expression levels after normalization with four internal control genes (Bact, Gapdh, Hprt1, and Sdha).
The Actb (β-actin), Gapdh (glyceraldehyde-3-phosphate dehydrogenase), Hprt1 (hypoxanthine phosphoribosyl-transferase I), and Sdha (succinate dehydrogenase complex subunit A) genes were used as internal controls to normalize for potential quality and quantity differences between samples. The primers for the internal controls are shown in Table 1. Every PCR reaction was performed in a total volume of 20 μl containing 1 μl of first-strand cDNA, 1X concentrated QuantiTect SYBR Green PCR Master Mix (Qiagen; Hilden, Germany), and 0.5 μmol/liter of each primer. Amplifications and subsequent detection were carried out as described. After an initial activation step of 15 min at 95C, amplification was performed in a three-step cycling procedure: denaturation at 95C, 15 sec, ramp rate 20C/sec; annealing according to the melting temperature of the primers, 20 sec, ramp rate 20C/sec; and elongation at 72C, 20 sec, ramp rate 20C/sec for 45 cycles, and a final cooling step. The melting curve analysis was always performed for each PCR amplicon to verify specific amplification.
To quantify the concentration of the Actb, Gapdh, HprtI, Sdha, Neo1, and Hjv transcripts in the studied tissues, a standard curve for each gene was established using 5-fold serial dilutions of known concentrations of purified PCR products generated from the same primer sets. Every cDNA sample was tested in duplicate and the obtained crossing point (Cp) value facilitated the determination of the levels of starting message using a specific standard curve. The geometric mean of the four internal control genes was used as an accurate normalization factor for gene expression levels (Vandesompele et al. 2002). The level of expression of the target gene in each tissue was divided by the corresponding normalization factor and subsequently multiplied by 102.
Immunohistochemistry
Tissue specimens were obtained from eight adult mice including four NMRI (two male and two female), two Balb/c (male and female), and two C57 (male and female) mice. All available samples were used for neogenin studies, whereas hemojuvelin staining was performed on samples from female C57 mice. After extraction, samples were fixed in 4% neutral-buffered formaldehyde at 4C for 24 hr and then dehydrated in an alcohol series, treated with xylene, and embedded in paraffin wax. Four-μm sections were cut and placed on SuperFrost Plus (Menzel-Glaser; Braunschweig, Germany) microscope slides.
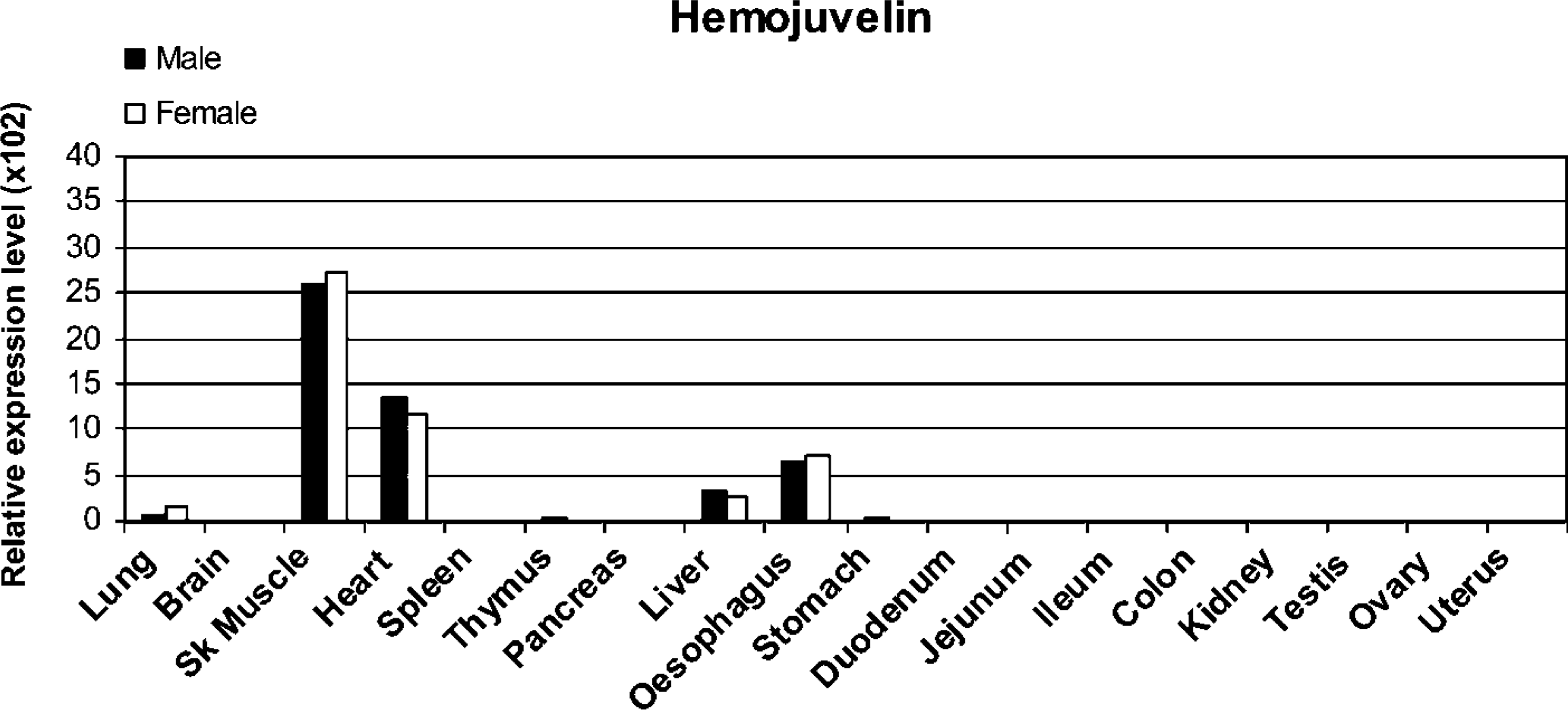
Mouse Hjv mRNA expression analyzed by real-time PCR. Four house-keeping genes (Bact, Gapdh, Hprt1, and Sdha) were used for the normalization of Hjv expression values.
After deparaffinization, immunostaining was performed by the biotin-streptavidin-peroxidase complex method. Briefly, antigen retrieval was performed in an autoclave at 95C. The parameters for neogenin were 30 min in 10 mmol/liter citrate buffer, pH 6.0. For hemojuvelin experiments, slides were treated for 25 min in 10 mmol/liter citrate buffer, pH 9. After endogenous peroxidase activity was quenched and nonspecific binding was blocked, the slides were incubated overnight at 4C with the rabbit anti-neogenin polyclonal antibody (1:50 dilution, sc-15337; Santa Cruz Biotechnology, Santa Cruz, CA) previously characterized (Lee et al. 2005) or alternatively with the 1:250 diluted rabbit anti-hemojuvelin polyclonal antibody that has also been previously characterized (Rodriguez Martinez et al. 2004). The slides were then washed three times with PBS for 10 min. Biotinylated goat anti-rabbit secondary antibody (Zymed Laboratories; South San Francisco, CA) was used at a dilution of 1:500 in a 60-min incubation step at room temperature. After washing, the sections were incubated with streptavidin-horseradish peroxidase conjugate (1:750 dilution; Zymed Laboratories) for 30 min at room temperature. The slides were then washed again four times for 5 min in PBS and the staining reaction was carried out using 3,3′-diaminobenzidine tetrahydrochloride (DAB) as a chromogen. Finally, a 4-min incubation in hematoxylin was performed to stain the nuclei of the cells and thus facilitate interpretation of the results. Neogenin experiments were accompanied by negative and positive control stainings to detect for possible nonspecific signals. Negative controls were processed by replacing the primary antibody with diluent. As a positive control parallel tissue sections were immunostained using a polyclonal rabbit anti-mouse PCNA antibody (Santa Cruz Biotechnology). Two kinds of negative control were used to detect possible nonspecific signals in the hemojuvelin staining. The preimmune serum controls consisted of replacement of the primary antibody with normal pre-immune rabbit serum. The diluent controls were performed as described above.
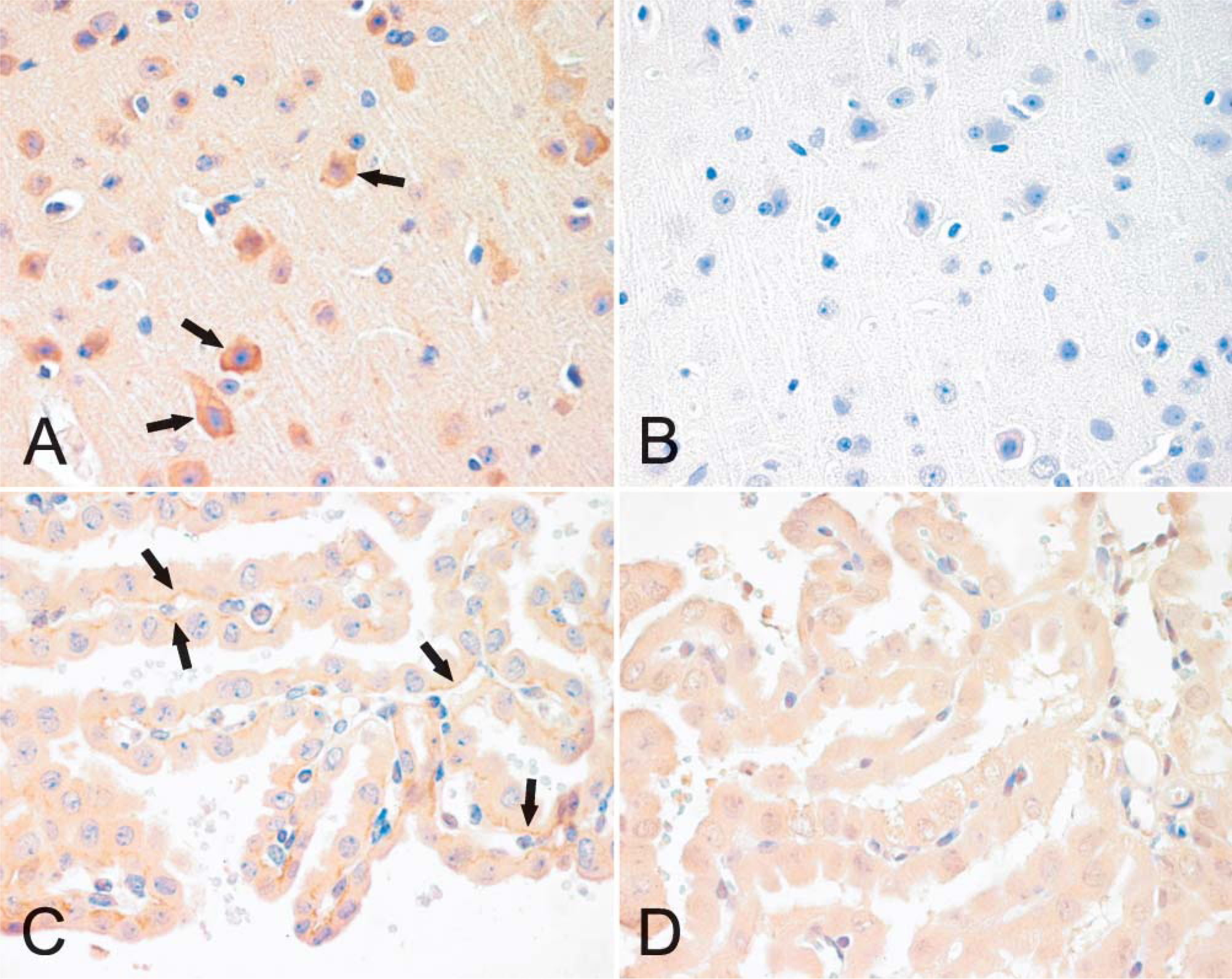
Immunohistochemical analysis of neogenin in mouse brain (
Results
Neo1 and Hjv Gene Expression Profiles in Male and Female Mice
Expression levels of Neo1 mRNA were examined in a broad range of mouse tissues by quantitative real time RT-PCR. As shown in Figure 1, Neo1 transcripts were detectable in each of the tissues tested for both genders with the highest expression observed in the testis, ovary, and uterus. A relatively high signal was also observed in brain, followed by lung, skeletal muscle, and heart. In the digestive system, the highest expression for Neo1 was found in distal parts of the intestine (the ileum and colon). However, moderate signal levels were also observed in the esophagus, stomach, duodenum, and jejunum, with a similar intensity as in the kidney, outside of the digestive system. The lowest Neo1 transcript levels were observed in the liver, thymus, spleen, and pancreas. There were no marked differences in the levels of expression observed in males vs females.
The expression of Hjv mRNA was also determined by real-time RT-PCR (Figure 2). Positive tissues included skeletal muscle, heart, esophagus, and liver in which the signal intensities were at approximately the same levels in both males and females. Very weak signals were also evident in the lung and in the thymus and stomach of male mice.
Expression of Neogenin and Hemojuvelin Proteins in Mouse
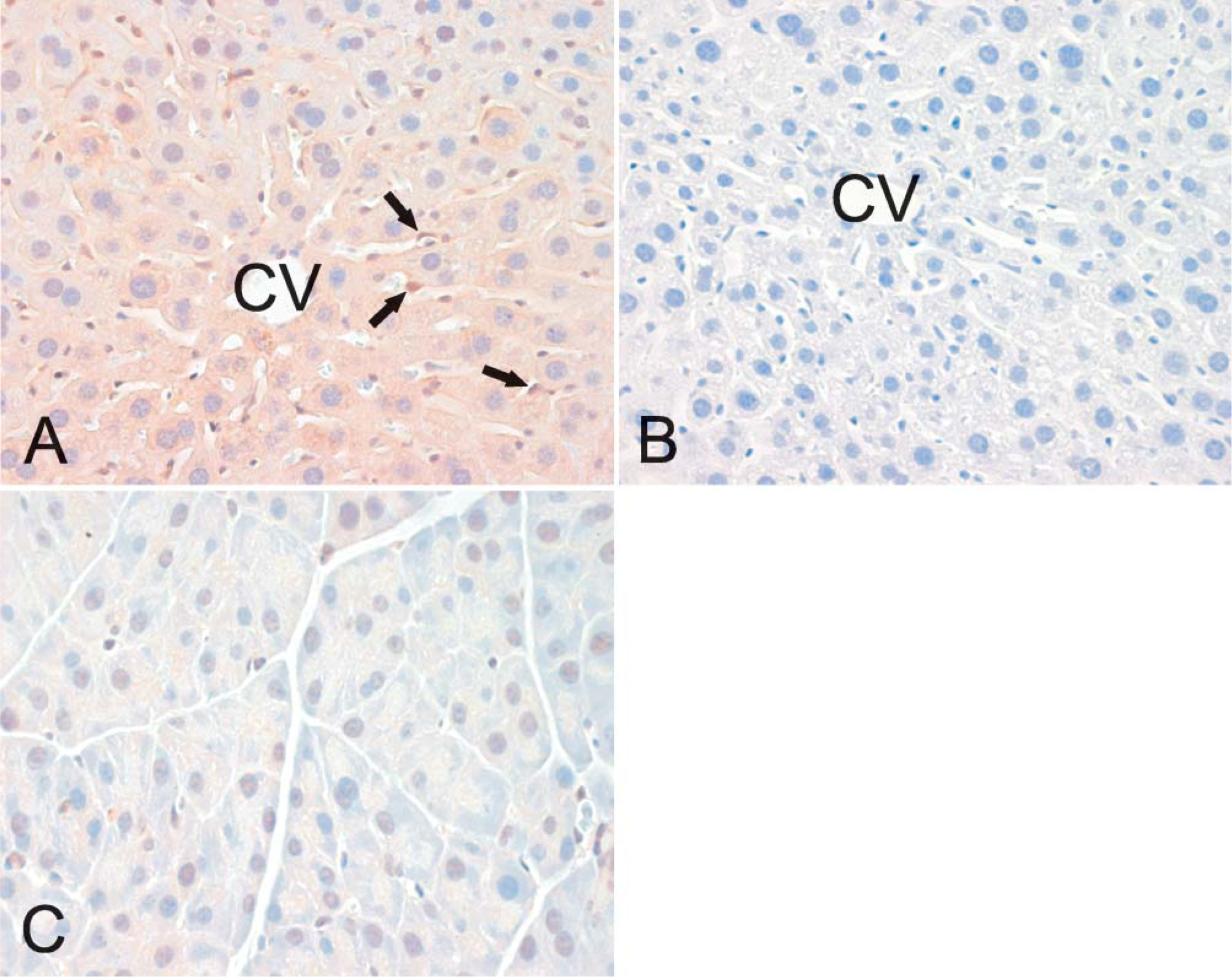
Localization of neogenin protein in mouse tissues was examined by immunohistochemistry and revealed no marked differences in the staining patterns between different strains or genders (data not shown). In the brain, positive staining was observed in neuronal bodies and weaker signals in the nervous fibers (Figure 3A). The diluent control in the brain remained mainly negative (Figure 3B). Neogenin was also detectable in the basolateral membrane of the choroid epithelial cells (Figure 3C). Figure 4 demonstrates the expression of neogenin in skeletal and heart muscles. In both cases the signal was slightly stronger in the sarcolemma than in the sarcoplasm of the myocytes (Figures 4A and 4C). The sinusoid-lining cells of the liver were also found to be positive for neogenin, whereas a weaker cytoplasmic staining was seen in the hepatocytes (Figure 5A). Furthermore, no expression was observed in the pancreas (Figure 5C).
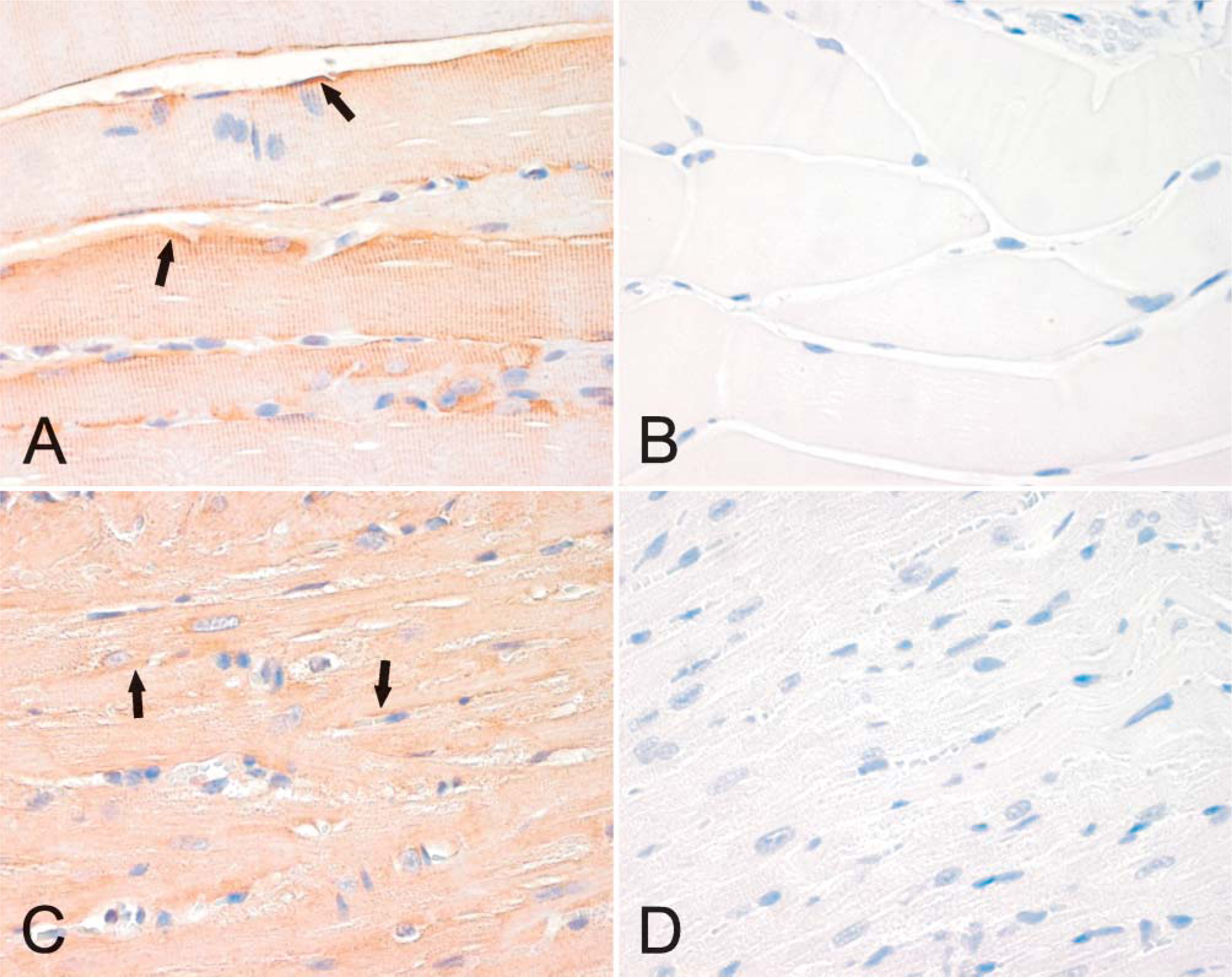
Immunolocalization of neogenin in mouse skeletal (
Figure 6 is representative of the expression profile of neogenin in different areas of the intestinal tract. Neogenin was observed in the mucus-secreting cells and chief cells of the gastric mucosa but was mainly absent from the parietal cells (Figure 6A). A weak staining pattern was also observed in the enterocytes of the duodenal mucosa, whereas the Brunner gland cells displayed a slightly stronger signal (data not shown). As shown in Figure 6C, the strongest reaction in the jejunum could be observed in the cytoplasm of the cryptal enterocytes, and the signal became slightly weaker toward the tips of the villi. In the ileum, neogenin showed cytoplasmic staining in the enterocytes of both the crypts and the villi (Figure 6D). The colon also showed a positive reaction inside the epithelial cells, as seen in Figure 6F. Neogenin expression was detected in the glomeruli and in some renal proximal and distal tubules (Figure 7A), whereas no staining was evident in the renal medulla (data not shown). The neogenin signals in the lung were weak and mainly located in round cells of the alveolar wall, most probably representing type II pneumocytes (Figure 7C). Neogenin localization was also found in the reproductive organs of both male and female mice (Figure 8). In the seminiferous tubules of the testis, positive cytoplasmic staining was observed in different stages of developing sperm cells (Figure 8A). Neogenin signal in the ovary was positive in the follicular cells (Figure 8C). As shown in Figure 8E, the cytoplasm of the epithelial cells of the endometrium also stained positively for neogenin and a similar signal was observed in the oviduct (data not shown).
Immunohistochemical analysis of neogenin in mouse liver (
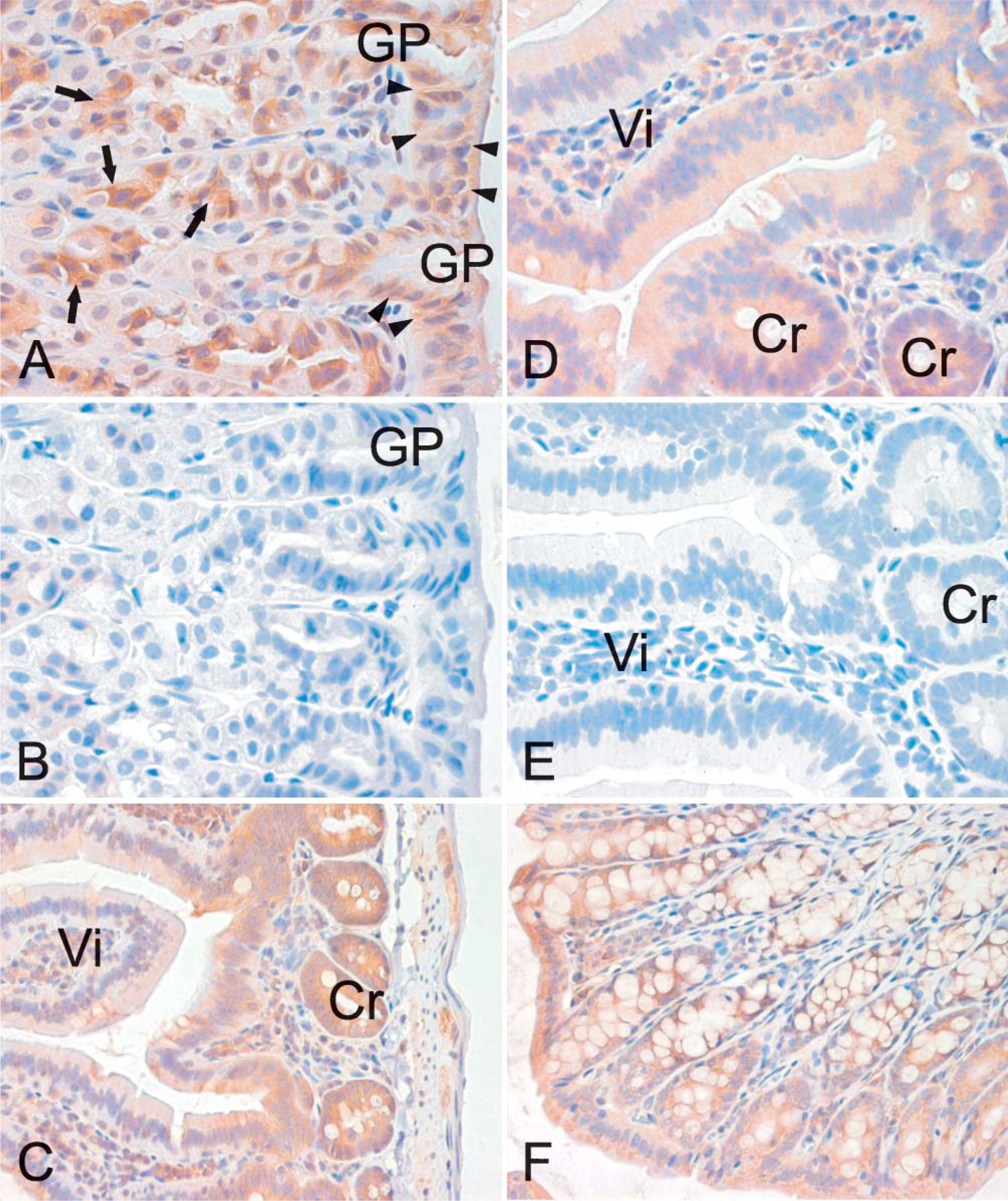
Localization of neogenin in different segments of the mouse intestine by immunohistochemistry. Neogenin protein shows a cytoplasmic location in the gastric mucosa (
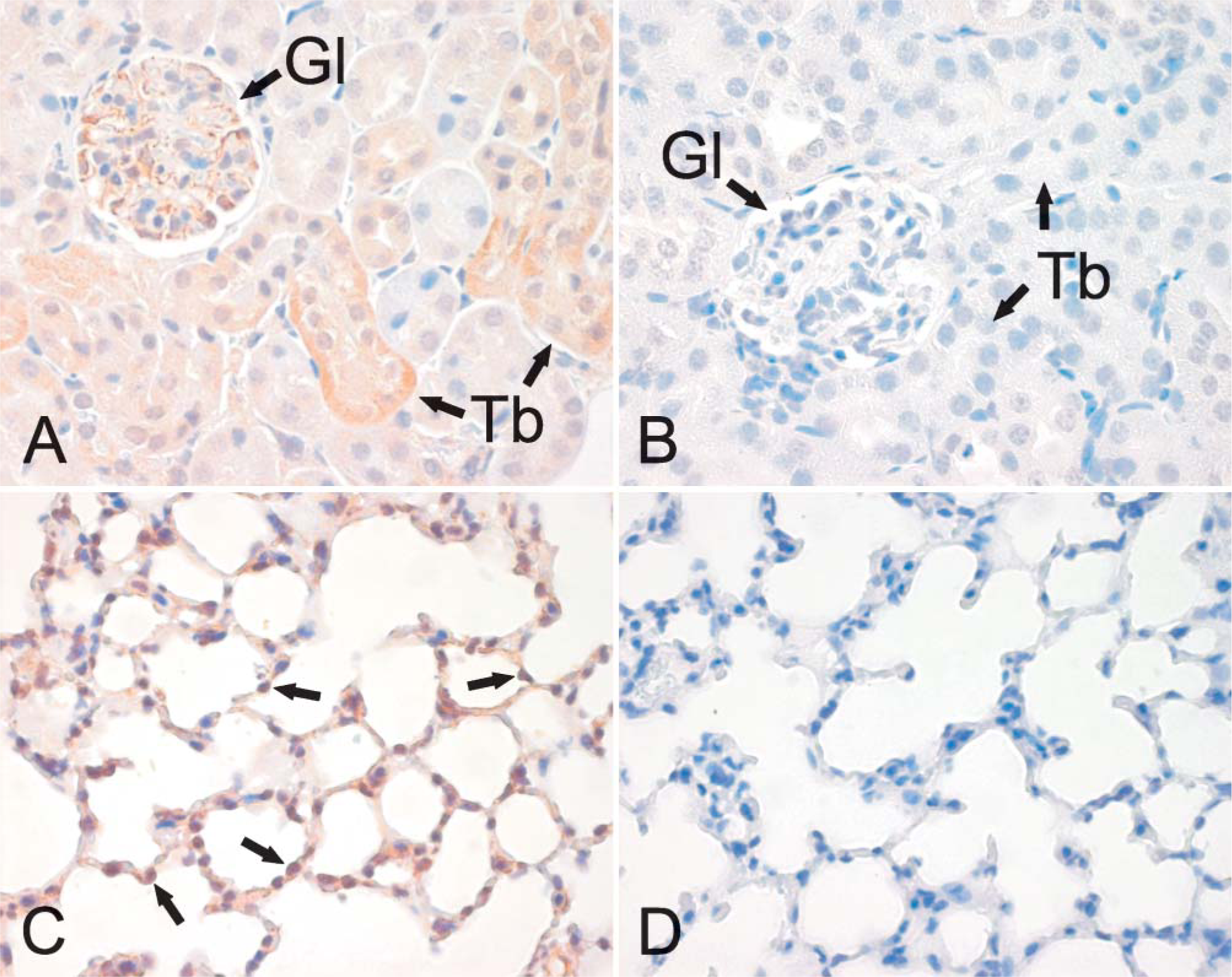
Immunohistochemical staining of neogenin in the mouse kidney and lung. The renal cortex shows a positive signal confined to the glomeruli and some segments of the renal proximal and distal tubules (
The localization of hemojuvelin protein in mouse skeletal muscle, heart, and liver is shown in Figure 9. There is a faint intracellular reaction in the skeletal muscle, whereas expression of hemojuvelin is negligible in the heart and liver.
Discussion
Most of the neogenin studies performed to date have focused on its roles during the development of the central nervous system. Hence, little is known about the functions of this protein in other tissues, in particular during adulthood. Currently, it seems clear that neogenin is a receptor for hemojuvelin and thus may be an important factor in the regulation of iron metabolism.
In our present study we further elucidate the specific sites of Hjv and Neo1 mRNA expression in mouse tissues using the quantitative and sensitive real-time RT-PCR method. Although four alternatively spliced exons have been described for the mouse Neo1 gene (Keeling et al. 1997), our primer design ensured that there would be no interference with any of these exons. Hence, our assay detects all transcripts that are present in individual tissues. It is also noteworthy that our use of four internal control genes for the normalization of gene expression levels enabled a higher degree of accuracy in our present study, compared with traditional normalizations based on only one housekeeping gene. This is of special importance when treating heterogeneous sets of samples, in which case the use of a single internal control may lead to erroneous conclusions.
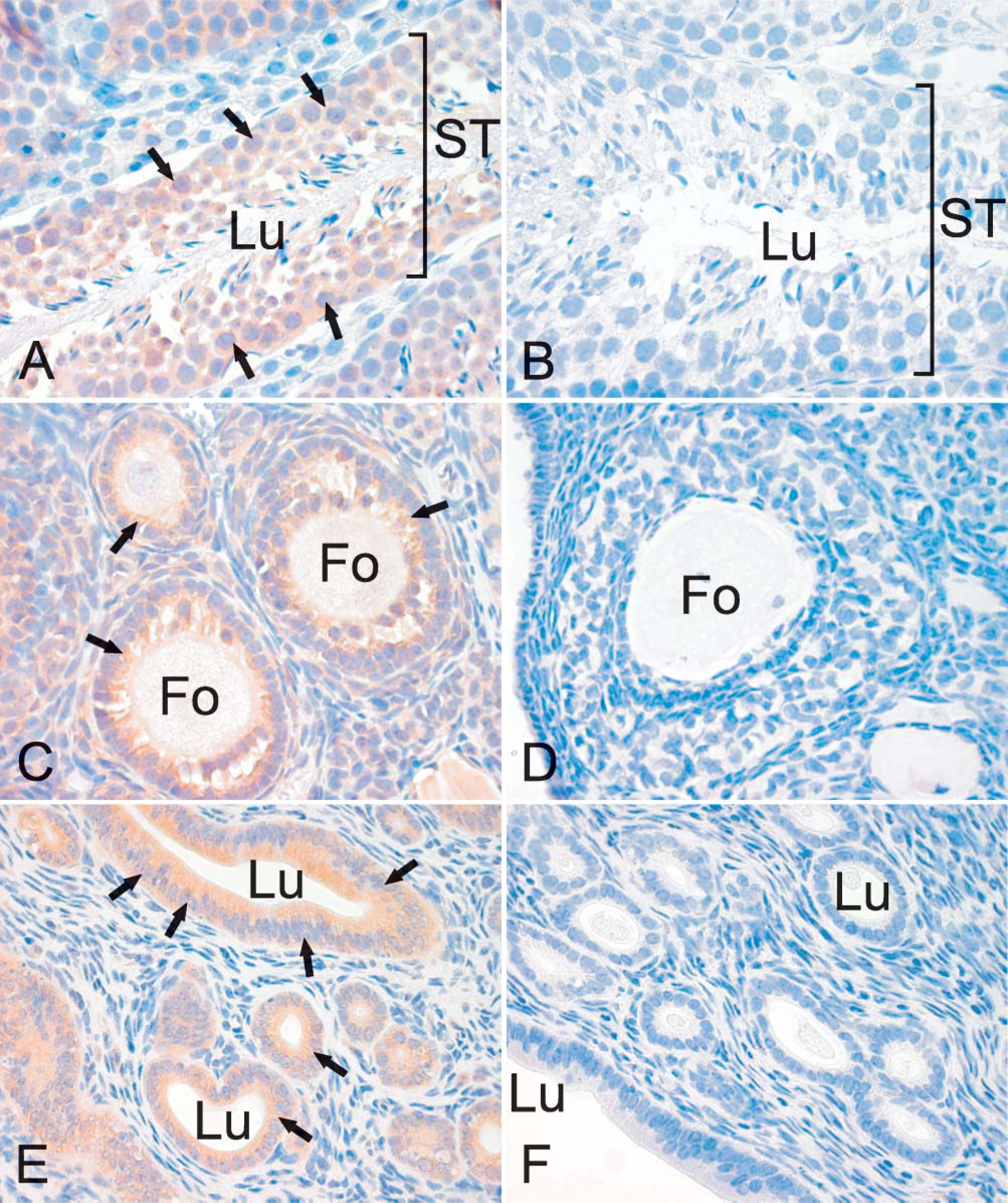
Immunolocalization of neogenin in the mouse reproductive system. In the longitudinal section of the seminiferous tubules of the testis (
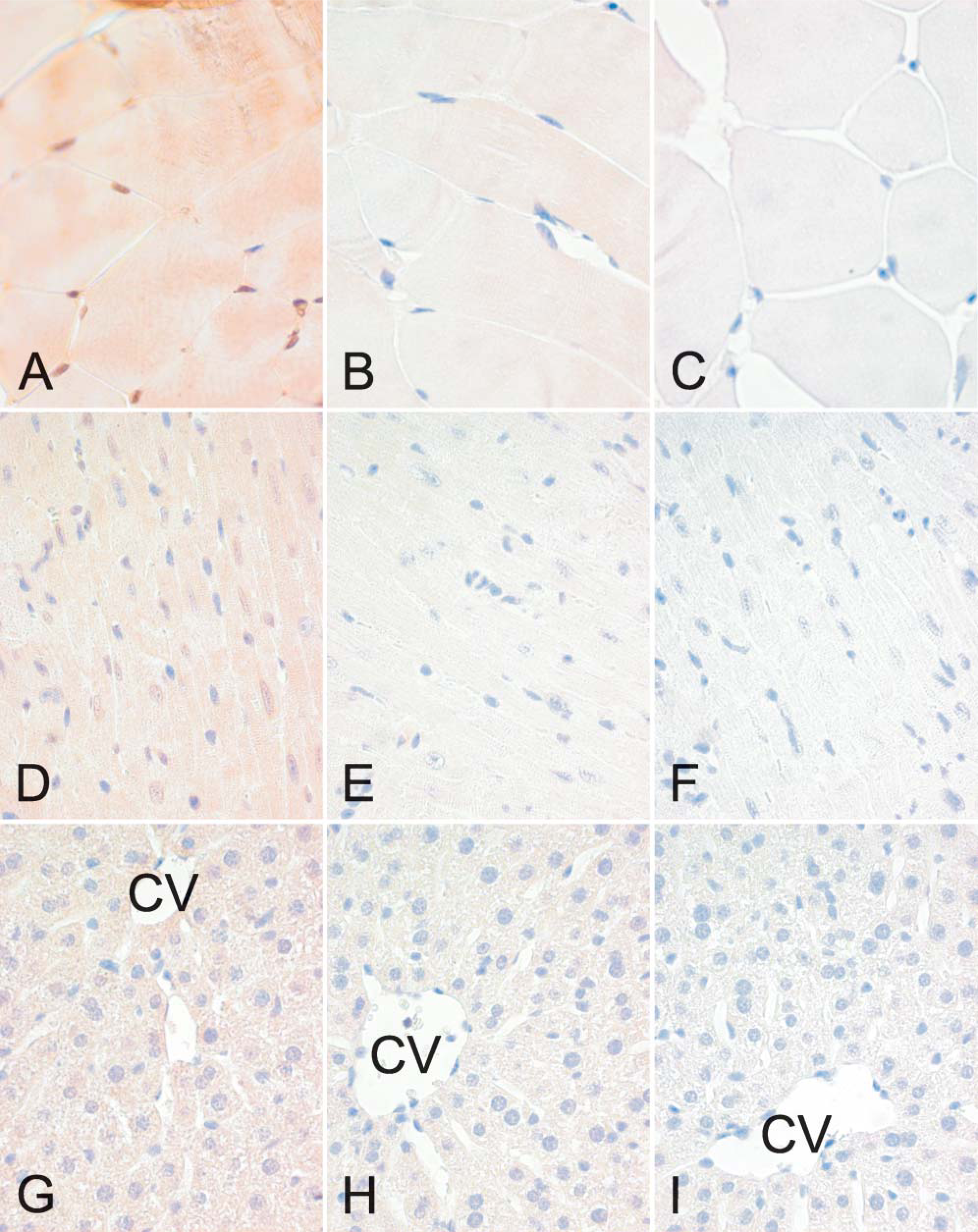
Localization of hemojuvelin in the mouse skeletal muscle (
Neo1 transcripts were detected in all tissue samples tested, with no notable variations observed between male and female mice. Our results are in agreement with a previous report in which Neo1 expression was detectable by Northern blotting in each of a smaller set of samples (Keeling et al. 1997). In addition, NEO1, the human ortholog of the mouse Neo1, was analyzed earlier by Northern blotting, with the strongest signal evident in skeletal muscle (Meyerhardt et al. 1997; Vielmetter et al. 1997). In our current experiments, the highest levels of Neo1 mRNA expression were detectable in the male and female reproductive systems (testis, ovary, and uterus). Accordingly, Neo1 had been shown previously to be present in the ovary by Northern blotting (Keeling et al. 1997). Northern analysis was also adopted in a study by Meyerhardt et al. (1997) that showed expression of NEO1 in testis and ovary. The role of neogenin and the identity of its ligand(s) in reproductive organs will certainly provide interesting novel targets for future investigations.
We also explored the localization of the neogenin protein in several mouse tissues using immunohistochemistry and found positive signals in the brain, skeletal muscle, heart, liver, stomach, duodenum, ileum, colon, kidney, lung, testis, ovary, oviduct, and uterus. No neogenin protein was detectable in the spleen, thymus, or pancreas, and the absence of any protein product is most likely due to a very low level of gene expression in these tissues, as seen by quantitative PCR and to differences in the sensitivities of the two methods.
Hjv mRNA expression profile was less extensive when compared with Neo1. As was observed for Neo1, differences between Hjv expression patterns in female and male tissues were small. Moreover, the expression analysis of murine Hjv mRNA reported herein is in agreement with previously reported Northern analyses of postnatal mice (Niederkofler et al. 2004). Previous in situ hybridization analyses in mouse embryos has also shown that Hjv is restricted to the skeletal and cardiac muscle (Oldekamp et al. 2004) and, more recently, it has been shown that mouse periportal hepatocytes express hemojuvelin protein (Niederkofler et al. 2005). Earlier reports of human HJV expression profiles obtained by Northern blot and conventional RT-PCR are also highly coincident with our present data (Papanikolaou et al. 2004; Rodriguez Martinez et al. 2004). In hemojuvelin immunohistochemistry, the highest staining reactions were detected in the skeletal muscle, which agrees well with the quantitative RT-PCR results. It is notable that expression levels of HJV mRNA were much lower as compared to neogenin. Thus, it is not surprising that hemojuvelin protein expression seemed to be very low in murine tissues.
Neogenin has turned out to be a multifaceted receptor with several ligands. The interaction between membrane-associated hemojuvelin and neogenin can only take place at sites where both molecules are expressed, and we have determined these overlapping regions to be the skeletal muscle, heart, liver, and esophagus. Interestingly, the liver is also the major site of hepcidin expression, which is considered to be a downstream target of hemojuvelin. Hepatocytes have been proposed to sense the iron status of the body and then either release or downregulate hepcidin. Thus, the interaction between hemojuvelin and neogenin might play a role in this sensing machinery. Although our immunohistochemical results indicated a very low expression of both neogenin and hemojuvelin in hepatocytes, the tempting possibility remains that iron levels or other factors could affect the expression and cellular distribution of these iron regulatory proteins in the liver. On the other hand, in spite of the narrow hemojuvelin expression profile that we identified in our current experiments, existence of a soluble form of hemojuvelin implies that it can probably function in several organs and cell types, depending on the presence of its receptor. The wide expression of neogenin also suggests that the hemojuvelin-neogenin pathway may, in fact, represent an important and extensive signaling cascade in the body.
Footnotes
Acknowledgements
This work was supported by grants from the Sigrid Juselius Foundation, Academy of Finland, and the Finnish Cancer Foundation.