
Abstract
There is evidence that estrogens can directly modulate human prostate cell activity. It has also been shown that cultured human prostate cancer LNCaPcan synthesize the active estrogen estradiol (E2). To elucidate the metabolism of estrogens in the human prostate, we have studied the expression of enzymes involved in the formation and inactivation of estrogens at the cellular level. 17β-Hydroxysteroid dehydrogenase (17β-HSD) types 1, 2, 4, 7, and 12, as well as aromatase mRNA and protein expressions, were studied in benign prostatic hyperplasia (BPH) specimens using in situ hybridization and immunohistochemistry. For 17β-HSD type 4, only in situ hybridization studies were performed. Identical results were obtained with in situ hybridization and immunohistochemistry. All the enzymes studied were shown to be expressed in both epithelial and stromal cells, with the exception of 17β-HSD types 4 and 7, which were detected only in the epithelial cells. On the basis of our previous results, showing that 3β-HSD and 17β-HSD type 5 are expressed in human prostate, and of the present data, it can be concluded that the human prostate expresses all the enzymes involved in the conversion of circulating dehydroepiandrosterone (DHEA) to E2. The local biosynthesis of E2 might be involved in the development and/or progression of prostate pathology such as BPH and prostate cancer through modulation of estrogen receptors, which are also expressed in epithelial and stromal cells.
T
In recent years, the existence of local biosynthesis of active sex steroids in several tissues has been supported by the demonstration of enzymes involved in the conversion of the inactive adrenal precursor dehydroepiandrosterone (DHEA) and DHEA sulfate to E2 and dihydrotestosterone (DHT) (Martel et al. 1994; Labrie et al. 2000). Human prostate has been shown to express all the steroidogenic enzymes involved in the conversion of DHEA to the active androgen DHT (Pelletier et al. 1998; El-Alfy et al. 1999). DHEA is transformed into androstenedione by 3β-hydroxysteroid dehydrogenase (3β-HSD) and then into testosterone by 17β-HSD type 5 in basal cells of the glandular epithelial and stromal cells, whereas DHT is synthesized in the luminal cells and stromal cells by the action of 5α-reductase.
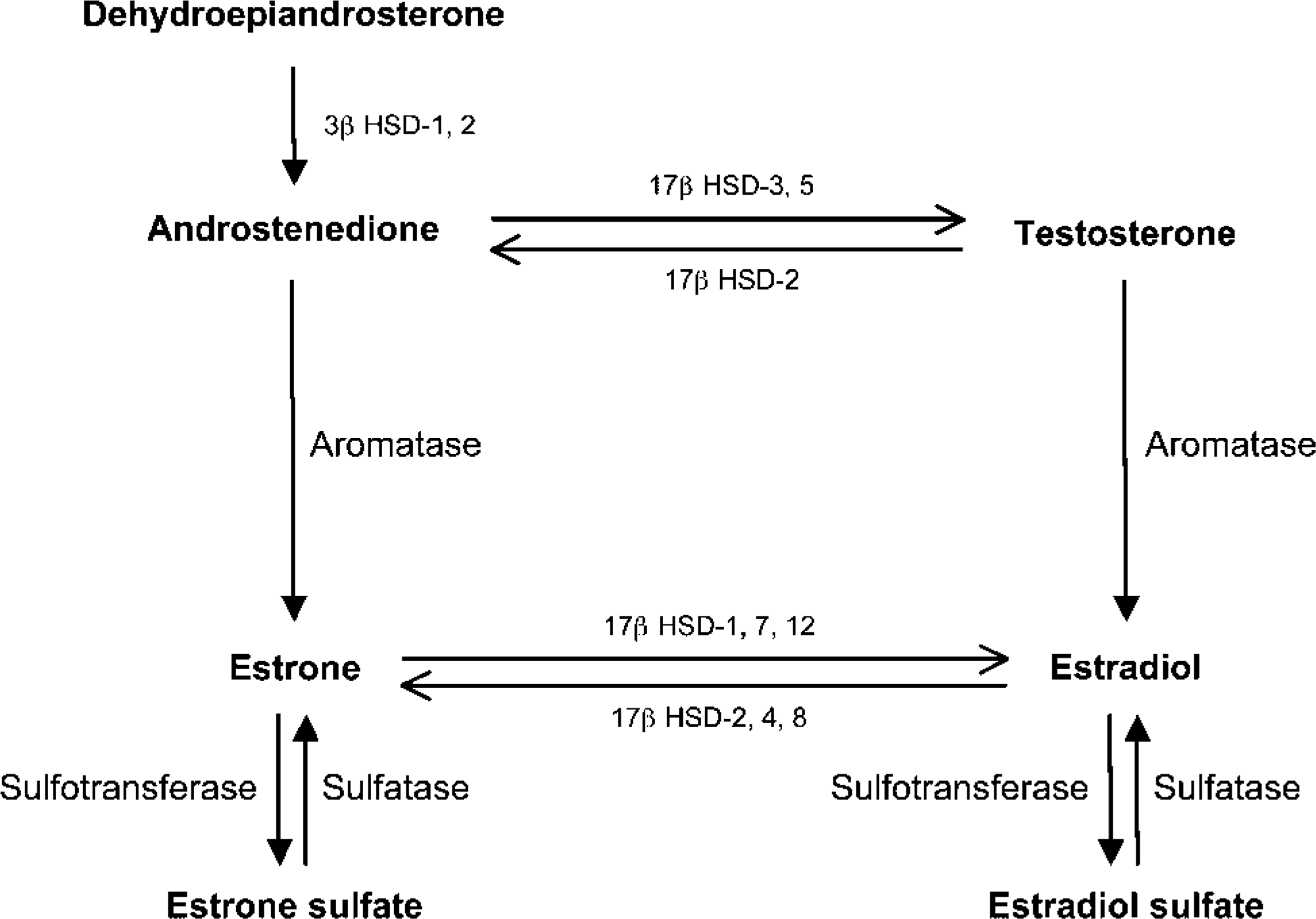
Schematic representation of pathways involved in biosynthesis and metabolism of estradiol from circulating dehydroepiandrosterone.
There is some evidence that E2 can be locally produced in the prostate. Aromatase, which converts testosterone into E2 and androstenedione into the weak estrogen estrone (E1), has been found to be expressed in human prostate (Figure 1) (Matzkin and Soloway 1992; Tsugaya et al. 1996; Ellem et al. 2004). On the other hand, there are reports indicating that aromatase is not expressed in human prostate and prostate cancer (Smith et al. 1982; Brodie et al. 1989; Negri-Cesi et al. 1998). Reductive 17β-HSDs, such as types 1, 7, and 12 17β-HSD are involved in the conversion of E1 to E2, whereas types 2,4,8,10, and 11 are oxidative enzymes that convert E2 to E1 (Figure 1) (Luu-The 2001; Mindnich et al. 2004; Vihko et al. 2004). We (Pelletier et al. 2004) have recently reported that in the mouse prostate, 17β-HSD types 1, 2, 4, 7, 8, 10, and 11 mRNAs are expressed in the epithelial cells of acini. In human, 17β-HSD type 2 has been found to be expressed in benign and malignant prostate tissue (Vihko et al. 2004).
To identify the enzymes that might be responsible for E2 production or inactivation, we have studied by in situ hybridization and immunohistochemistry the localization of aromatase and 17β-HSD types 1, 2, 4, 7, and 12 in human hyperplastic prostatic tissue.
Materials and Methods
Tissue Preparation
Adult prostatic tissue was obtained from eight patients with symptomatic benign prostatic hyperplasia (BPH) undergoing transurethral prostatectomy (age of the patients, 60 to 80 years). The specimens were immediately fixed by immersion in 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). After 24-hr fixation, the specimens were embedded in paraffin or frozen at −70C.
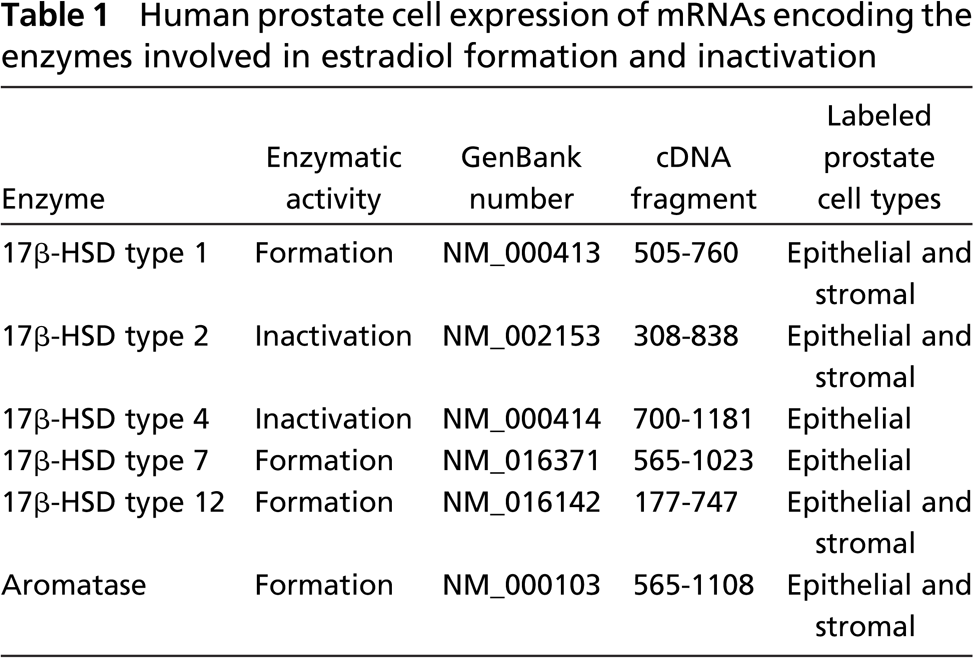
Human prostate cell expression of mRNAs encoding the enzymes involved in estradiol formation and inactivation
In Situ Hybridization
In preliminary experiments, no demonstrative labeling could be obtained when in situ hybridization was performed on paraffin sections. We then used frozen sections (10-μm) mounted onto glass slides. The vector used for production of the cRNA probe was constructed by insertion into a pCR-Blunt II-TOPO (Invitrogen; Ontario, Canada) of cDNA fragments. The characteristics of the fragments of the human enzyme cDNAs that have been studied are listed in Table 1. In situ hybridization with the antisense and sense 35S-labeled cRNA probes was performed as previously described (Givalois et al. 1997). Briefly, the sections were prehybridized at room temperature in a humid chamber for 2 hr in 450 μl/slide of a pre-hybridization buffer containing 50% formamide, 5 × SSPE (1 × SSPE = 0.1 M NaCl, 10 mM NaH2PO4, pH 7.4, 0.8 mM EDTA), 5X Denhart's buffer, yeast-t-RNA 200 μg/ml, denatured salmon testis DNA (Sigma; St Louis, MO), 200 μg/ml Poly A (Boehringer-Mannheim; Montreal, Canada), and 4% dextran sulfate. After prehybridization treatment, 100 μl hybridization mixture (prehybridization buffer containing 10 mM dithiothreitol and the 35S-labeled cRNA probe at a concentration of 10 × 106 cpm/ml) was spotted on each slide, sealed under a coverslip, and incubated at 60C overnight (15-20 hr) in a humid chamber.
After hybridization, coverslips were removed and slides were rinsed in 2 × SSC at room temperature for 30 min. Sections were then digested by RNase A (20 μg/ml in 2 × SSC) at 37C for 30 min, rinsed in decreasing concentrations of SSC (2 × SSC and 1 × SSC) for 30 min at room temperature, washed in 0.5 × SSC for 30 min at 37C, followed by 90 min at room temperature in 0.5 × SSC, and finally for 30 min at room temperature in 0.1 × SSC.
(
(
After hybridization, the sections were exposed to Hyperfilm (Kodak; Rochester, NY) for 4-5 days and subsequently coated with liquid photographic emulsion (Kodak-NTB2; diluted 1:1 with water). Slides were exposed for 14-45 days, developed in Dektol developer (Kodak) for 2 min, and fixed in rapid fixer (Kodak) for 4 min. Thereafter, the sections were rinsed and stained with hematoxylin and eosin.
Immunohistochemistry
The immunohistochemical localization of 17β-HSD types 1, 2, 7, and 12 and aromatase was performed as previously described (Pelletier et al. 1999). Deparaffinized sections were incubated at 4C with the antisera to 17β-HSD types 1, 2, 7, and 12 and aromatase diluted 1:500. The antiserum against human 17β-HSD type 1 has been described previously (Luu-The et al. 1989; Dupont et al. 1991). For preparation of antibodies to human 17β-HSD type 2, we overproduced the enzyme in Escherichia coli in fusion with glutathione-S-transferase, using pGEX-1λT expression vector (Amersham Biosciences; Baie d'Urfe, Quebec, Canada). To develop antibodies to 17β-HSD types 7 and 12, we selected the following peptide sequences: P261 to L342 for the type 7 and H217 to N312 for the type 12. These peptide sequences were overproduced in E. Coli BL-1 using PET23a expression vector (EMD Biosciences; San Diego, CA). The purified proteins were subsequently diluted in PBS containing 50% complete Freund's adjuvant (concentration: 3 mg/ml) and injected subcutaneously with 1 ml of the solution at multiple sites on New Zealand rabbits. The animals were treated twice with the same amounts of proteins in 50% incomplete Freund's adjuvant at 1-month intervals. Antisera were analyzed by immunoblot using HEK-293 cells non-transfected and stably transfected with 17β-HSD types 2, 7, or 12, as negative and positive controls, respectively. The experiment was conducted in an animal facility approved by the Canadian Council on Animal Care (CCAC; Ottawa, Ontario, Canada) and the Association for the Assessment and Accreditation of Laboratory Animal Care (Rockville, MD). The study was performed in accordance with the CCAC guide for the care and use of experimental animals. The rabbit antiserum to human P450 aromatase was kindly provided by Dr. G.E. Seralini (Caen, France) (Almadhidi et al. 1995).
Control experiments were performed on adjacent sections by substituting preimmune rabbit serum (1:500) or the antiserum preabsorbed with an excess of the corresponding antigen (10-6 M). All the primary antibodies were detected following incubation at room temperature for 4 hr with peroxidaselabeled goat anti-rabbit γ-globulin (HyClone Laboratories, Inc.; Logan, UT) diluted at 1:200 (Pelletier et al. 1999). The peroxidase was revealed during incubation with 10 mg of 3.3î-diaminolenzidine in 100 ml of Tris-saline buffer containing 0.03% H2O2. The intensity of staining was controlled under the microscope.
Results
In Situ Hybridization
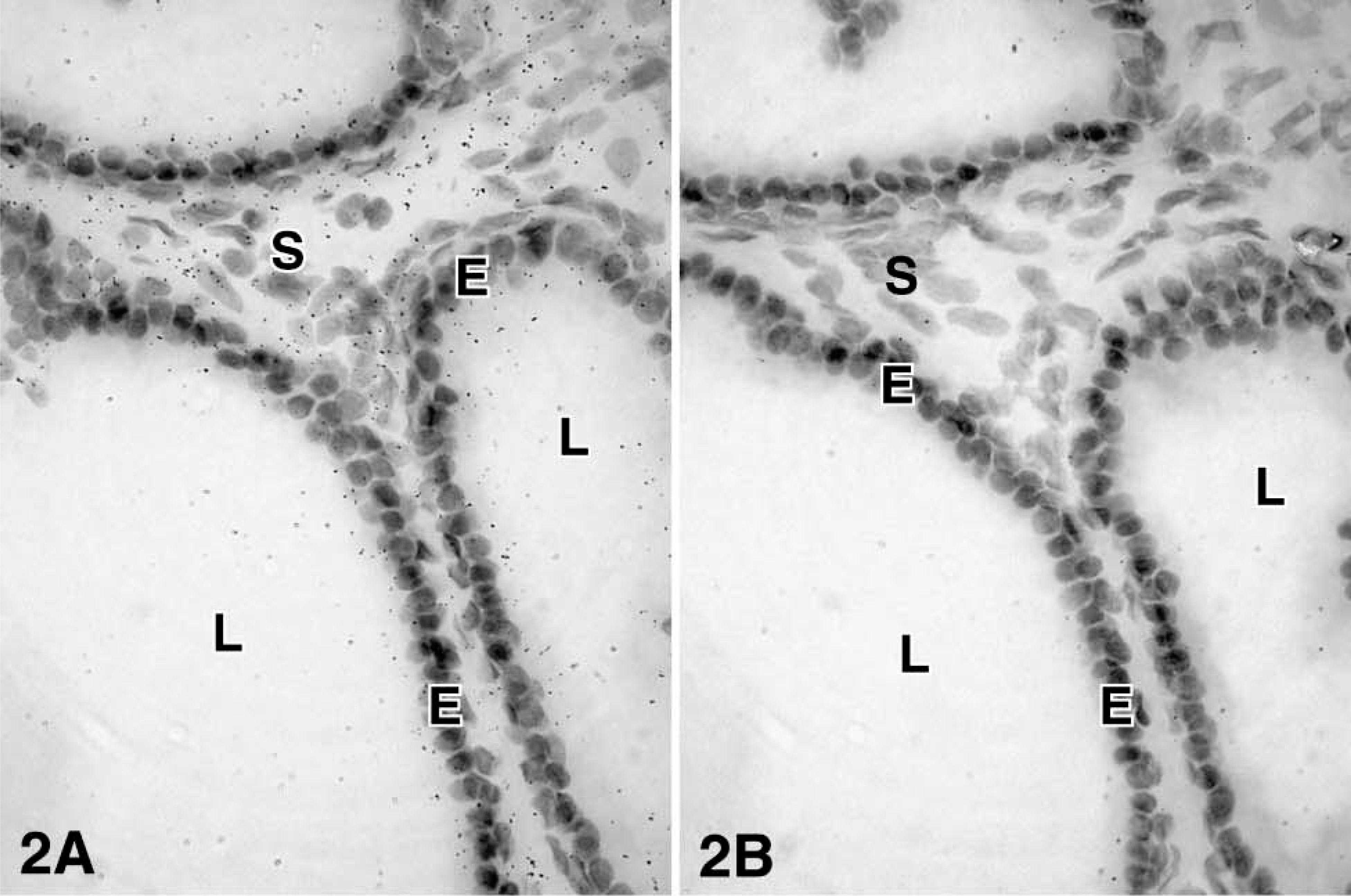
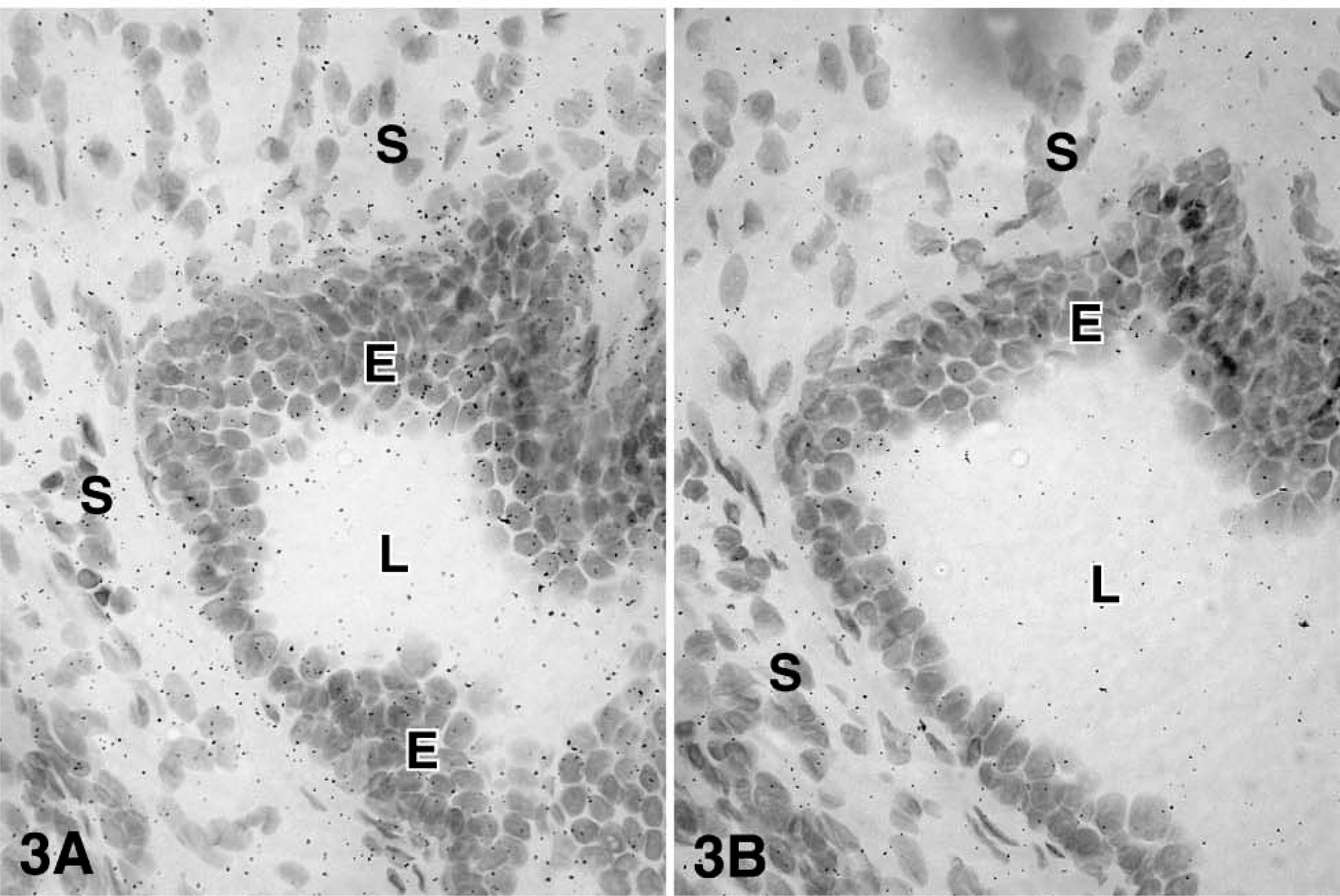
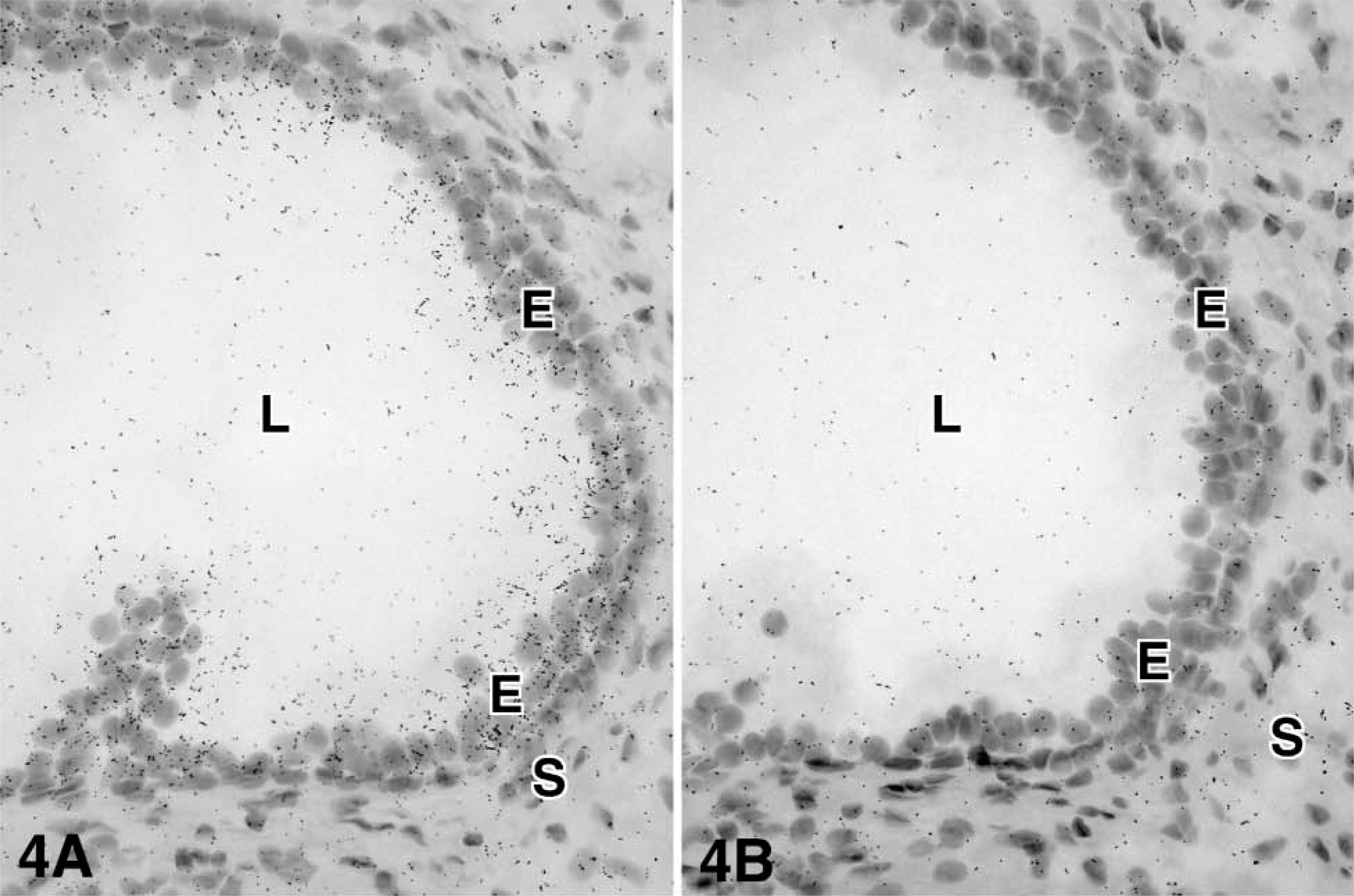
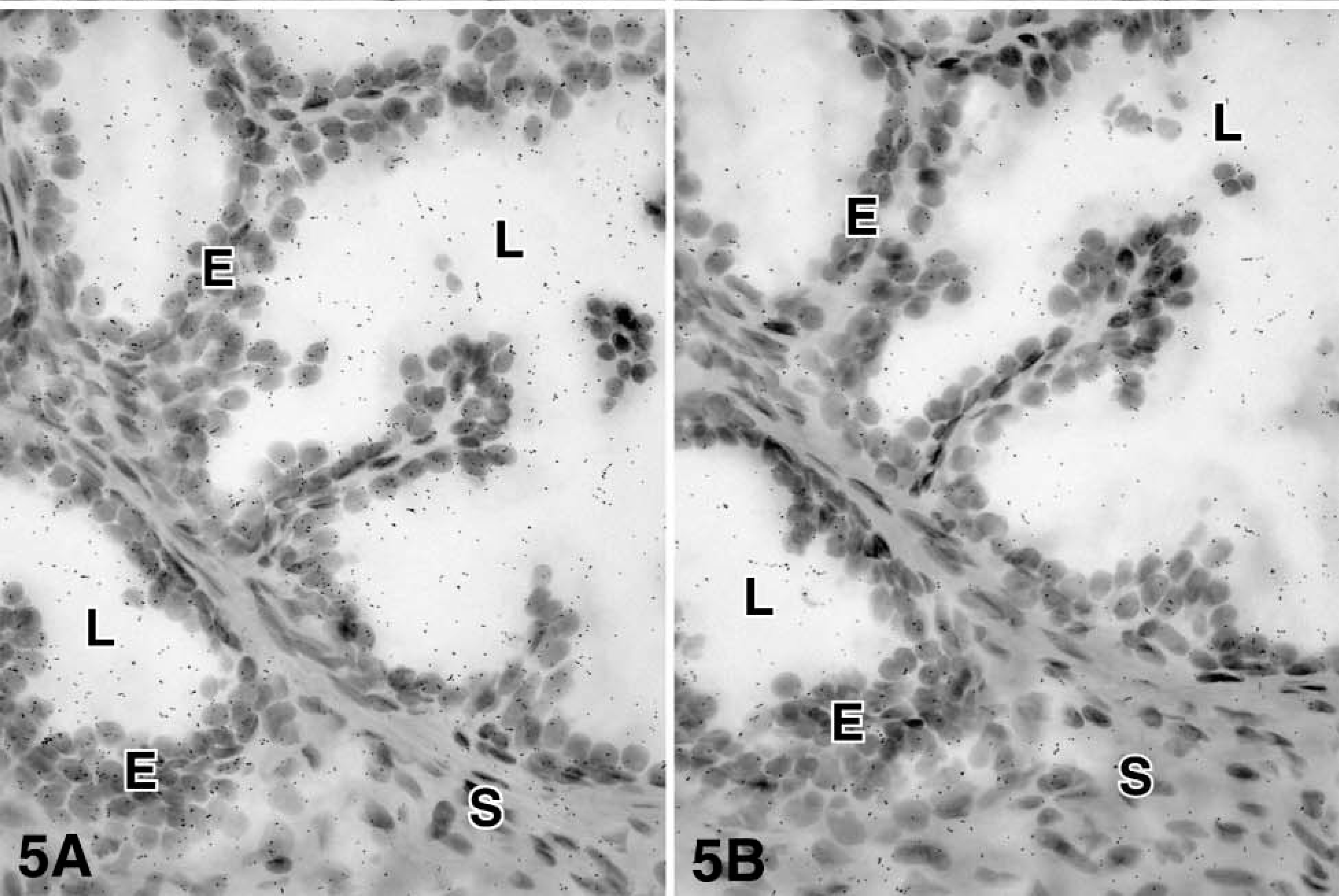
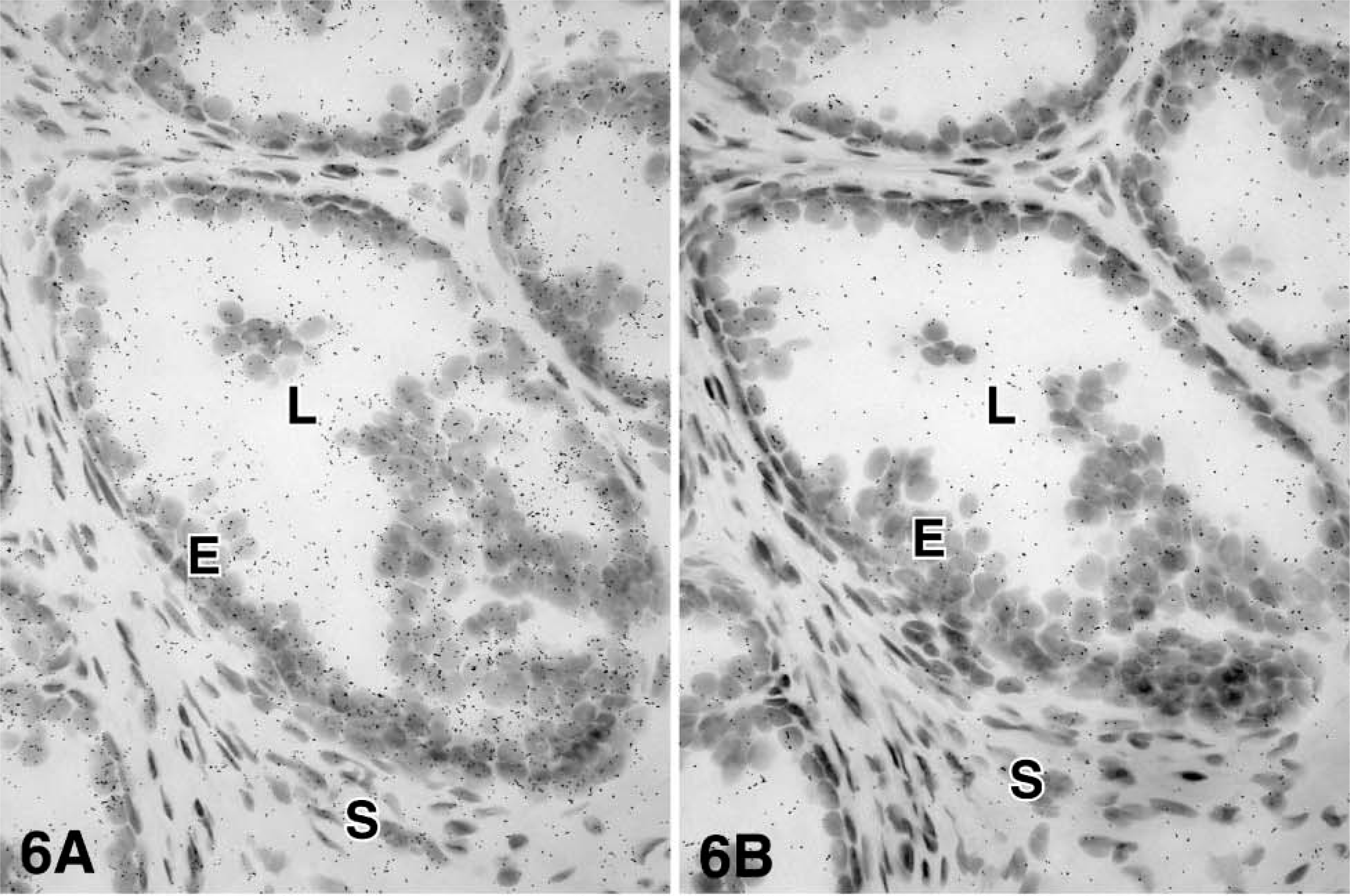
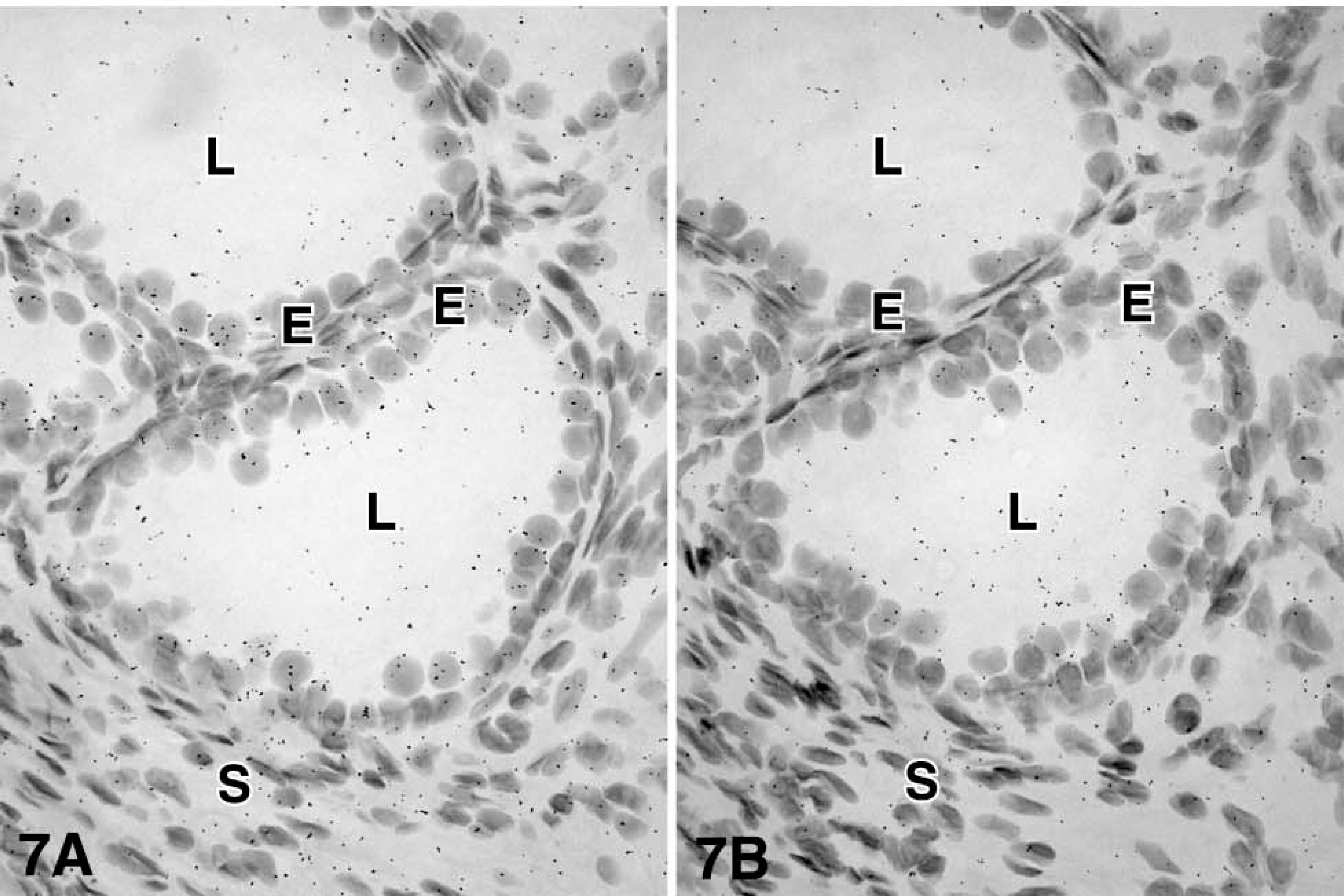
For each enzyme, at least three different specimens were studied, and the results were consistent. After 4-5 days of exposure to the films, specific hybridization signal could be obtained for all the cRNA probes. At the light microscopic level, specific labeling was detected in both epithelial cells of the prostatic alveoli and stromal cells for the following enzymes: 17β-HSD types 1, 2, and 12 and aromatase. On the other hand, 17β-HSD types 4 and 7 mRNA have been found to be expressed only in epithelial cells (Table 1 and Figures 2-7). Because frozen sections do not allow clear identification of the basal cells of the alveoli, it cannot be excluded that labeling occurs not only in luminal cells, which are clearly labeled, but also in the basal cells. Also, because the cytoplasm of stromal cells could not be stained by hematoxylin and eosin, it was not possible to define the outlines of those cells. No specific signal could be detected in blood vessel walls.
Immunohistochemistry
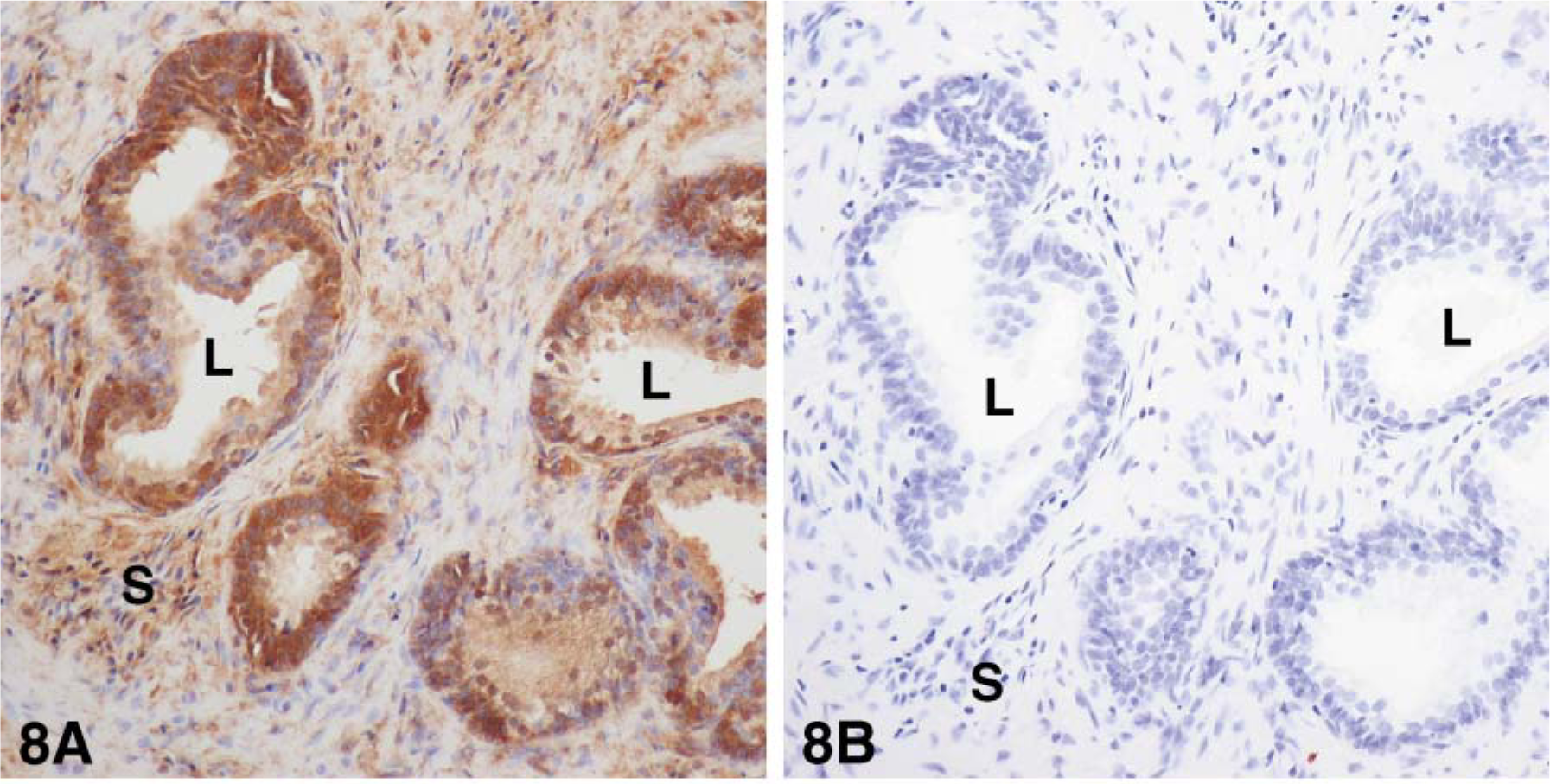
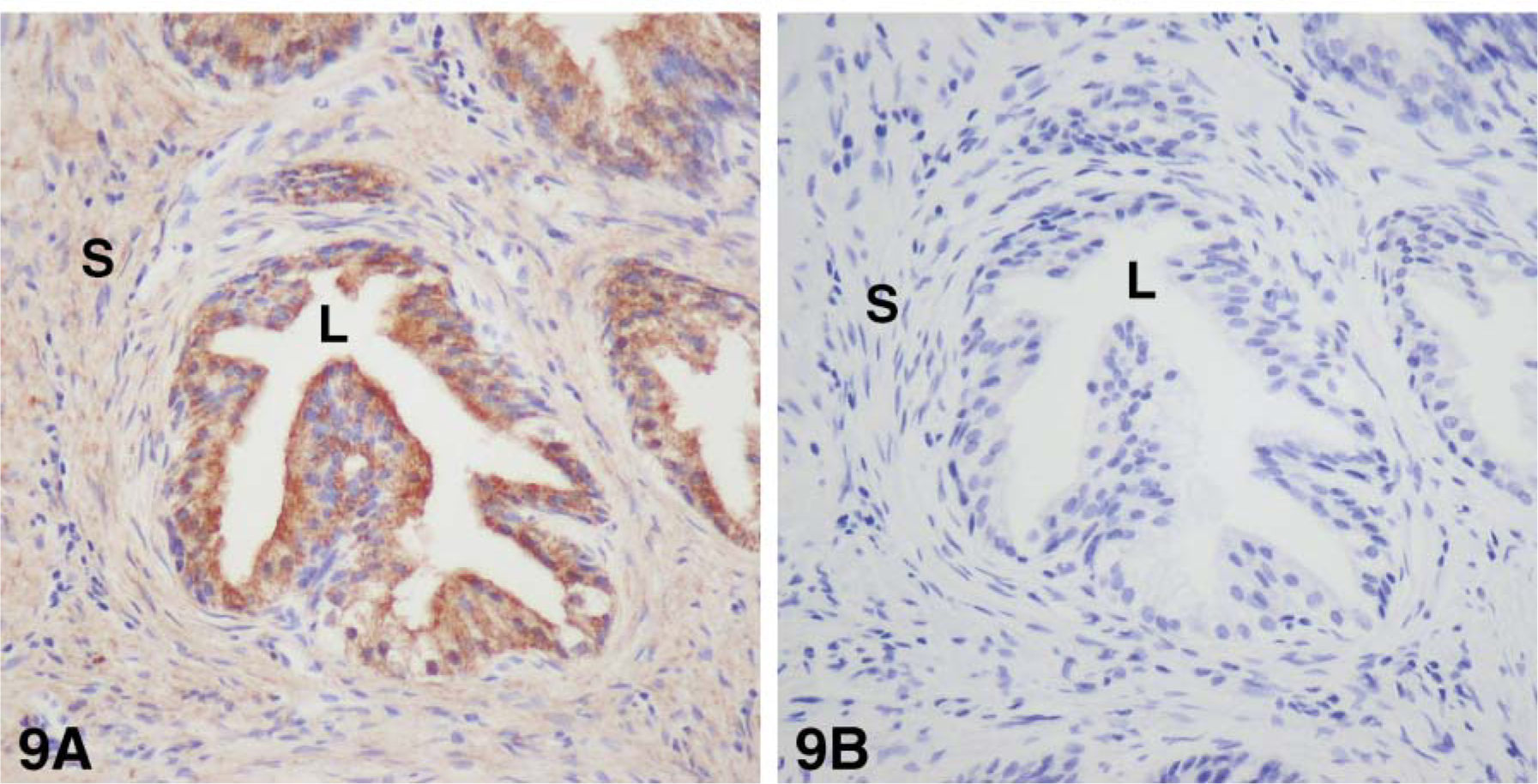
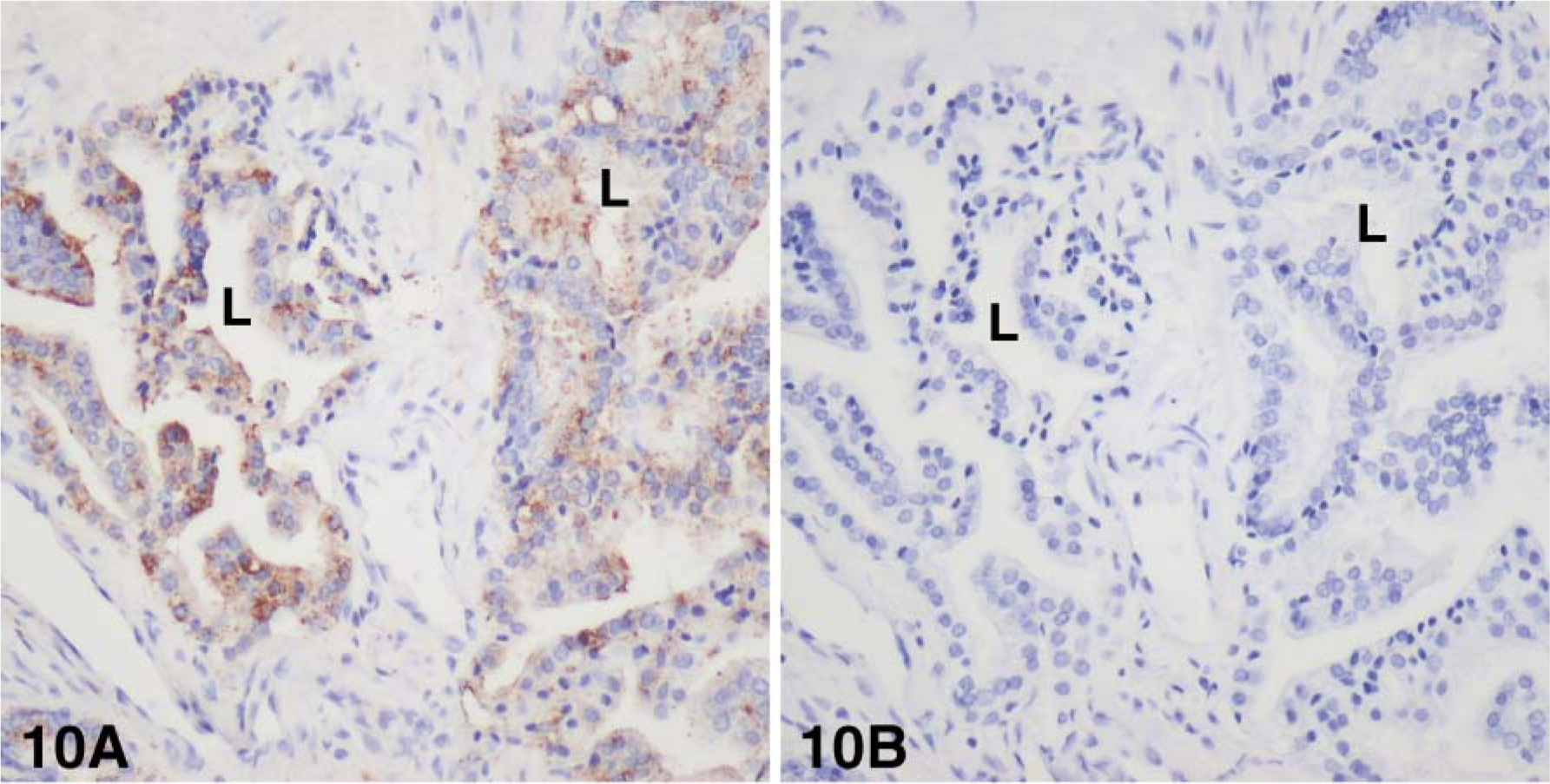
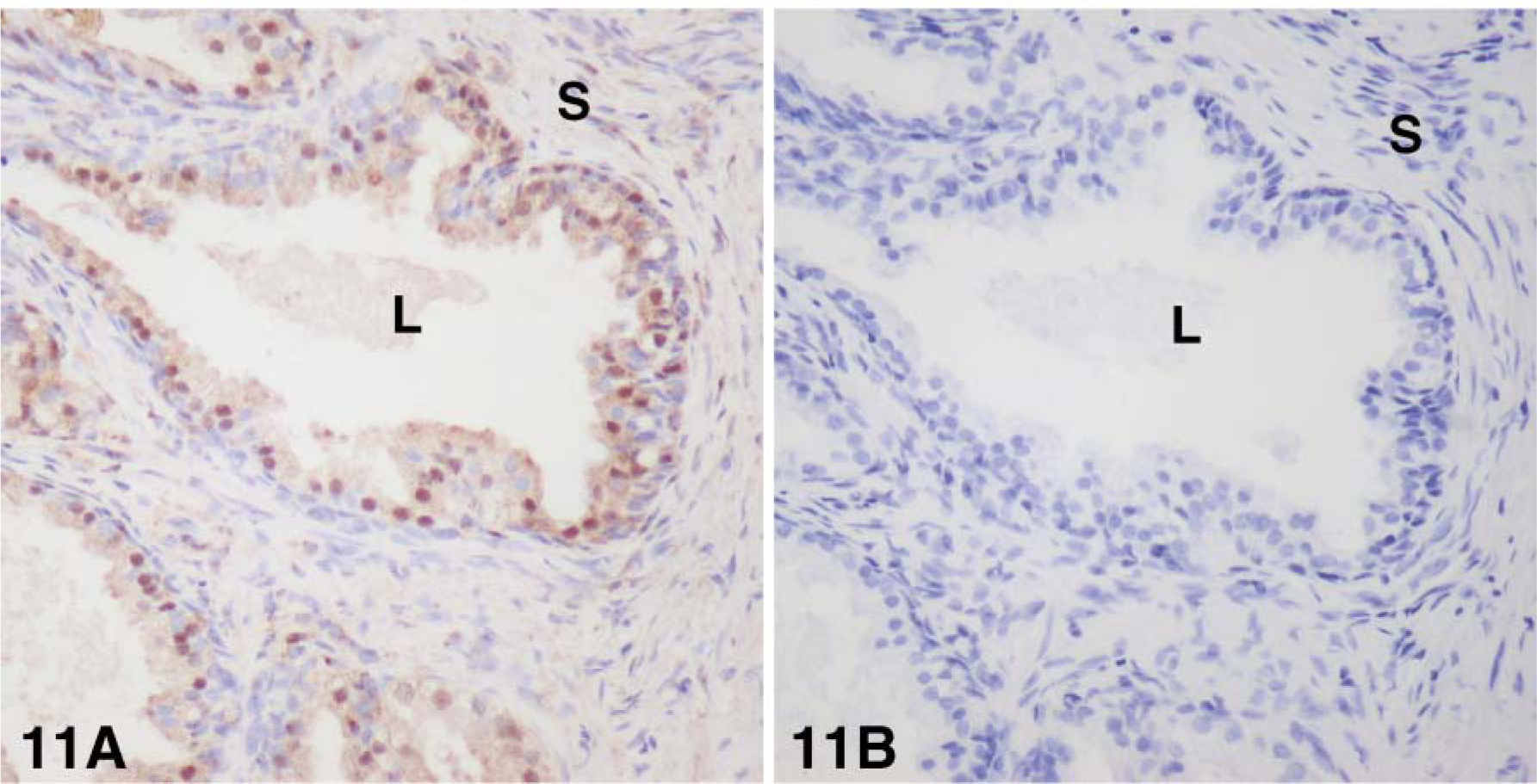
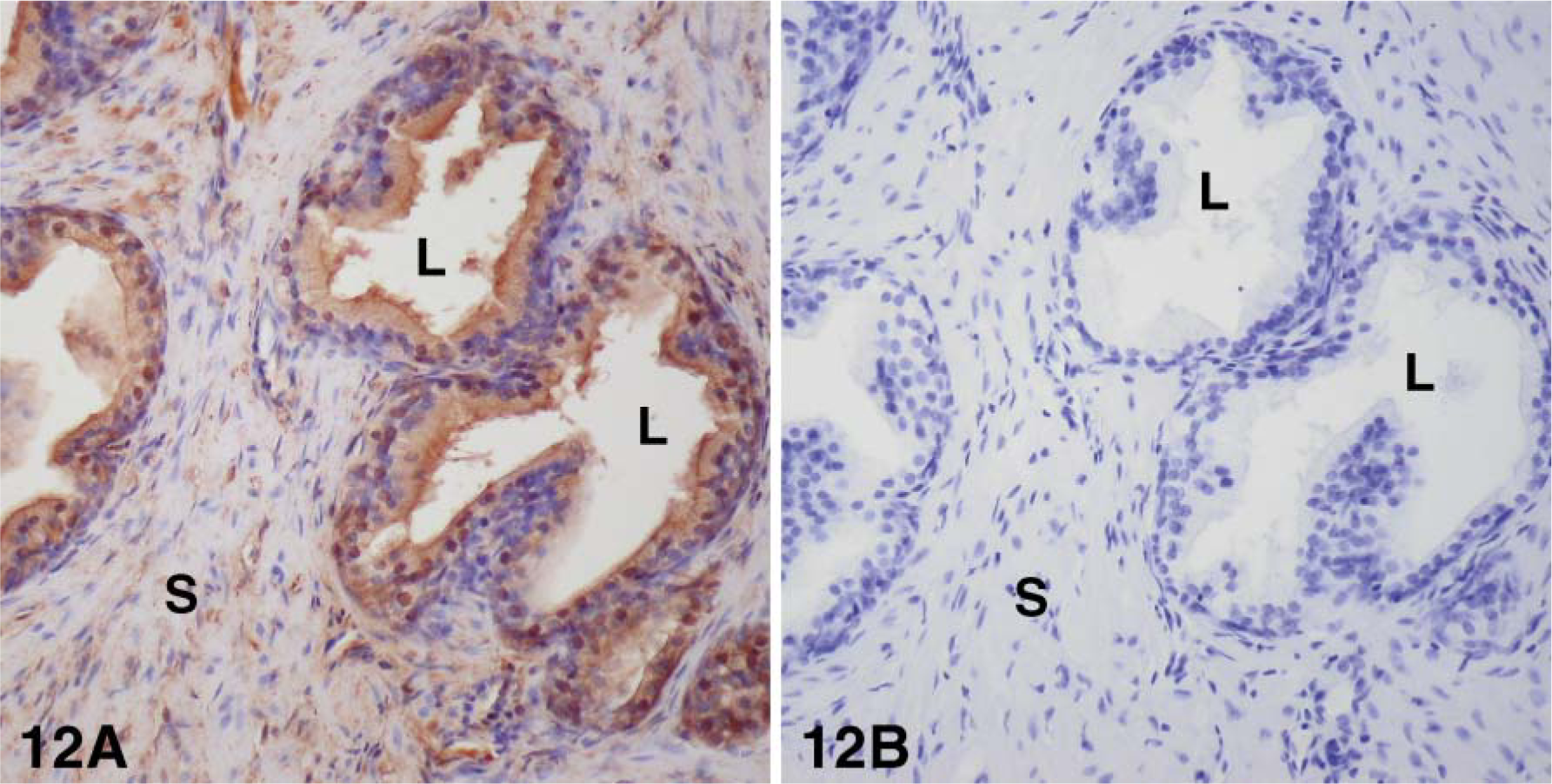
Using specific antibodies to the enzymes 17β-HSD types 1, 2, 7, and 12 and aromatase, we have been able to localize each enzyme by immunohistochemistry. Type 1 17β-HSD immunoreactivity was observed in the luminal cells of the epithelium, as well as in several stromal cells. The staining was located in the cytoplasm, in some nuclei in epithelial cells, and in the cytoplasm in stromal cells (Figure 8). With antibodies to 17β-HSD type 2, strong staining was detected in the cytoplasm, especially at the apex, and in the luminal cells of the alveoli, and light staining could be associated with fibers in the stromal compartment (Figure 9). Immunostaining for 17β-HSD type 7 was restricted to the cytoplasm of luminal cells of alveoli; the stroma were devoid of any immunoreactivity (Figure 10). Immunoreactive 17β-HSD type 12 was found in luminal cells of the alveoli, where the staining was both cytoplasmic and nuclear (Figure 11). In the stroma, several cells also exhibited nuclear and cytoplasmic labeling. Fibers were lightly stained. Aromatase immunoreactivity was found in the cytoplasm of luminal cells as well as several stromal cells (Figure 12). As for 17β-HSD type 2, a strong staining could be observed at the apex of luminal cells.
(
(
When we used antisera immunoabsorbed with the corresponding antigen or preimmune antisera, no reaction could be detected (Figures 8-12). Because no antibodies to 17β-HSD type 4 are available, the expression of the enzyme protein could not be evaluated in the present study.
Discussion
Combining two complementary approaches, namely in situ hybridization and immunohistochemistry, we have studied 17β-HSD types 1, 2, 4, 7, and 12 and aromatase expression in human prostate. For all the enzymes studied, except for 17β-HSD type 4, for which no immunostaining was performed, the results were identical with both approaches.
We previously reported that 3β-HSD, which converts DHEA into androstenedione, and 17β-HSD type 5, which transforms androstenedione into testosterone, are expressed in epithelial cells (predominantly basal cells) as well as stromal cells in BPH (El-Alfy et al. 1999; Pelletier et al. 1999). In the present study, we report for the first time the cellular localization of 17β-HSD types 1, 2, 4, 7, and 12. All of these enzymes are expressed in luminal cells bordering the alveolar lumen. Moreover, 17β-HSD types 1, 2, and 12 were also found to be expressed in stromal cells. These findings are in agreement with our previous findings on mouse prostate showing that 17β-HSD types 4 and 7 are expressed in epithelial cells, whereas types 1 and 2 are expressed in both epithelial and stromal cells (Pelletier et al. 2004). In this previous study, the recently identified type 12 (Luu-The et al. 2006) was not investigated. 17β-HSD types 2, 4, and 7 have already been reported to be expressed in the human prostate, as determined by RT-PCR analysis (Krazeisen et al. 1999; Peltoketo et al. 1999; Luu-The 2001; Vihko et al. 2004).
In the present study, we have observed expression of aromatase mRNA and protein in both epithelial and stromal cells in human prostate. By immunohistochemistry, it has been previously reported that aromatase is localized in the stroma (Hiramatsu et al. 1997) or in both epithelium and stroma in BPH (Matzkin and Soloway 1992). Aromatase activity and immunoreactivity, as well as aromatase mRNA, as evaluated by RTPCR, have also been detected in BPH, as well as in normal prostate and prostate cancer tissues (Stone et al. 1986; Matzkin and Soloway 1992; Hiramatsu et al. 1997). Preliminary results recently obtained in our laboratory suggest that the expression of 17β-HSD types 1, 2, 4, 7, and 12 and aromatase in normal and cancerous prostate tissues is similar to that found in BPH.
The enzymes aromatase and reductive 17β-HSDs are involved in the local synthesis of E2 in a variety of peripheral tissues, including the prostate (Luu-The 2001; Simpson and Davis 2001; Mindnich et al. 2004) (Figure 1). Aromatase catalyzes the conversion of androstenedione to E1, whereas reductive 17β-HSDs, including the types 1, 7, and 12, convert E1 to E2. On the other hand, the oxidative 17β-HSD types 2 and 4 convert E2 into the weak estrogen E1. On the basis of previous reports (El-Alfy et al. 1999; Pelletier et al. 1999) as well as the present localization studies, it then appears that enzymes involved in the formation and inactivation of E2 are expressed in both epithelial (including basal cells) and stromal cells. Recently, Harkonen et al. (2003) have reported that cultured prostate cancer LNCaP cells could produce E2, possibly by the reductive activity of 17β-HSD type 7, and could convert E2 to E1 through the action of 17β-HSD type 2. Inactivation of estrogens can also be induced by sulfonation and glucuronidation. The expression of the enzymes involved in sulfonation and glucuronidation in human prostate is currently under investigation. Estrogen sulfotransferase type 1 has been found to be expressed in epithelial cells in mouse prostate (Pelletier et al. 2004).
It is noteworthy that intense staining for 17β-HSD type 2 and aromatase could be observed at the apex of luminal cells. We have no explanation for such a localization. At the apex of luminal cells, there is generally an accumulation of vesicles in close proximity of the microvilli, which project into the alveolar lumen (El-Alfy et al. 2000). So far, there has been no report on the presence of enzymes involved in steroid metabolism in prostatic fluid. Clearly, more studies involving immunoelectron microscopy are required to clarify this point.
The role and mechanism of action of estrogens in human prostate is still unclear. There is accumulating evidence that estrogen itself can exert an influence on prostate cell division and differentiation in a variety of mammals. Estrogen administration to castrated or hypothysectomized dogs induced glandular hyperplasia (Leav et al. 1978; Tunn et al. 1979). In castrated rats, E2 administration induced moderate hypertrophy of epithelial cells and increased androgen receptor immunostaining in nuclei of epithelial and stromal cells (Pelletier 2002). The estrogen receptor (ER) belongs to the nuclear receptor superfamily, which also includes receptors for androgens, progestins, glucocorticoids, and mineralocorticoids, as well as thyroid hormone, retinoic acid, and vitamin D (Tsai and O'Malley 1994). Recently, a second ER has been cloned from rat prostate and called ERβ (Kuiper et al. 1996), and the original ER is now designated the ERα subtype. No other major ER types have been identified so far. In human prostate, ERβ is the predominant ER type (Enmark et al. 1997; Pelletier and El-Alfy 2000; Pasquali et al. 2001). By immunohistochemistry, ERβ was found to be expressed in both basal and secretory cells in alveoli, as well as in stromal cells, whereas no ERα could be detected (Bonkhoff et al. 1999; Pelletier and El-Alfy 2000). Interestingly, in BPH tissue, ERβ, but not ERα, has been detected in the transition zone, where BPH nodules develop (Tsurusaki et al. 2003). This suggests that E2 synthesized in stromal cells could act on ERβ by an autocrine mechanism to stimulate stromal cell proliferation.
(
(
(
(
(
(
In human, intraprostatic E2 concentration increases and DHT concentration decreases with age, accompanied by an increase in prostate volume (Seppelt 1978; Partin et al. 1991; Shibata et al. 2000). In fact, there is a significant correlation between the E2:DHT ratio and stromal hypertrophy. Serum E2 concentrations remain constant with aging (Rubens et al. 1974; Seppelt 1978; Shibata et al. 2000). Whether the age-related increase in intraprostatic E2 is due to an increase in local biosynthesis of E2 remains to be clarified. Because all the enzymes responsible for E2 biosynthesis from circulating DHEA are expressed in the prostate, it is tempting to speculate that locally produced E2 can modulate epithelial and stromal cell activity through interaction with ERβ in an intracrine fashion.
(
The physiopathological role of estrogens in the development of BPH and prostate cancer should be further investigated by evaluating the effects of pure antiestrogens and inhibitors of enzymes involved in estrogen biosynthesis in BPH patients. In one study, administration of tamoxifen, which exerts both antagonist and agonist activity at the ER level, provided only some improvement of urinary obstruction (Di Silverio and Tenaglia 1986). This lack of major improvement could be attributed to the agonist activity of tamoxifen. With the most widely used aromatase inhibitor, atamestane, only a minor reduction of urinary symptoms due to BPH could be observed (Schweikert et al. 1993; Radlmaier et al. 1996). A possible explanation could be that the reduced E2 synthesis is counterbalanced by a parallel increase in testosterone and DHT (Figure 1) (Gingell et al. 1995). The development of reductive 17β-HSD antagonists, which could be administered in combination with aromatase and 5α-reductase inhibitors, should provide a better nonsurgical treatment for BPH.
In summary, we report that several enzymes involved in E2 metabolism are expressed in epithelial and stromal cells in human prostate. It is hypothesized that estrogen can modulate the activity of these cell types by interaction with estogen receptors (probably ERβ) by an intracrine mechanism.