
Abstract
The present study showed that the HIS-C7 monoclonal antibody, which recognizes the chick form of CD45, is a specific marker for macrophages/microglial cells in the developing and mature chick central nervous system (CNS). HIS-C7-positive cells were characterized according to their morphological features and chronotopographical distribution patterns within developing and adult CNS, similar to those of macrophages/microglial cells in the quail CNS and confirmed by their histochemical labeling with Ricinus communis agglutinin I, a lectin that recognizes chick microglial cells. Therefore, the HIS-C7 antibody is a valuable tool to identify brain macrophage and microglial cells in studies of the function, development, and pathology of the chick brain. CD45 expression differed between chick microglia (as revealed with HIS-C7 antibody) and mouse microglial cells (as revealed with an antibody against mouse form of CD45). Thus, a discontinuous label was seen on mouse microglial cells with the anti-mouse CD45 immunostaining, whereas the entire surface of chick microglial cells was labeled with the anti-chick CD45 staining. The functional relevance of these differences between species has yet to be determined.
M
CD45 is a membrane-bound protein tyrosine phosphatase thought to be required for antigen-receptor signaling in cells of the immune system, although it has been proposed that CD45 may also play other roles (Penninger et al. 2001) including regulation of the proliferation, differentiation, and function of hematopoietic cells (Irie-Sasaki et al. 2003). CD45 antigen appears in all cells of monocyte/macrophage lineage and permanently labels them when they are outside the central nervous system (CNS) parenchyma (Penninger et al. 2001).
In mammals, anti-CD45 antibodies recognize ameboid and poorly ramified microglial cells in the developing brain, suggesting that microglial cells derive from precursors related to the monocyte/macrophage lineage. CD45 immunoreactivity decreases as microglial cells acquire features of mature microglia and increases again when microglial cells are activated (Herber et al. 2006). The decrease in immunoreactivity is not similar in all areas of the normal CNS, so that CD45 labeling of mature ramified cells varies among different regions. Therefore, anti-CD45 antibodies appear to label most microglial cells in human retinas (Provis et al. 1995) but fail to reveal the presence of mature microglial cells in many localizations of the mature human brain (Mittelbronn et al. 2001).
Most studies on the development and function of microglia in birds have been performed in the quail (Cuadros and Navascués 1998; Marín-Teva et al. 1999; Jeon et al. 2004; Sánchez-López et al. 2004,2005) as a consequence of the existence of the QH1 monoclonal antibody, which is a reliable marker of quail microglial cells (Cuadros et al. 1992b). In contrast, microglial studies in chicks have been scarce because of the lack of a specific marker for microglia in this species. The few reports on chick microglia revealed them by histochemical techniques (Fujimoto et al. 1987; Shin et al. 2003; Ignacio et al. 2005), which are less specific than immunocytochemical techniques. Availability of a reliable immunostaining of microglial cells in the chick would be a valuable tool for investigators of this model system. The possibility that microglia in the chick might express CD45, as occurs in mammals, and the commercial availability of the HIS-C7 monoclonal antibody (Jeurissen et al. 1988a), which recognizes the CD45 homolog in chicks, led us to use this antibody in the chick CNS to test its utility as an immunomarker for microglia in this species. Immunocytochemical analysis of developing and adult brains and retinas showed that HIS-C7 recognizes macrophages and all forms of microglial cells, including mature ramified ones, and can therefore be used to identify microglial cells in the chick.
Materials and Methods
Animals and Histology
Chick embryos from the 2nd to 20th day of incubation (E2-E20) and chickens aged 1–34 days after hatching (P1-P34) were used.
Young embryos (up to E7) were fixed for 2-4 hr after elimination of external membranes. Brains of older embryos were removed after decapitation and placed in fixative for 3-6 hr according to the age of the embryo. Posthatched animals were anesthetized with a mixture of ketamine and chlorbutol (Imalgene; Merial, Barcelona, Spain) and perfused with buffer followed by fixative; brains and eyes were then removed from the skull and immersed in the same fixative for 6-8 hr. Two different fixatives were used: 4% paraformaldehyde in 0.1 M PBS, pH 7.4, and 4% paraformaldehyde in 0.1 M cacodylate buffer, pH 7.4.
Quail embryos (14- to 16-days old) and 7- to 15-day-old mice (strain C57BL/6) were also used. Brains of quail and mice were processed as described above. All animal protocols followed guidelines approved by the Ethics Committee on Animal Experimentation of the University of Granada.
Fixed tissues thus obtained were frozen in liquid nitrogencooled isopentane and kept at −40C. Twenty-μm-thick sections were cut on a cryostat (CM 1850; Leica, Wetzlar, Germany), attached to superfrost slides (Menzel-Glaser; Braunschweig, Germany), and stored at −40C until use.
Antibodies and Immunocytochemistry
The primary antibodies used in this study, with their sources and dilutions, are listed in Table 1. Sections from chicken specimens were immunostained with monoclonal antibodies HIS-C7, which recognize a CD45 homolog in chickens (Jeurissen et al. 1988a); CVI-68.1, which labels chicken mononuclear phagocytes (Jeurissen et al. 1988b); and Lep-100, which labels a glycoprotein of the membrane of lysosomes (Lippincot-Schwartz and Fambrough 1986).
The secondary antibodies were biotin-conjugated anti-mouse and anti-rat IgGs (both from Sigma; St Louis, MO), FITC-conjugated anti-mouse IgG (Sigma), Alexa Fluor 488-conjugated goat anti-mouse IgG (Molecular Probes; Eugene, OR), and Cy3-conjugated goat anti-rabbit IgG (Amersham Biosciences; Buckinghamshire, UK). When biotin-conjugated secondary antibodies were used, a final step using extravidin-FITC or extravidin–peroxidase (both from Sigma) was performed.
For immunofluorescence techniques, sections were rehydrated in PBS, permeabilized for 15 min in 0.1% Triton X-100 in PBS, and incubated for 30 min in normal goat serum (Sigma) diluted 1:30 in 1% BSA in PBS. They were then incubated in the primary antibody for 24-40 hr at 4C. After washing in PBS, sections were placed in the corresponding secondary antibody for 2-3 hr. When a biotin-conjugated secondary antibody was used, a third step was added by incubating the sections in FITC-extravidin (dilution 1:100 in BSA-PBS) for 1 hr. Cell nuclei were labeled either with bisbenzimide (Hoechst 33324; Sigma) or propidium iodide (Sigma). Finally, sections were rinsed and coverslipped with antifading mounting medium (Vectashield; Vector Laboratories, Burlingame, CA).
Primary antibodies used
When immunoperoxidase labeling was performed, endogenous peroxidase activity was eliminated by incubating sections in 1-2% hydrogen peroxide for 30-45 min. Biotin-conjugated secondary antibodies were always employed in peroxidase techniques, and the sections were then incubated in peroxidase-extravidin (Sigma) diluted 1:200 in BSA-PBS. Peroxidase activity was revealed by using a nickel-enhanced diaminobenzidine reaction.
Some sections were used as negative controls by omitting the primary antibody. No specific staining was observed in these sections.
Cryostat sections from quail brains were immunocytochemically treated with HIS-C7; conversely, sections from chick brains were processed for QH1 (Pardanaud et al. 1987; Cuadros et al. 1992b) immunocytochemistry.
NDPase and Lectin Histochemistry
Brains from chick embryos were treated with diphosphatase (NDPase) histochemistry. After dissection, each brain was cut into several pieces and fixed for 4 hr in 4% paraformaldehyde in 0.1 M cacodylate buffer (pH 7.4) containing 5% sucrose at 4C. The tissue was then rinsed in 0.1 M cacodylate buffer containing 7.5% sucrose, and 60-μm-thick sections were obtained in a vibratome (Polaron; Watford, UK). They were incubated in a medium containing inosine 5′-diphosphate (Sigma) for 25 min at 38C (see Dalmau et al. 1998). The reaction was stopped by transferring the sections to buffer. Lead deposits were revealed by immersing sections in 2% ammonium sulfide for 2 min. Stained sections were mounted on slides, dehydrated, cleared, and coverslipped with DePeX mounting medium (DHB; Poole, UK). Sections incubated without substrate were used as negative controls.
Brains used for lectin histochemistry were fixed in 4% paraformaldehyde and frozen as described above. Cryostat sections were double labeled for HIS-C7 and Ricinus communis (RCA I) lectin. HIS-C7 immunostaining was performed as previously described using Alexa Fluor 488-conjugated goat anti-mouse IgG as secondary antibody. Subsequently, sections were incubated for 2 hr in biotin-conjugated RCA I lectin (Vector Laboratories) diluted 1:250 in PBS at 38C, and the presence of bound lectin was revealed with avidin-TRITC complex (Sigma) diluted 1:150 in PBS. Finally, sections were mounted as described for fluorescence techniques.
Immunolabeling of Mouse Sections
Sections from mouse brains were immunostained using either mouse-specific anti-CD45 antibody (clone IBL/3/16, reference MCA 1388; Serotec, Oxford, UK) diluted 1:10-1:50 in BSA-PBS or Iba1 antibody (concentration 3-4 μg/ml; gift from Y. Imai) that labels all forms of microglial cells in the mouse brain (Ito et al. 1998). Sections were then incubated in Alexa Fluor 488-conjugated goat anti-rat IgG for anti-mouse CD45 antibody and Cy3-conjugated goat anti-rabbit IgG for Iba1. Some sections were double immunostained with both antibodies by incubating them first in a solution containing the two primary antibodies at the concentrations reported above and then in a mixture of the two secondary antibodies.
Western Blot
Extracts from chick brains obtained using a Dounce homogenizer were washed with PBS and resuspended in 100 μl of lysis buffer (50 mM Tris-HCl, pH 8.0, 0.1 mM EDTA, 0.5% Triton X-100, 12.5 mM 2-mercaptoethanol) for 30 min on ice. Pellet was eliminated and sample buffer (50 mM Tris-HCl, pH 6.8, 6 M urea, 6% 2-mercaptoethanol, 3% SDS, 0.003% bromophenol blue) was added to the supernatant. Proteins were resolved on 7% SDS polyacrylamide gels and transferred onto Immun-Blot PVDF Membrane (Bio-Rad; Hercules, CA). Blots were blocked with PBS containing 5% milk powder and 0.1% Tween-20 for 30 min, washed with the same buffer without milk powder, and incubated overnight with the primary antibody. The primary antibodies were HIS-C7 (diluted 1:500) and anti-mouse CD45 (diluted 1:1000, reference MCA 1388; Serotec). Blots were then incubated for 2 hr in secondary antibodies (peroxidase-conjugated anti-mouse IgG for HIS-C7 and peroxidase-conjugated anti-rat IgG for anti-mouse CD45). Bands were visualized by ECL-PLUS (Amersham Biosciences), and pictures were taken using the ChemiDoc XRS System (Bio-Rad).
Results
Identification of HIS-C7-labeled Cells as Microglial Cells on the Basis of Their Morphology and Distribution in the Developing and Mature Chick CNS
Numerous HIS-C7-positive cells were always present in developing and mature brains of chicks. Morphological features and chronotopographical distribution pattern of these cells allow them to be characterized as macrophages and microglial cells.
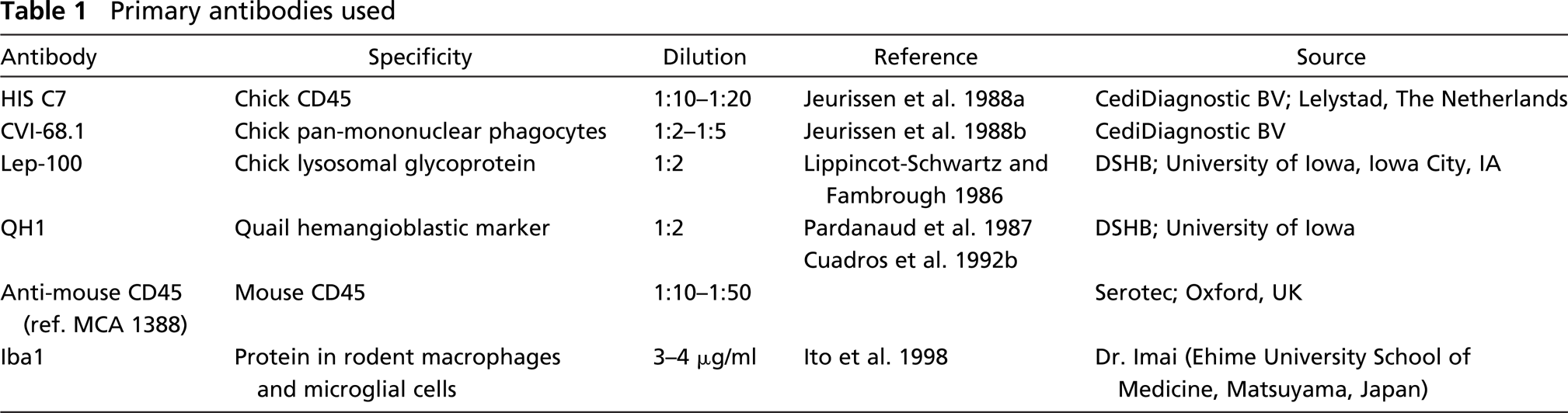
Cells labeled by HIS-C7 were present in the chick embryos as early as E3. At this age, many HIS-C7-positive cells appeared in the mesenchyme. Some were seen in the neuroepithelium (Figure 1A), with a progressive increase in the number of the latter during subsequent developmental stages (Figure 1B). Distribution pattern and morphological features of the labeled cells matched those reported for primitive macrophages in avian embryos (Cuadros et al. 1992a,1993). Thus, HIS-C7-positive cells appeared within the developing retina and vitreous (Figure 1C) and in the developing brain. In general, these cells were rounded or ovoid but sporadically presented an elongated morphology with short processes.
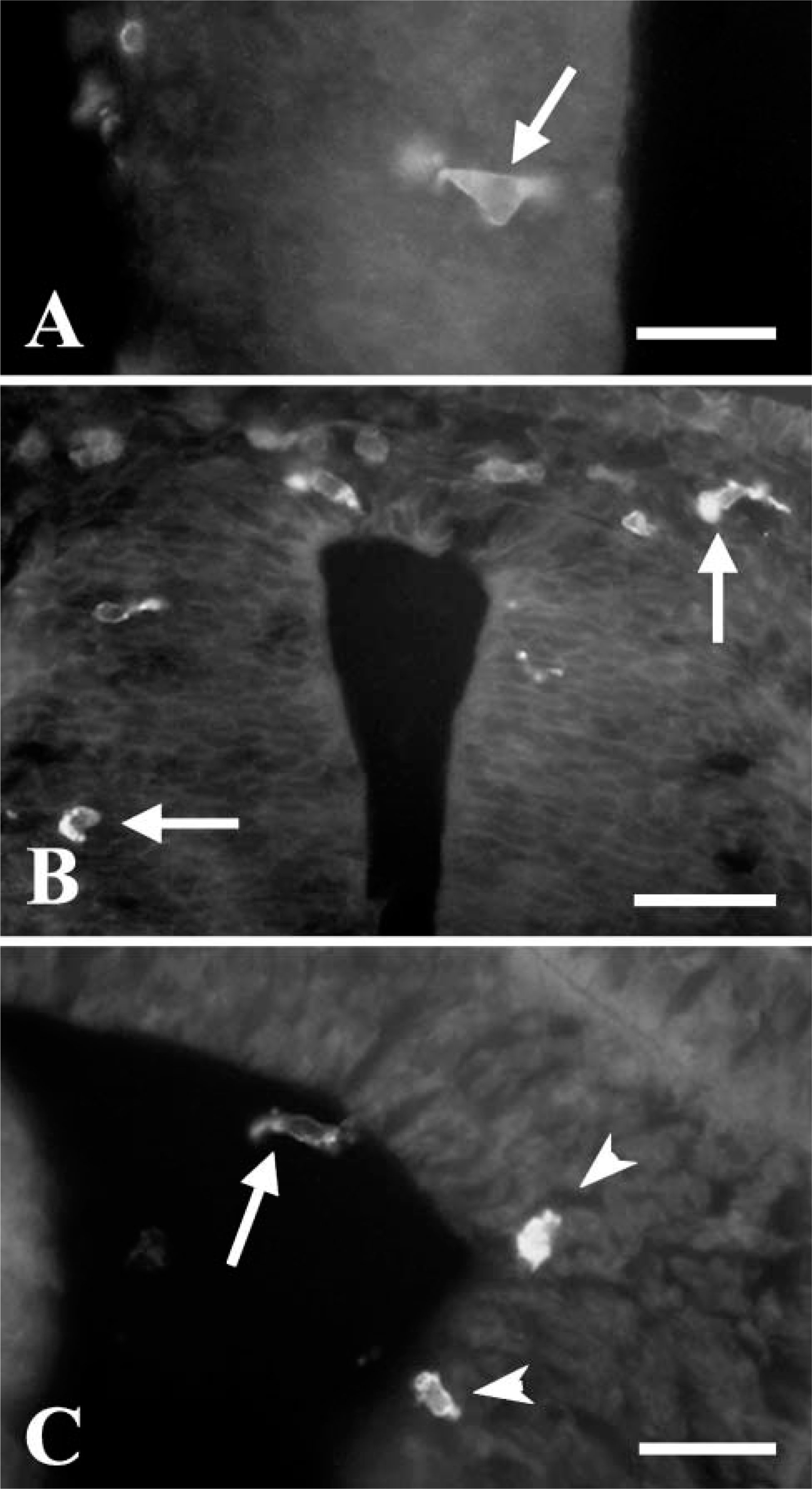
Two morphological types of HIS-C7-positive cells were clearly distinguishable within the developing CNS. Thus, in the optic tectum at E12 (Figure 2A), there were round or ameboid-shaped cells with some short pseudopods (Figure 2B) and another poorly ramified type (Figure 2C). The intensity of HIS-C7 staining also differed: staining of cells with ameboid morphology was generally stronger than cells with ramifications. As developmental time progressed, the proportion of ramified HIS-C7-positive cells increased, and the cells with ameboid morphology were progressively restricted to specific areas of the brain (e.g., around the ventricles or near blood vessels).
Fluorescent labeling of HIS-C7- (anti-CD45) positive cells in the neuroepithelium of incubation day (E3-E4) embryos. (
Posthatching, most HIS-C7-labeled cells showed increasingly complex ramifications throughout the nervous parenchyma (Figure 3A). The distribution pattern of these cells in different areas of the brain, e.g., developing optic tectum and cerebellum, matched that previously described for macrophages/microglial cells in the developing quail CNS (Cuadros and Navascués 1998).
HIS-C7-positive cells in E12 embryos as revealed with immunoperoxidase. (
Confocal microscopy of HIS-C7-positive cells in adult and embryonic chick central nervous system. (
During development, HIS-C7-positive cells were frequently near or in contact with pyknotic fragments (Figure 3B), suggesting their involvement in phagocytosis of apoptotic cell remains.
It is noteworthy that the composition of the fixative was critical for revealing the HIS-C7 labeling in some specimens. In fact, some sections of mature and developing brains showed no labeling with HIS-C7, and retinal sections of adult chicks fixed with paraformaldehyde in PBS consistently lacked any HIS-C7 immunoreactivity, although labeled cells were seen in the connective tissue surrounding the retina. However, strongly HIS-C7-positive cells were seen in all CNS regions, including the retina, when paraformaldehyde in 0.1 M cacodylate buffer was used during fixation instead of the usual paraformaldehyde in PBS (Figure 4).
Characterization of HIS-C7-labeled Cells as Microglial Cells on the Basis of Their Histochemical Labeling with NDPase and RCA I
Previous studies used detection of NDPase activity to characterize microglial cells in various species, including chick (Fujimoto et al. 1987) and mammals (Murabe and Sano 1982; Dalmau et al. 1998). In the present study, NDPase activity was detected in two types of cells in the developing chick CNS: some of the labeled cells exhibited no cell processes and had an irregular or round outline (not shown), whereas others showed a ramified morphology (Figure 5A). As observed in the HIS-C7-labeled cells, NDPase-positive ramified cells increased in number during development and were the most abundant cell with NDPase activity at E20. Double labeling of the same cell using HIS-C7 immunocytochemistry and NDPase histochemistry was precluded because the two methods need very different histological procedures. However, HIS-C7-positive cells and NDPase-positive cells had similar features (small cell body bearing long ramified processes) (Figure 5B), suggesting that the markers labeled the same cells.

Confocal microscopy showing HIS-C7-positive ramified cells in the retina of an adult chick fixed with paraformaldehyde in cacodylate. PE and V mark the location of pigment epithelium and vitreal space, respectively. ONL, outer nuclear layer. Bar = 25 μm.
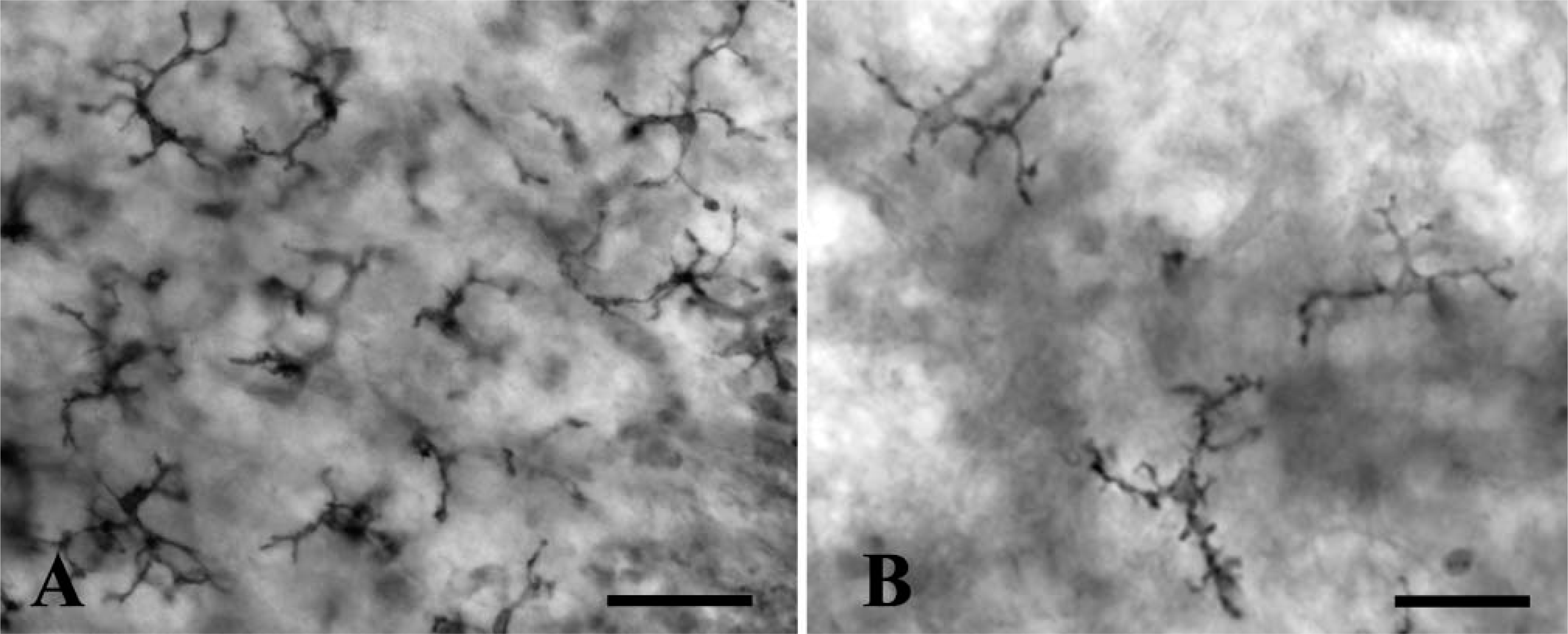
Ramified microglial cells labeled with NDPase in an E20 chick embryo. (
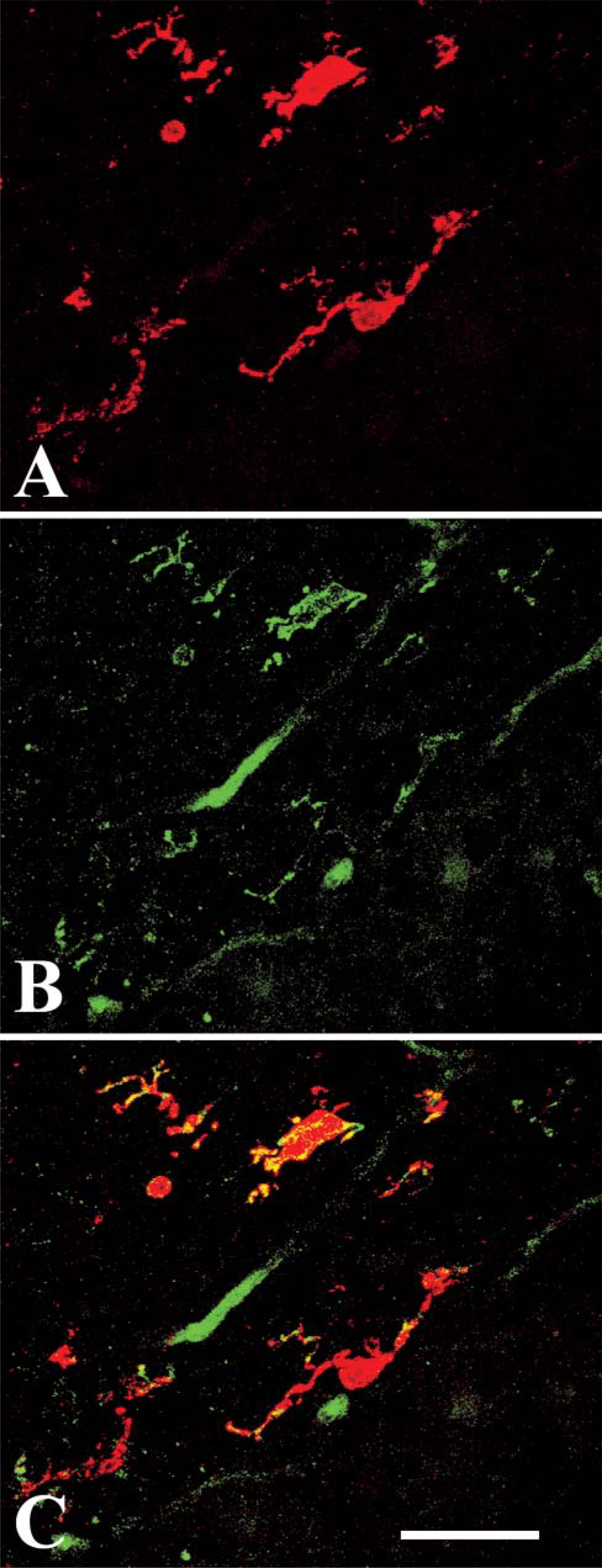
RCA I lectin, which labels microglial cells in the chick brain (Shin et al. 2003), was used to confirm that the HIS-C7-positive cells were microglia. Double-labeled sections revealed that HIS-C7 and RCA I recognize the same microglia in the nervous parenchyma (Figure 6). Blood vessels were also labeled by the lectin (Figures 6D-6F).
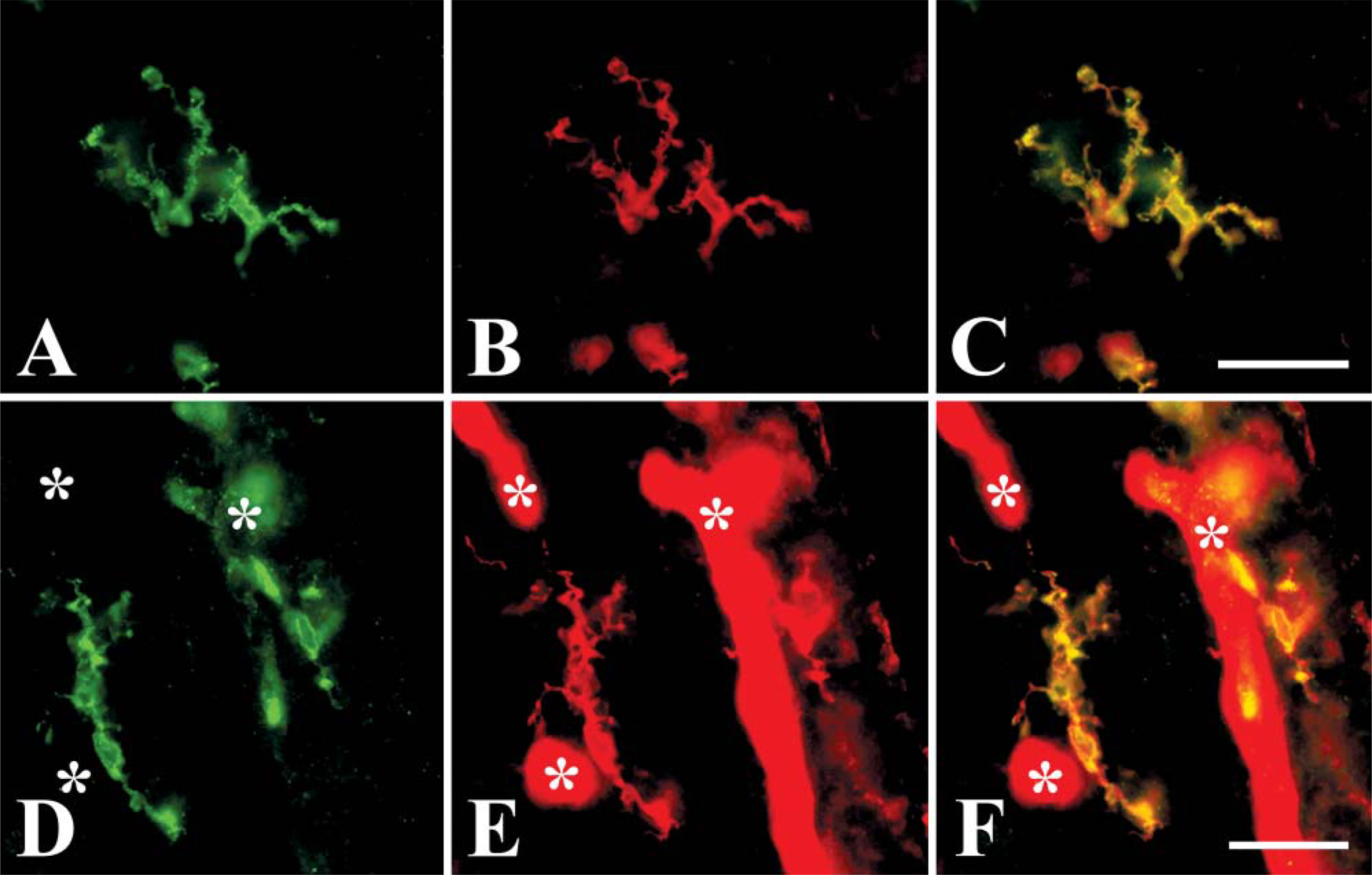
Cells labeled by HIS-C7 are also labeled by RCA I lectin. Pictures are taken from sections of the forebrain of E19 chick embryos. (
HIS-C7 Antibody Is Specific for an Antigen Present in the Chick Brain
Information from the commercial source of the HIS-C7 antibody states that it recognizes chicken CD45. Nevertheless, Western blot experiments were performed to obtain further information about the molecule recognized by HIS-C7 in the chick brain. The only band recognized by HIS-C7 had a molecular weight of 100-150 kDa (Figure 7A). This label did not appear in mouse brain extracts, whereas the antibody used to recognize mouse CD45 labeled a band in mouse brain extracts but not in chick brain extracts (Figure 7B).
CVI-68.1 and Lep-100 Immunolabelings Do Not Distinguish Microglial Cells
The antibody CVI-68.1 has been described as a marker of all mononuclear phagocytes in the chicken, including microglial cells (Jeurissen et al. 1988b). In our material, however, we were unable to stain microglial cells in the nervous parenchyma with this antibody, most likely due to technical reasons. Jeurissen et al. (1988b) used frozen sections from non-fixed tissue, which were subsequently briefly fixed with acetone, whereas we used the marker on sections from paraformaldehyde-fixed tissue. Few CVI-68.1-positive cells were seen within the brain in our preparations, usually within blood vessels, despite the fact that numerous cells were strongly labeled in the meninges and ventricles (not shown).
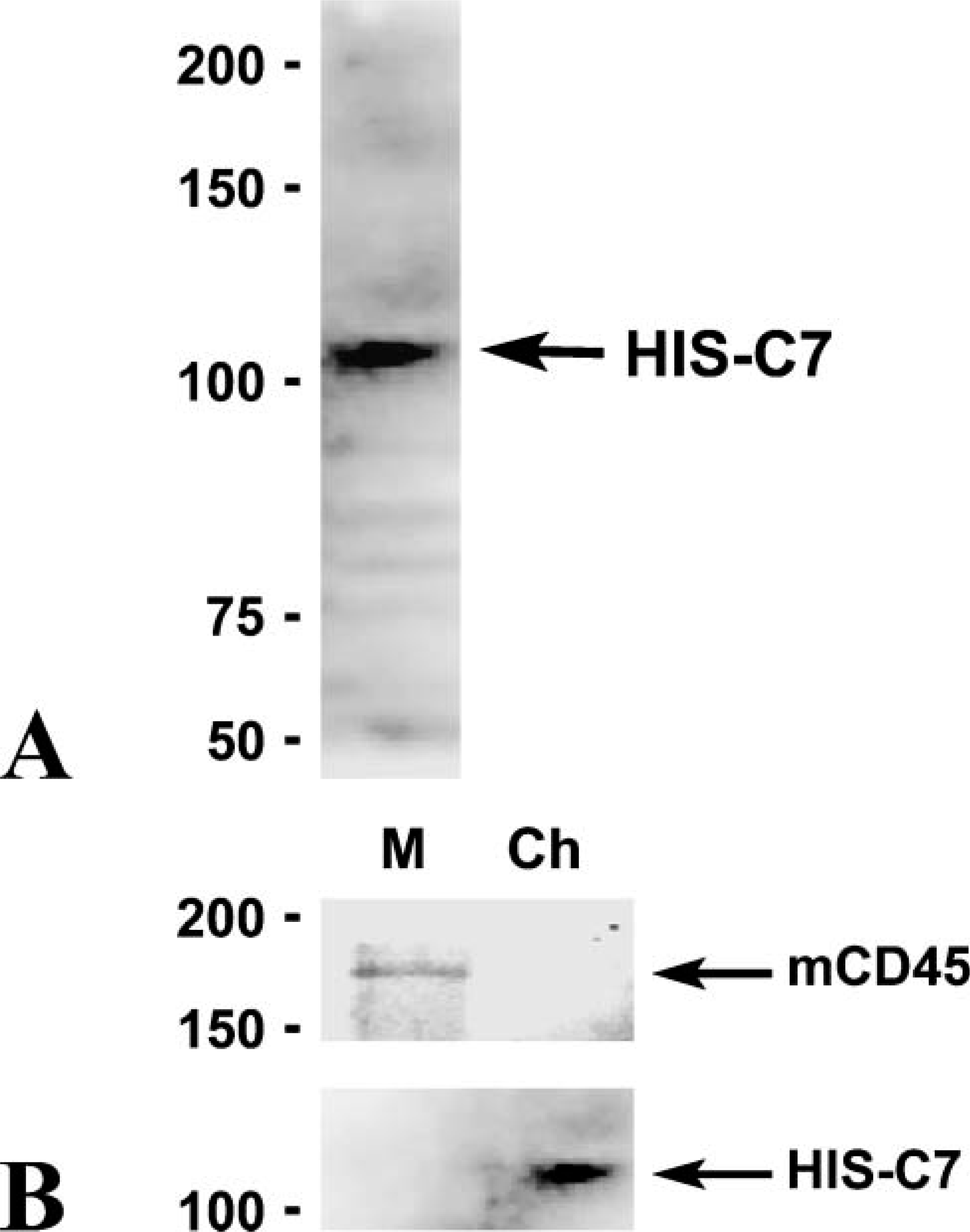
Western blots showing immunoreactivity of HIS-C7 and anti-mouse CD45 in extracts obtained from chick and mouse brains. Molecular weights are indicated in the left part of the figure. (
The Lep-100 antibody recognizes a glycoprotein of the lysosome membrane and has been used to label activated microglial cells in the chick retina after an injury (Fischer et al. 1998), although it does not distinguish microglial cells from other cell types in normal retinas. In the present study, Lep-100 immunostaining in the chick brain revealed the presence of lysosomes within the cytoplasm of small cells, many of them presumably microglial cells (Figure 8A). However, similar labeling was also present in the cytoplasm of neuron-like cells (Figure 8B).
HIS-C7 Does Not Label Microglial Cells in the Quail, and QH1 Does Not Label Microglial Cells in the Chick
Microglial cells were not labeled in HIS-C7 immunostained sections from quail brain, although some faintly labeled cells were observed in the meninges and brain parenchyma (not shown). Conversely, immunocytochemistry with the QH1 antibody, which recognizes all forms of microglial cells in the quail (Cuadros et al. 1992b), showed no labeling on chick brain sections with negative results (not shown).
CD45 Immunoreactivity in Mice
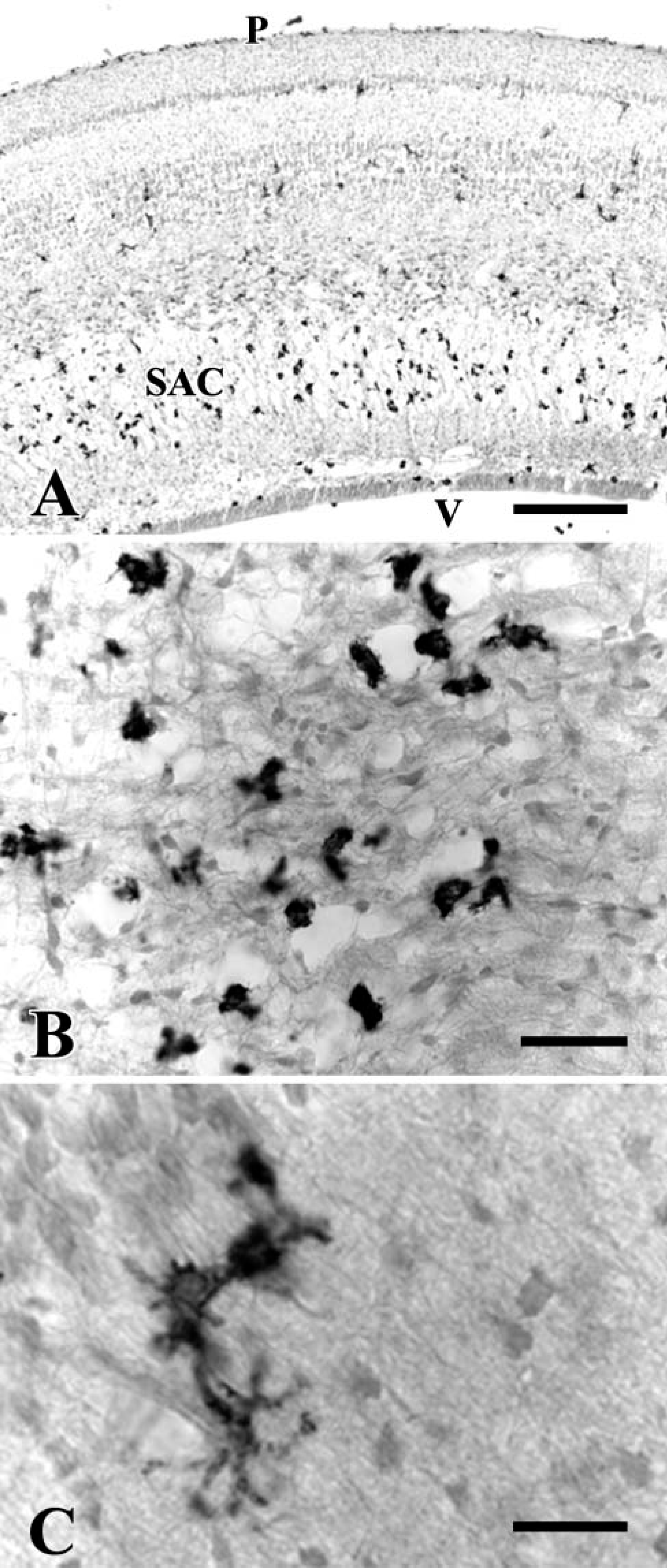
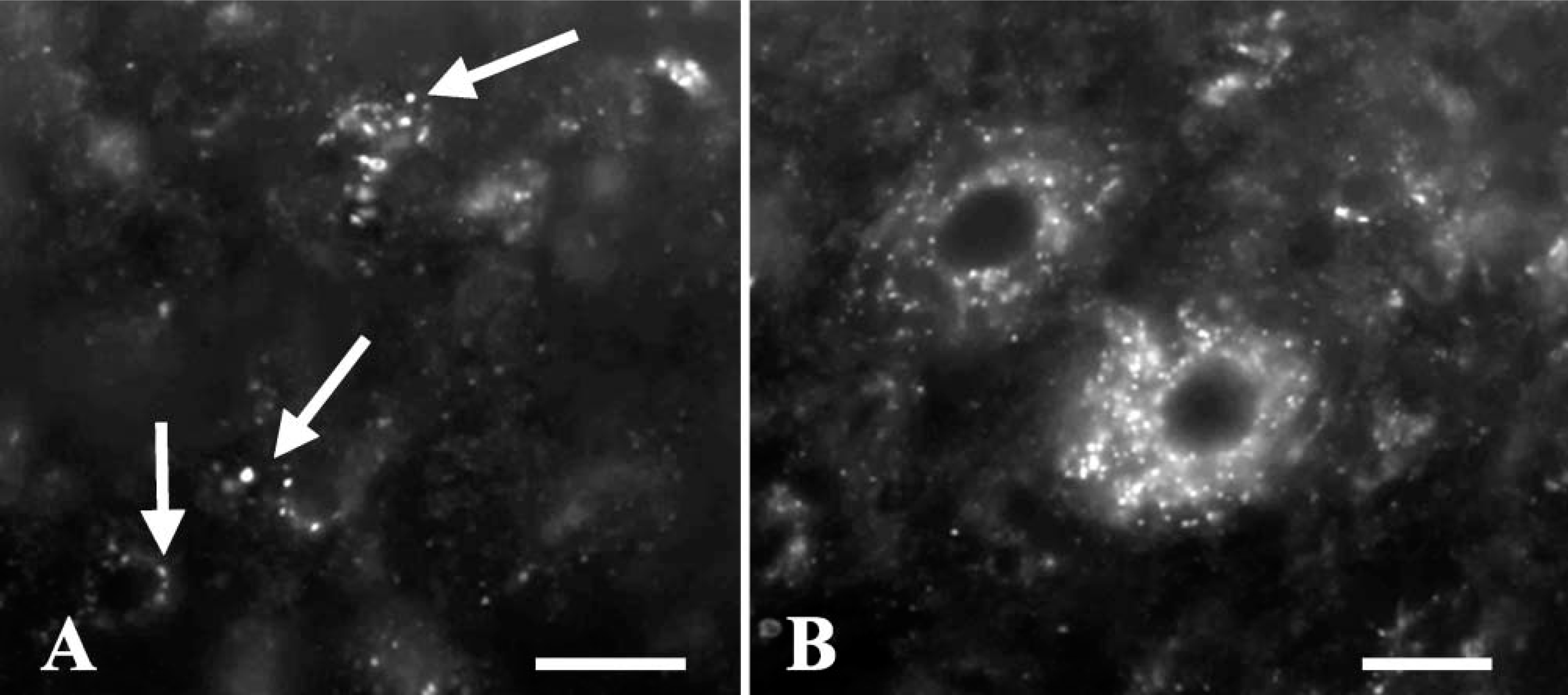
To determine whether the CD45 immunolabeling is also a reliable tool to recognize microglial cells in other species, sections from developing mouse CNS were immunostained with a specific antibody against mouse CD45. Microglial cells in postnatal and adult mouse brains were not clearly labeled: some cells were labeled by the antibody in some areas of the CNS (Figure 9A) but not in others (Figure 9B). The endothelium of some brain vessels was also immunolabeled.
To confirm that the anti-CD45 antibody does not mark all microglial cells in the mouse brain, some sections were double labeled with anti-CD45 and Iba1 antibodies. This latter is a specific marker of microglial cells in rodents (Ito et al. 1998). In some brain areas, anti-CD45 and Iba1 antibodies recognized the same cells, except for the CD45 labeling of some Iba1-negative endothelial cells. However, in other areas (many regions of the forebrain, cerebellum, and pons), Iba1 staining enabled the identification of microglial cells that were not labeled or were only faintly labeled by anti-CD45 immunostaining (Figure 10). After careful examination, it was found that most Iba1-positive cells were also labeled by the anti-CD45 antibody, although the staining was patchy and weak and usually only visible after considerable amplification of the signal produced by the fluorescent marker.
Lep-100 immunolabeling of lysosomes in the P8 chick brain. (
Analysis was conducted of brains and retinas of mice fixed in paraformaldehyde in cacodylate buffer to test whether the use of this fixative allowed mouse microglial cells to be labeled by anti-CD45. Normal Iba1 immunoreactivity was observed in this material, but no specific labeling with the anti-CD45 antibody was found (not shown).
Discussion
This study clearly showed that HIS-C7-positive cells were present within the neuroepithelium of the chick embryo at E3. Before this age, HIS-C7-positive cells were observable in the mesenchyme and other nonneural structures, as already reported (Jaffredo et al. 1998). Labeled cells in the neuroepithelium showed a similar distribution to that previously described for early macrophages in avian embryos (Cuadros et al. 1992a, 1993). Process-bearing HIS-C7-positive cells became increasingly ramified with longer developmental time, with the result that ramified cells were virtually the only HIS-C7-positive cells to appear in the CNS at the second week after hatching. The morphology and chronotopographical distribution pattern of HIS-C7-labeled cells strongly suggest that this anti-CD45 antibody specifically recognizes macrophages/microglial cells throughout development and adulthood. Whereas the first labeled cells seen in the chick embryo neuroepithelium appear to be early macrophages invading the avian CNS during the first days of embryonic development (Cuadros et al. 1993), the poorly or entirely ramified labeled cells in more developed brains would correspond to microglial cells at different stages of their differentiation.

Microglial cells in the mouse brain as revealed with peroxidase anti-CD45 immunocytochemistry. (
Confocal microscope images showing that not all microglial cells in the forebrain of a P7 mouse are labeled by CD45 in mice. All microglial cells were stained with Iba1 (red), whereas incomplete or no staining was obtained with the anti-CD45 antibody (green). (
To test this proposition, we performed histochemical detection of NDPase activity, which labels all forms of microglial cells (Murabe and Sano 1982; Fujimoto et al. 1987; Dalmau et al. 1998) in different parts of the CNS. Cells showing NDPase activity (round/ameboid and ramified cells) were similar to those labeled after anti-CD45 immunostaining. Although CD45 immunocytochemistry and NDPase histochemistry cannot be performed on the same section, both techniques revealed cells with similar morphological features and distribution. Hence, these techniques appear to stain the same cell type. In addition, double labeling with HIS-C7 and RCA I lectin, a marker of microglial cells in the chick (Shin et al. 2003), revealed that the same cells were marked by the antibody and the lectin, demonstrating that the HIS-C7-labeled cells were macrophages/microglia. In contrast, the HIS-C7 labeling was clearly different from that shown by astrocyte and oligodendrocyte markers in the developing chick brain (Linser and Perkins 1987; Kalmán et al. 1998). Therefore, HIS-C7-labeled cells in the chick embryo can be considered macrophages or microglial cells.
Although the molecular characterization of the antigen recognized by HIS-C7 was not the aim of this paper, our Western blot data show that the HIS-C7 antibody recognizes a molecule with a molecular weight of 100-150 kDa. A previous study (Paramithiotis et al. 1991) showed that HIS-C7, in common with other antibodies to chicken leukocyte antigens, recognizes chick CD45, and different anti-chick CD45 antibodies bind molecules of different molecular weights. One of these antibodies, LT40, recognizes an antigen with a molecular weight of 150-200 kDa (Paramithiotis et al. 1991), whereas the molecule recognized by HIS-C7 in the chick brain has a molecular mass <150 kDa. These findings strongly suggest the presence of different forms of CD45 in the chick.
All forms of microglial cells appear to be labeled by the HIS-C7 antibody in the chick. Other antibodies identifying macrophages such as CVI-68.1 cannot be used as microglial markers in the chick because expression of the molecule they recognize is absent in microglial cells. Lep-100 labeling of the lysosomal compartment was previously used to show the presence of activated microglial cells within the injured chick CNS (Fischer et al. 1998). Our results demonstrate that this labeling does not allow microglial cells to be unequivocally identified during normal development of the CNS because of the labeling of other non-microglial cells.
We conclude, therefore, that the HIS-C7 antibody can be used as a specific marker for microglia in the developing and adult chick and constitutes a useful tool for detecting this type of cell in this species, which is widely used in research and, unlike mammals, is easy to manipulate throughout development, including its first stages. To the best of our knowledge, no specific marker for chick microglia has previously been described. In this study we used the only two reagents described in the literature for labeling microglial cells in chick material: lectin staining, which also labels endothelial cells and therefore does not allow an unequivocal identification of microglial cells, and NDPase histochemistry, which requires specific technical procedures that give rise to preparations of poor morphological quality. HIS-C7 immunostaining demonstrates none of these problems and is therefore suitable for the specific labeling of microglia in morphological studies of normal and pathological chick CNS.
A surprising finding of this study was that HIS-C7-labeled cells were absent from some sections from material fixed in paraformaldehyde in PBS but present in others of comparable age. The reasons for these differences in immunoreactivity are not clear, although they might be due to differences in specimen size and fixation time, with a longer fixation time required by larger than smaller pieces. We recommend a prolonged fixation (overnight) when this does not interfere with the detection of other molecules. We found that we could circumvent this technical problem by using paraformaldehyde in cacodylate buffer, which permitted shorter fixation times. The adult retina was consistently devoid of HIS-C7 labeling when paraformaldehyde in PBS was used, despite the clear labeling of macrophages in the connective tissue. In contrast, abundant HIS-C7-positive microglia were seen in the adult retina when it was fixed in paraformaldehyde in cacodylate buffer. The reason for the strong CD45 labeling of microglia in paraformaldehyde-cacodylate-fixed retinas has yet to be established.
Expression of CD45 in cells of hematopoietic lineage is not unexpected. However, the fact that this expression is sustained in microglial cells during development and adulthood has not previously been reported. Labeling of specific microglial cells by anti-CD45 antibodies has been described in other species. In humans, CD45 is weakly expressed in the developing spinal cord and is no longer present at adulthood, although positive cells are detected in the spinal cord of patients with motor neuron diseases (Rezaie et al. 1999). Anti-CD45 antibodies appear to label most microglial cells in the human retina (Diaz-Araya et al. 1995a,b) but recognize only a proportion of microglial cells in the human brain (Mittelbronn et al. 2001). Anti-CD45 antibodies also label microglia, although less strongly, in the brain of rat (Karp et al. 1994) and mouse (this study). It appears from the present observations that the anti-CD45 antibody used in this study recognizes most microglial cells in the mouse, but this labeling is not strong enough to distinguish microglial cells. A study by Karp et al. (1994) emphasized that CD45 is not present in all microglial cells and that its labeling may consequently be useful to distinguish different subpopulations of microglial cells. Therefore, it appears that labeling of microglia with anti-CD45 antibodies differs among species as well as among different CNS regions in a single species. These differences may result from a variation in the amount of recognized molecules in microglial cells but can also depend on other factors. Thus, modifications in the recognized molecule may render it more sensitive to fixative or hide the epitope that binds the antibody. Moreover, the use of different antibodies recognizing different epitopes or spliced variants of CD45 in these studies raises the possibility that the reported differences in CD45 expression depend in part on the heterogeneous expression of different forms of the same molecule. At any rate, CD45 immunolabeling of microglia is increased in the pathological CNS (Rezaie et al. 1999; Penninger et al. 2001), including experimentally injured brains (Karp et al. 1994; Wilcock et al. 2001, Herber et al. 2006) and those affected by Alzheimer disease (Tan et al. 2000). Some authors (Mott et al. 2004) have proposed that CD45 expression is related to the regulation of microglial activation.
Flow cytometry has revealed that cells expressing different levels of CD45 are present in the brain of normal rodents. It has been proposed that parenchymal microglia (ramified cells located in the nervous parenchyma) and perivascular microglia (ameboid or poorly ramified cells located immediately behind the basal lamina of brain capillaries) are distinguished by their CD45 expression. Thus, whereas low CD45 expression is shown by parenchymal microglia, high levels of CD45 expression are observed in perivascular microglia (Sedgwick et al. 1991) and microglial cell precursors (Walker 1999). The low expression of CD45 in parenchymal microglia reported in flow cytometry studies means that many of them might not be labeled by CD45 immunocytochemistry.
Further research is required to establish the functional implications of the presence in chick microglia of a sufficiently high CD45 expression to be recognized by immunocytochemical methods. At any rate, this expression can be used to specifically label microglial cells of the chick CNS.
Footnotes
Acknowledgements
This work was supported by grant BMC2001-3274 from the Spanish Ministry of Science and Technology.
Monoclonal antibodies QH-1 and Lep-100 were developed by F. Dieterlen-Lièvre and D.M. Fambrough, respectively, and obtained from the Developmental Studies Hybridoma Bank under the auspices of the National Institute of Child Health and Human Development and maintained by the University of Iowa, Department of Biological Sciences, Iowa City. We are grateful to Prof. Y. Imai (Ehime University School of Medicine, Matsuyama, Japan) for the gift of the Iba1 antibody. The authors thank Richard Davies for correcting the English style of the manuscript.