
Abstract
We present the generation of a panel of monoclonal antibodies (F55A10, F55A12, F64A6B4, and F65A2) against the homeodomain transcription factor Nkx6.1, one of the essential transcription factors that regulates the multistep differentiation process of precursor cells into endocrine β-cells in the pancreas. Expression of Nkx6.1 can be detected in developing pancreatic epithelium and in adult insulin-producing β-cells, making this transcription factor a unique β-cell marker. For production of monoclonal antibodies, RBF mice were immunized with a GST-Nkx6.1 fusion protein containing a 66-amino acid C-terminal fragment of rat Nkx6.1. Four clones were established as stable hybridoma cell lines and the produced antibodies were of the mouse IgG1/κ subtype. When applied for immunohistochemistry on frozen sections of adult mouse pancreas, monoclonal antibodies stain specifically the β-cells in the endocrine islets of Langerhans with patterns comparable to that of a previously produced polyclonal rabbit serum. Monoclonal antibodies can be divided into two groups that appear to recognize different epitopes, as determined by competition ELISA. The presented antibodies are useful tools for the further characterization of the role and function of Nkx6.1 in pancreatic development, especially for use in double-labeling experiments with existing polyclonal rabbit antibodies.
T
In developing spinal cord of mouse, Nkx6.1 plays an important role in dorsoventral patterning of the neural tube, where its expression domain defines progenitor cells for V2, MN, and V3 classes of ventral neurons (Briscoe et al. 2000; Sander et al. 2000a). It functions here as a repressor of Dbx1, Dbx2, and Nkx6.2, which all have expression domains immediately dorsal to Nkx6.1 in the neural tube (Vallstedt et al. 2001). Nkx6.1-deficient mice fail to generate V2 interneurons and motor neurons (Sander et al. 2000a).
In the pancreas, Nkx6.1 deficiency results in severely reduced numbers of β-cells after E13.5, whereas development of the pancreatic buds at earlier stages appears to proceed normally despite the lack of Nkx6.1 in the epithelium (Sander et al. 2000b). Thus, Nkx6.1 is essential for formation and differentiation of normal β-cells but is dispensable for the developmental process of pancreas ontogeny. In parallel to what has been observed in the spinal cord, Nkx6.1 acts as a repressor of Nkx6.2 expression in developing pancreatic tissue (Henseleit et al. 2005). Furthermore, the presence of Nkx6.1 expression is necessary for normal expression levels of the zinc-finger protein myelin transcription factor-1 in endocrine precursor cells (Henseleit et al. 2005). Lack of Nkx6.1 results in derepression of Nkx6.2, suggesting that some functional redundancy between the two Nkx6 factors exists. The β-cell phenotype is even more severe in Nkx6.1/Nkx6.2 double-mutant mice, but the early pancreatic epithelium still seems to develop normally until the onset of the secondary transition and does not reveal the role of Nkx6 transcription factors at the early stages (Henseleit et al. 2005).
Antibodies against Nkx6.1 are invaluable tools for the study of the role and function of this transcription factor in pancreatic development. The central role of the Antibody Core facility within the Beta Cell Biology Consortium is to develop and provide antibody tools for important pancreatic cell differentiation markers. We present here the generation of a panel of novel mouse monoclonal antibodies against Nkx6.1.
Materials and Methods
Expression and Purification of GST-Nkx6.1
A fusion protein containing glutathione-S-transferase (GST) and Nkx6.1 that had previously been utilized for the production of a polyclonal rabbit antiserum was used as the antigen for generation of the mouse monoclonal antibodies (Jensen et al. 1996). The plasmid, kindly provided by Susan Brenner-Morton (Columbia University, New York, New York), encodes a 66-amino acid (aa) fragment of the C terminus of rat Nkx6.1 (aa 299-365) in-frame with GST in the pGEX-4T plasmid. The C termini of the rat and mouse Nkx6.1 proteins are 100% identical. Purification of the fusion protein was performed as previously described (Jensen et al. 1996), using the RediPack GST purification module (Amersham Biosciences; Uppsala, Sweden).
Immunization
To obtain monoclonal antibodies, four RBF mice (Taggart and Samloff 1983) were immunized IP with 20 μg GST-Nkx6.1 biweekly. The first immunization was performed with complete Freund's adjuvant, and this was followed by two injections in incomplete Freund's adjuvant. Ten days after the third immunization, the animals were eye bled, and the antibody response was measured by ELISA.
Cell Fusion
The mouse selected for generation of monoclonal antibodies was boosted IV with 10 μg antigen in saline. Three days later the spleen was harvested and used for hybridoma production. The 2.3 × 108 spleen cells were fused to 5.24 × 107 FOX-NY myeloma cells (Taggart and Samloff 1983) using polyethyleneglycol 1500 (Roche; Hvidovre, Denmark) as previously described (Harlow and Lane 1988). Fused cells were initially seeded in 96-well tissue culture plates, and clones that tested positive for the production of anti-Nkx6.1 antibodies were subsequently subcloned by limiting dilution. All animal experiments were performed according to Danish National Research Council guidelines.
Antibody Purification
Isotypes of the antibodies were determined using the Mouse Monoclonal Antibody Isotyping Kit (Boehringer Mannheim; Hvidovre, Denmark) according to the manufacturer's instructions. All antibodies were IgG1 subtype and were purified from culture supernatant by use of a protein A Sepharose 4 Fast Flow column (Amersham Biosciences). The pH of the filtered supernatants was adjusted to 8.5 and the solutions supplemented with 3 M NaCl. Samples were applied to the column, initially equilibrated, and subsequently washed with 50 mM Tris, 3.5 M NaCl, pH 8.5. Elution was performed with 25 mM citric acid, 4.5 mM tri-natrium citrate, pH 3. Pooled eluates were dialyzed against PBS, and 0.05% NaN3 was added to the purified antibodies. Antibody concentrations were determined to be between 0.3 and 0.6 mg/ml by measuring the absorbance at OD280.
ELISA
ELISA plates were coated overnight (O/N) with 1 μg/ml GST-Nkx6.1, GST-Nkx6.2 (kindly provided by Maike Sander, UC Irvine, Irvine, CA), or pure GST in PBS. Washing and blocking were performed with PBS, 0.05% Tween 20. Samples diluted in PBS, 0.5% Tween 20, 1 M NaCl, pH 7.2, were added, and the plates were incubated for 1 hr at room temperature (RT). The presence of antigen-specific antibodies was detected with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Caltag; Hamburg, Germany) diluted 1:2000. One hundred μl of 3,3′,5,5′-tetramethylbenzidine (TMB; Kem-En-Tec, Copenhagen, Denmark) was added to each well, and the reaction was stopped 10 min later with 100 μl 4MH3PO4. Absorbance of the ELISA plates was read at 450 and 620 nm. For epitope mapping, the purified monoclonal anti-Nkx6.1 antibodies were biotinylated with N-hydroxy-succinimide-d-biotin (Sigma; Brøndby, Denmark) and diluted in dimethylformamide. Then, 7.7 μg biotin was incubated on a shaker for 4 hr at RT with 1 mg of the purified antibodies in 0.1 M NaHCO3, pH 8.5. Biotinylated antibodies were dialyzed against PBS and used in the direct ELISA at a concentration of 300 ng/ml in the presence of varying amounts of non-biotinylated antibodies. Binding of the biotinylated antibodies was detected using streptavidin-peroxidase (KPL; Gaithersburg, MD) at a 1:20,000 dilution.
Immunohistochemistry
Animal sera, hybridoma supernatants, and purified antibodies against Nkx6.1 were analyzed by immunohistochemical staining of frozen sections from either adult mouse pancreas fixed O/N in 4% PFA in PBS at RT or E11.5 embryos fixed O/N at 4C in Lilly's formalin buffer, pH 7.4, cryoprotected in 30% sucrose in PBS, embedded in Tissue-Tek (Sakura; Værlose, Denmark), and cut at 8-μm sections on a Leica cryostat. Antigen retrieval was performed by microwave treatment for 4 min at 600 W in 200 ml 0.01 M citrate buffer, followed by 15 min at 250 W, and finally left to cool for 20 min. Tissue sections were rinsed in PBS, quenched in 3% H2O2 for 5 min, and rinsed again. Then, 0.1 M Tris-HCl, pH 7.5, 0.15 M NaCl, and 0.5% blocking reagent (TNB; PerkinElmer, Hvidovre, Denmark) were added and the sections blocked for 30 min before O/N incubation with the purified mouse anti-Nkx6.1 antibodies diluted 1:1000 and rabbit anti-Nkx6.1 antiserum diluted 1:4000 in TNB blocking buffer. Sections were washed three times for 5 min each in PBS between incubations with primary antibody and the following incubations with biotinylated secondary antibody (Zymed; Aarhus, Denmark) for 30 min and streptavidin-peroxidase conjugate (Zymed) for 15 min. Bound antibodies were visualized with the Cy3 Fluorophore Tyramide Amplification agent (PerkinElmer) diluted 1:100 after a 10-min incubation period and a subsequent wash in PBS. Images were collected with an Olympus (Tokyo, Japan) BX51 microscope and captured using a chilled Hamamatsu C5810 CCD camera (Hamamatsu City, Japan) and Image Pro Plus 4.5 software and by confocal microscopy (Zeiss LSM 510 Meta; Birkerod, Denmark). Triple stainings were developed subsequently by adding guinea pig anti-insulin (1:100) (ABCAM; Cambridge, UK) and rabbit anti-glucagon (1:100) (ABCAM) O/N and visualized with anti-guinea pig Alexa 488 and anti-rabbit Cy5 (Jackson ImmunoResearch; Soham, UK), respectively. Images were recorded using confocal microscopy (Zeiss LSM 510). Preabsorption with GST fusion proteins and the polyclonal rabbit anti-Nkx6.1 antiserum has been described previously (Jensen et al. 1996).
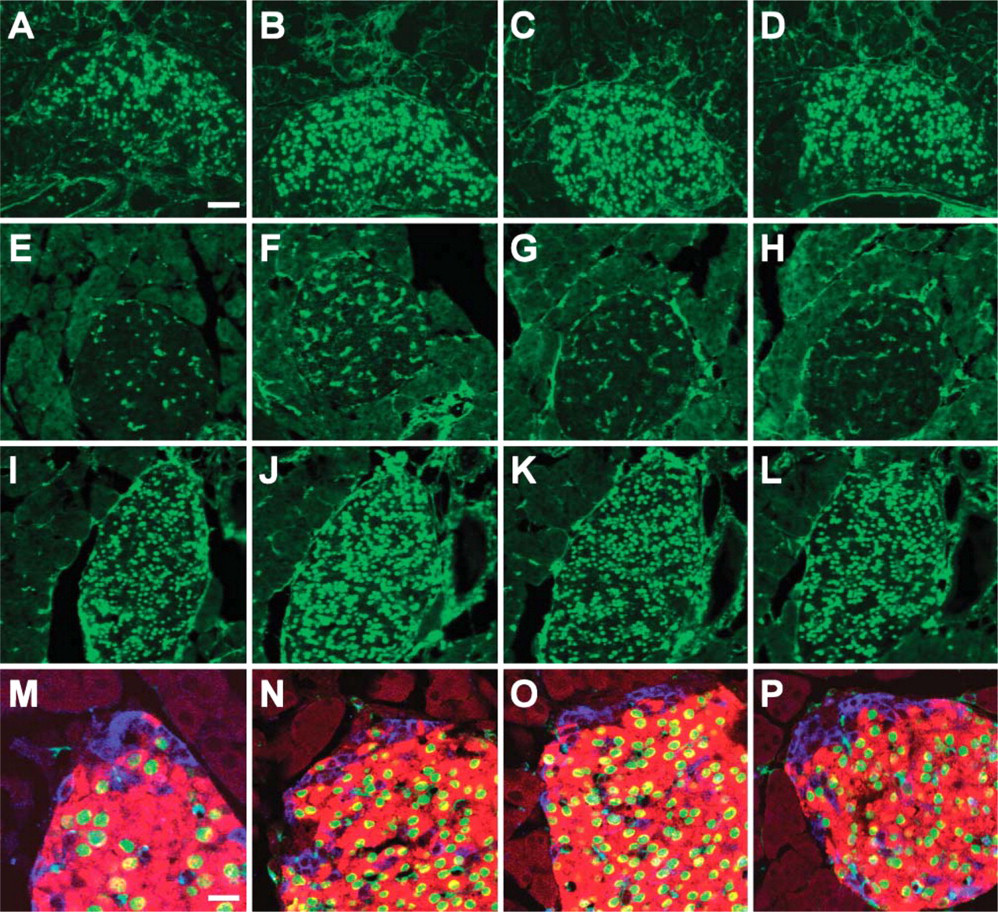
Antigen-specific anti-Nkx6.1 monoclonal antibodies recognize β-cell nuclei. Immunohistochemical stainings of PFA-fixed frozen sections of adult mouse pancreas with F55A10 (
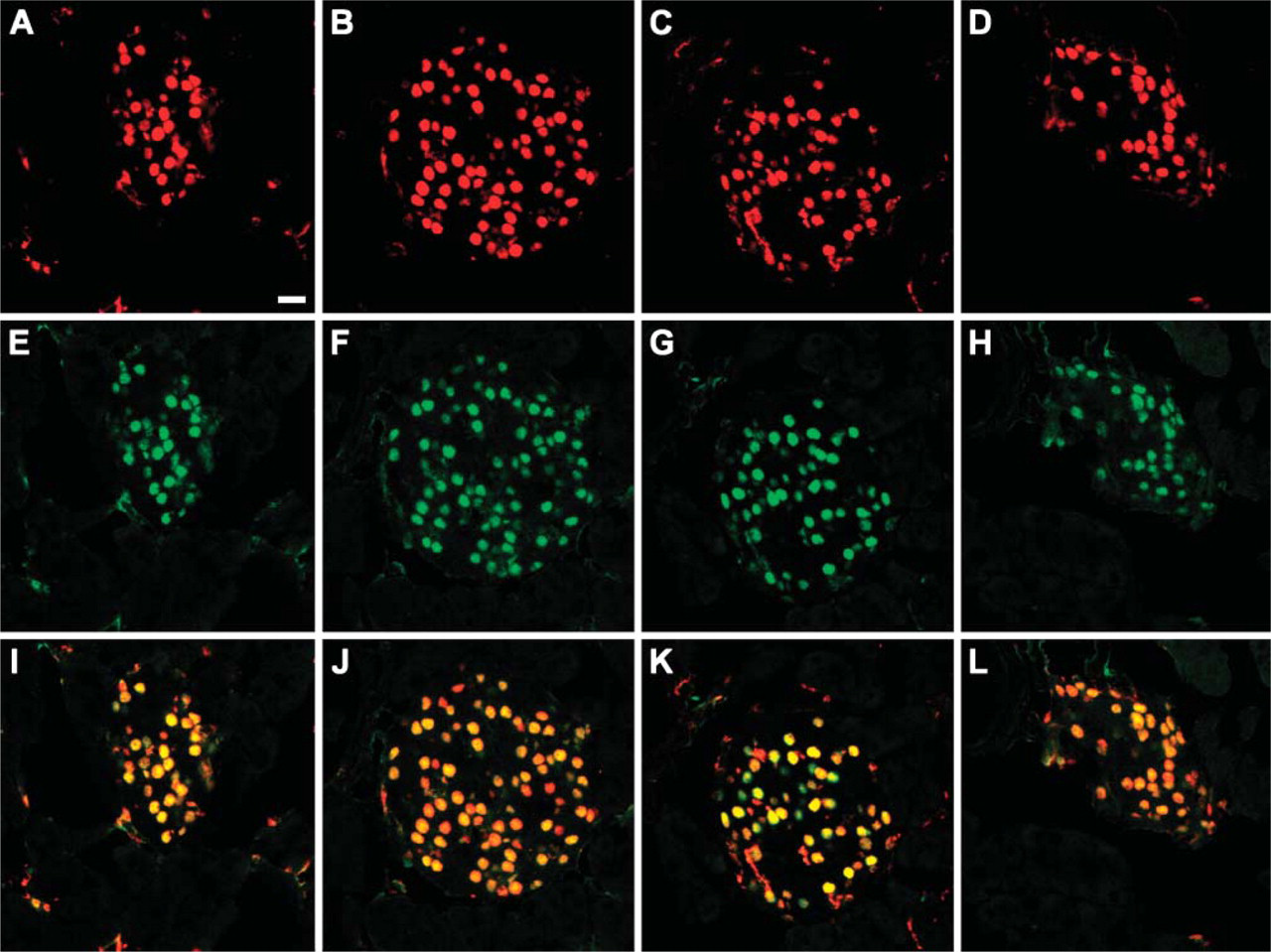
Mouse and rabbit anti-Nkx6.1 antibodies show matching staining patterns in adult islets. Double immunohistochemical stainings of adult mouse pancreas with a previously characterized polyclonal rabbit anti-Nkx6.1 antiserum (Jensen et al. 1996) (green) together with F55A10 (
Western Blots
One μg GST-Nkx6.1 and 4 μg GST-Nkx6.2 were loaded on NuPAGE SDS PAGE gels, run alongside the SeeBlue Plus2 prestained Standard (Invitrogen; Taastrup, Denmark), and blotted onto a nitrocellulose membrane using the NuPAGE MOPS buffer system. Nitrocellulose was washed in washing buffer (PBS supplemented with 50 mM Tris, 150 mM NaCl, 0.5% Tween 20) and blocked for 20 min at RT in washing buffer containing a total of 2.5% Tween 20. Monoclonal anti-Nkx6.1 antibodies were diluted to a concentration of 0.5 μg/ml before O/N incubation at 4C. A HRP-conjugated rabbit anti-mouse IgG (DAKO; Glostrup, Denmark) was used as the secondary antibody at a dilution of 1:2000 for 1 hr at RT and detected using TMB Blotting Ready-to-Use Substrate (Kem-En-Tec).
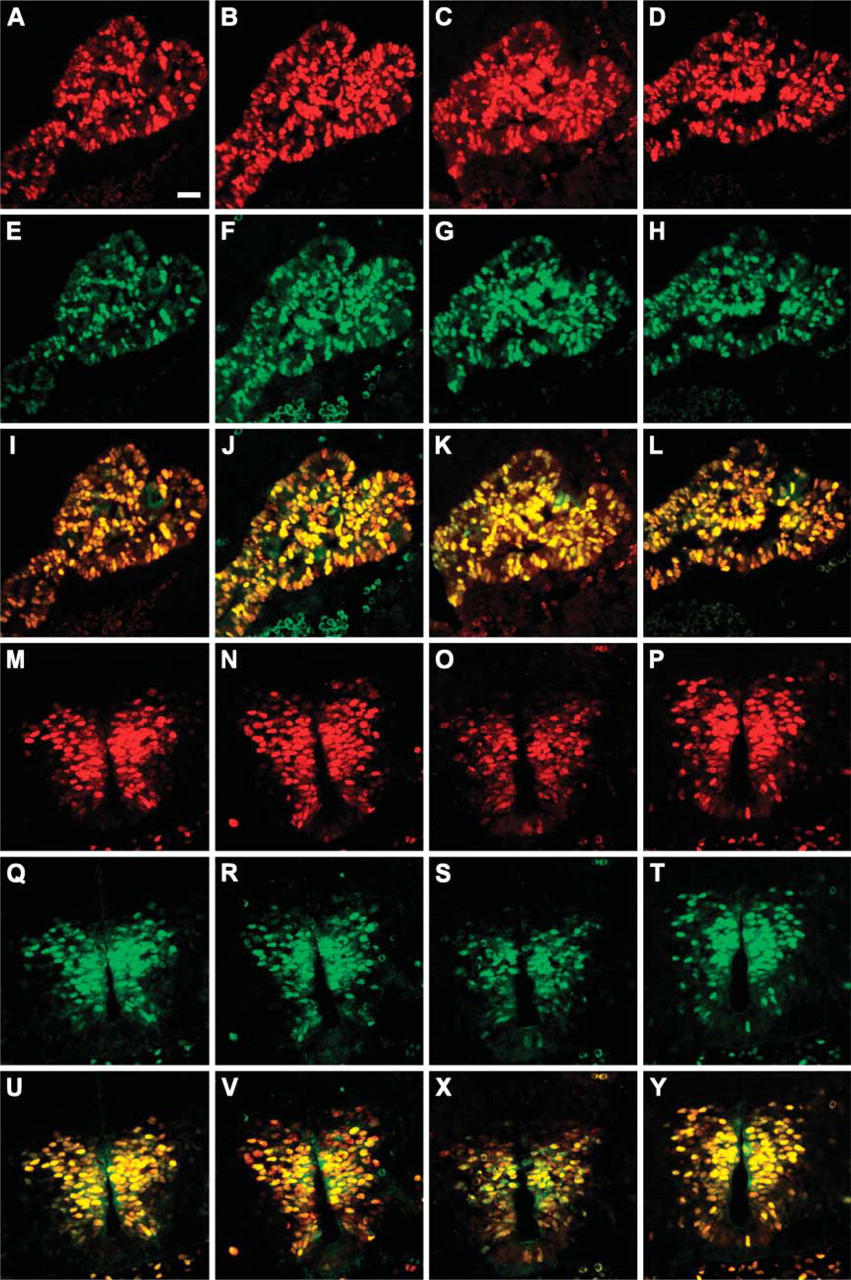
Mouse and rabbit anti-Nkx6.1 antibodies show matching staining patterns in embryonic tissues. Double immunohistochemical stainings of formalin-fixed frozen sections of an embryonic day (E11.5) mouse embryo with the polyclonal rabbit anti-Nkx6.1 antiserum (green) together with F55A10 (
SV40 large T-antigen transformed βTC-3 insulinoma cells were lysed in radioimmunoprecipitation assay lysis buffer (150 mM NaCl, 1% Nonidet P40, 0.5% natrium deoxycholate, 0.1% SDS, 50 mM Tris, pH 8.0, 1 mM 4-(2-amino-ethyl)benzenesulfonyl fluoride), and 30 μg whole-cell extract protein samples were denatured for 5 min at 80C in 5 μl NuPAGE 4x lithium dodecyl sulfate (LDS) sample buffer (Invitrogen) and 2 μl 1 M dithiothreitol. Samples were loaded on a NuPAGE 10% BIS-TRIS Gel (Invitrogen) and run alongside the Magic Mark XP Western standard (Invitrogen). This was followed by electroblotting at 400 mA for 75 min at 4C onto a nitrocellulose membrane in transfer buffer (20 mM Tris base, 150 mM glycine, 20% EtOH). The membrane was washed for 5 min in TBS (50 mM Tris, 150 mM NaCl), blocked for 1 hr in TBST (TBS + 0.1% Tween 20) with 2% skimmed milk powder, washed three times for 5 min each in TBST, and incubated O/N at 4C with monoclonal anti-Nkx6.1 antibody diluted 1:1000 in TBST 2% milk. After three 5-min washes in TBST, the blot was incubated for 30 min with goat anti-mouse HRP-conjugated secondary antibody (Santa Cruz Biotechnology; Santa Cruz, CA) diluted 1:10,000 in TBST 2% milk. Chemiluminescence was detected with Lumigen TSA-6 (Amersham Biosciences) using LAS 3000 from FUJIFILM (Tokyo, Japan) and viewed by Image Reader LAS 3000 software.
Results
Monoclonal antibodies against the Nkx6.1 homeodomain transcription factor were generated by immunizing RBF mice with a GST-Nkx6.1 fusion protein containing a C-terminal fragment of the Nkx6.1 transcription factor. This construct has previously been used for the generation of a polyclonal anti-Nkx6.1 rabbit antiserum (Jensen et al. 1996). All mice responded extremely well and developed a high titer immune response of <1:300,000 as tested by binding to the GST-Nkx6.1 fusion protein in ELISA. Because the aim was to generate antibodies for detection of Nkx6.1-expressing cells by immunohistochemical analysis, all sera were tested on sections of the adult mouse pancreas and shown to specifically stain the nuclei of a subpopulation of cells corresponding to the β-cell population. The mouse with the most prominent staining pattern was selected for fusion with FOX-NY myeloma cells to generate hybridomas producing monoclonal antibodies against Nkx6.1. The fusion was performed using a standard polyethylene glycol protocol, and the fused cells were seeded in 96-well plates. All clones were screened by ELISA for the differential reactivity against GST-Nkx6.1 and GST and resulted in 72 Nkx6.1-specific clones. In addition, the clones were screened against the corresponding GST-Nkx6.2 fusion protein, because the homeodomain transcription factor Nkx6.2 demonstrates a high degree of sequence similarity with Nkx6.1. Of the remaining clones, the 43 most-positive supernatants were further assayed for specific binding to PFA-fixed, frozen sections of adult mouse pancreas. Of these, 20 hybridomas demonstrated positive immunohistochemical stainings. The four clones (F55A10, F55A12, F64A6B4, and F65A2) were subjected to repeated subcloning and were established as stable hybridoma cell lines. These hybridomas, producing murine monoclonal antibodies against Nkx6.1 of the IgG1/κ isotype (data not shown), were grown in large scale and the antibodies purified from the culture supernatant using Protein A.
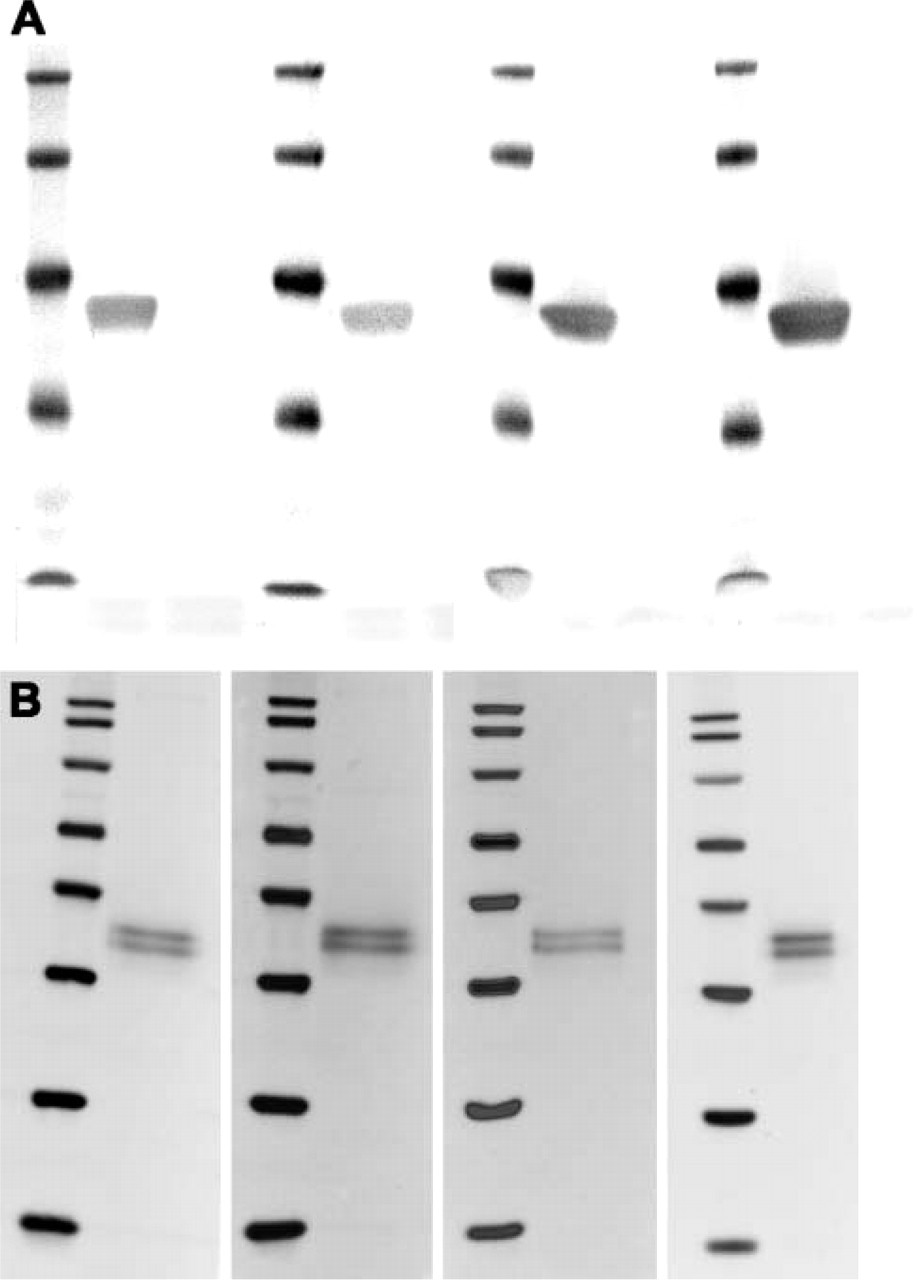
Specificity of Nkx6.1 monoclonals. (
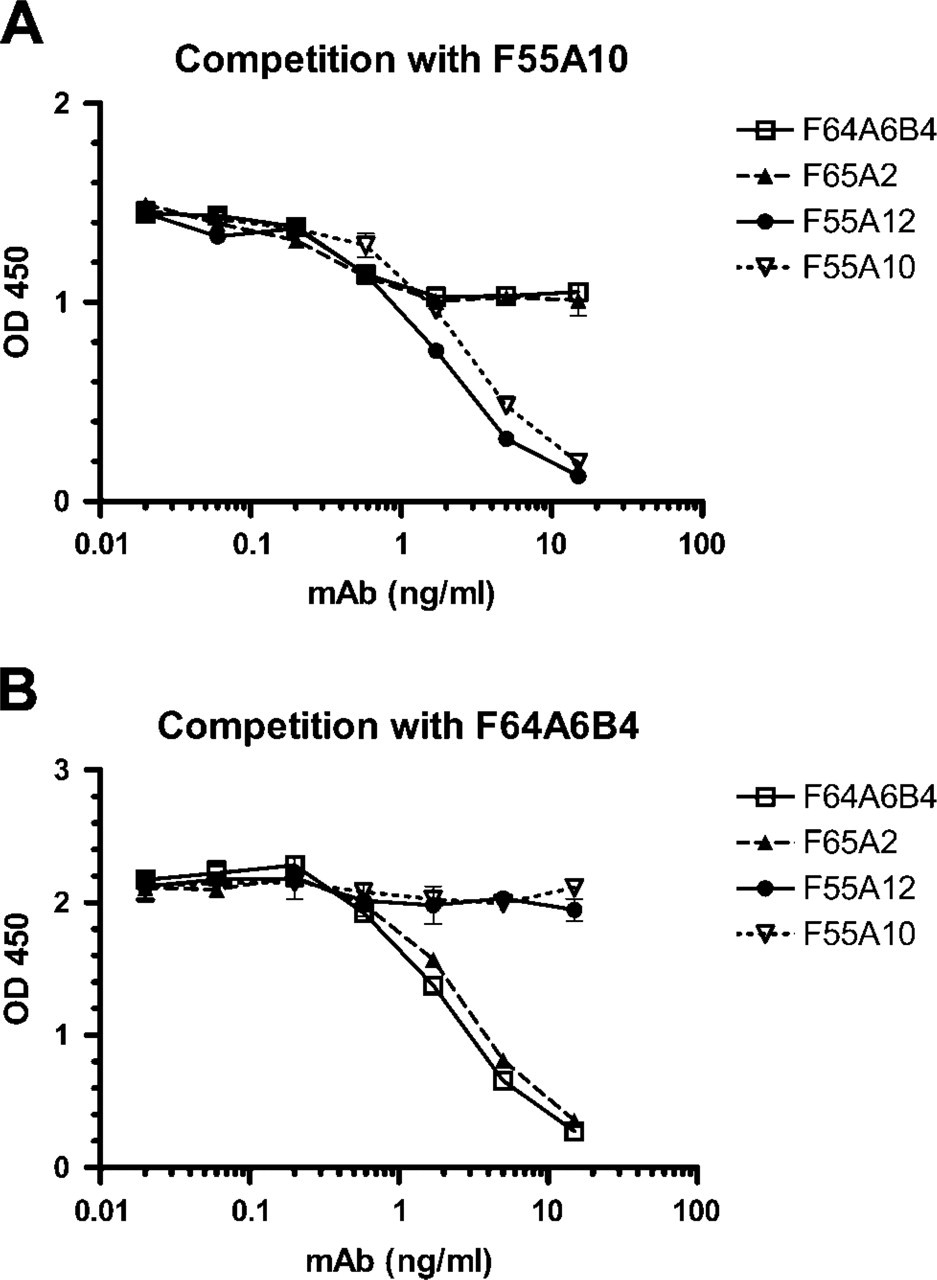
Antibody epitope mapping by competition ELISA. (
The four purified monoclonal anti-Nkx6.1 antibodies (F55A10, F55A12, F64A6B4, and F65A2) were characterized by staining of tissue sections of adult mouse pancreas. They all stain specifically the nuclei of a subpopulation of the cells in the endocrine islets of Langerhans (Figures 1A-1D). Specificity of the antibodies for Nkx6.1 was demonstrated by the selective absorption of antibody staining by GST-Nkx6.1 (Figures 1E-1H) but not by a GST-NGN3 fusion protein (Figures 1I-1L). When costaining for the peptide hormones insulin and glucagon produced by the β-cells and the α-cells, respectively, it is clear that the anti-Nkx6.1 antibodies localize to the nuclei of the insulin-producing β-cells and not to the glucagon-producing α-cells of mature islets of Langerhans in adult mouse pancreas (Figures 1M-1P). Furthermore, the staining patterns of all four antibodies are identical to that of the original polyclonal rabbit serum in both adult pancreatic islets (Figure 2) as well as in embryonic pancreatic epithelium and ventral neural tube (Figure 3). In addition, all four monoclonal antibodies demonstrated staining of the expected 36-kDa band in Western blots against purified GST-Nkx6.1 protein, whereas the antibodies did not react against the related GST-Nkx6.2 protein (Figure 4A). Also, they all stain the expected Nkx6.1 double band of 44 and 46 kDa in protein extracts from the SV40 large T-antigen transformed insulinoma βTC-3 cell line (Figure 4B) (Jensen et al. 1996).
Binding of the antibodies was mapped to two different epitopes of the GST-Nkx6.1 protein by competition ELISA. This experiment demonstrated that the two antibodies (F55A10 and F55A12) were able to compete with each other, but not with the antibodies F64A6B4 and F65A2, and vice versa (Figure 5). The F55A10 antibody has additionally been shown to recognize Nkx6.1 from chicken, mice, pigs, and humans (data not shown) to recognize Nkx6.1 in formalin-fixed, paraffin-embedded tissue after antigen retrieval (data not shown) and to work in standard indirect immunofluorescence experiments (i.e., without tyramide signal amplification) at a dilution of 1:200.
Discussion
This panel of new mouse monoclonal antibodies against the homeodomain transcription factor Nkx6.1 will become an important tool for investigations toward understanding the mechanisms of pancreatic development at the earliest stages of gut tube formation as well as the differentiation process of pancreatic endocrine β-cells from precursor cells. They will allow for the further characterization of the role and function of Nkx6.1 by the simultaneous detection of Nkx6.1 and a large number of other transcription factors involved in the development of pancreas for which only polyclonal rabbit antisera are available.
Footnotes
Acknowledgments
This work was supported by the National Institute of Diabetes & Digestive & Kidney Diseases (NIDDK; 1U19 DK-61244-4 and 5U19 DK-42502-15) as part of the Beta Cell Biology Consortium. M.C.J was supported by a fellowship from the Juvenile Diabetes Research Foundation. O.D.M. and P.S. were supported by the EU 6th Framework Programme.
We thank Susan Brenner-Morton for providing the GST-Nkx6.1 expression plasmid and Anette Bjerregaard, Evelyn Kury, Anders Moselund, Hanne Duus, and Camilla Schwartz for excellent technical assistance.