
Abstract
Tumorigenesis is associated with the activation of mitogenic signal transduction pathways. The expression of activated extracellular signal-regulated kinase (p-ERK) may play an important role in cell proliferation of malignant mesothelioma (MM). We compare the expression of p-ERK in 50 biopsy specimens of MM, non-small-cell lung cancer (NSCLC), and normal lung tissue. We hypothesized that phosphorylated extracellular signal-regulated kinase is increased in MM. We stained the sections by immunohistochemistry for activated ERK-1 and −2 and performed the quantification of the stained nuclei. Quantitative analysis of p-ERK showed a high percentage score in MM (30.3 ± 4.6%) as compared with NSCLC (12.2 ± 2.1%) (p<0.01) and control lung tissue (6.4 ± 1.3%) (p=0.0002). Furthermore, p-ERK was found significantly higher in poorly differentiated NSCLC (17.7 ± 3.1%) as compared with well-differentiated NSCLC (5.4 ± 1.2%) (p<0.01). Our data show that the nuclear quantification of p-ERK is significantly increased in MM and poorly differentiated NSCLC in comparison to well-differentiated NSCLC and normal lung tissue. These results corroborate previous experimental studies that suggest a critical role of p-ERK in cell proliferation of malignant disease and may represent new targets for therapeutic agents.
E
In the last two decades, the worldwide incidence of human malignant pleural mesothelioma (MM) has increased dramatically (Rizzo et al. 2001). Recent epidemiological data show that, in the U.S., MM accounts for 2000 deaths per year among the reported 130,000 deaths from lung cancer (Rizzo et al. 2001; Mossman and Gruenert 2002). Epidermal growth factor and platelet-derived growth factor (PDGF) and their receptors are implicated in the process of pathogenesis in these tumors (Viallet and Sausville 1996; Cacciotti et al. 2002). Asbestos exposure causes a specific EGF-r-dependent activation of ERK-1 and −2 in rat pleural mesothelial cells (Zanella et al. 1996). The nuclear localization of phosphorylated p-ERK (p-ERK-1 and −2) varies among different tumors in relation to both grade and/or stage (Mandell et al. 1998; Gioeli et al. 1999; Albanell et al. 2001). Few studies have compared the morphological expression of these activated kinases in MM (Mossman et al. 1990) and non-small-cell lung carcinoma (NSCLC) (Mukohara et al. 2003; Vicent et al. 2004). However, to date no study has assessed the p-ERK immunostain in MM and NSCLC. In this study we aim to analyze the expression of p-ERK-1 and −2 by immunohistochemistry with a phosphospecific antibody in lung tissue sections of MM, NSCLC, and control normal lung tissue. We hypothesized that in human samples an activation of p-ERK could be documented.
To document the relative importance of another pathway recently described in MM (Altomare et al. 2005), we also investigated the expression of p-phosphatidylinositol 3-kinase (PI3K/AKT). A better understanding of MM and NSCLC biology could lead to new treatments of these diseases.
Materials and Methods
Biopsies
A total of 50 specimens, diagnosed between 1996 and 2002, were analyzed in this study: 31 NSCLC (16 squamous cell carcinomas and 15 adenocarcinomas), 10 MM (6 epithelioid, 1 sarcomatous, and 3 biphasic phenotypes) and 9 non-neoplastic lung tissues. With regard to MM, asbestos exposure was known in seven of the cases, two had no history of asbestos exposure, and there were no data available for the last case. The material was fixed in buffered formalin and embedded in paraffin wax. For all the lung controls, NSCLC, and two MM, the sections came from the material of pneumectomy or pleuropneumectomy. For the eight MM, the sections came from the biopsies obtained by thoracoscopy.
Histological diagnosis of lung tumors was established according to the World Health Organization criteria (Travis et al. 2004). In addition, they were graded as well differentiated or poorly differentiated. For the purpose of the present analysis, moderately differentiated tumors were included in the poorly differentiated group. All sections were stained with immunohistochemistry. p-ERK stains were reviewed and diagnosed by two independent examiners.
Immunohistochemistry
All antibodies were purchased from Cell Signaling Laboratories Technology, Inc. (Beverly, MA). Immunostaining was performed on 4-μm tissue sections, placed on slides coated with 3-(triethoxysilyl)propylamine (Merck; Darmstadt, Germany), dewaxed in xylene, and dehydrated in alcohol. For heat-induced epitope retrieval of the p-ERK stains, the specimens were washed three times in citrate buffer, pH 6.0, for 5 min in a microwave at 650W. Sections were washed with Tris-buffered saline, pH 7.6, and then rinsed in 0.1% Triton X-100/TBS for 5 min at room temperature (RT). Samples were preincubated with blocking solution (normal swine serum 10%, BSA 3% in 0.1% Triton X-100/TBS) for 30min at RT. Immunostaining was performed with two rabbit polyclonal antibodies that detect the phosphorylated forms of the MAP kinase ERK-1 and −2 (p-ERK) and the total ERK protein levels (i.e., both phosphorylated and non-phosphorylated forms). After washing, the sections were incubated for 1 hr in a humid chamber at RT with the first antibody at a dilution of 1/80 for p-ERK and 1/20 for ERK. After subsequent washing in 0.1% Triton X-100/TBS, samples were incubated with a biotinylated swine anti-rabbit immunoglobulin (DAKO A/S; Glostrup, Denmark) at a dilution of 1/250 for 30 min at RT. They were then rinsed in 0.1% Triton X-100/TBS before incubation in StreptAB Complex/AP (DAKO) at a 1/100 dilution for 30 min at RT. Specimens were washed and developed with Fast Red Substrate solution (DAKO) for p-ERK for 15 to 30 min at RT, rinsed in H2O, and then counterstained with hematoxylin. They were then mounted in aqueous mounting media (Aquatex; Merck). p-AKT was stained with a rabbit monoclonal antibody. The sections were treated with heat-induced epitope retrieval in a pressure cooker with a buffer citrate at pH 6.0 for 3 min. They were incubated with p-AKT at a dilution 1/20 for 60 min. After subsequent washing, they were incubated with the rabbit Envision/ HRP (DAKO) and then developed with diaminobenzidine. The slides were dehydrated and mounted in Eukitt. Non-neoplastic lung tissues from nine different subjects were used as internal control. Negative controls for p-ERK consisted of sections stained with an isotype-matched non-immune antibody at the same protein concentration (DAKO A/S), and normal skin was used as a positive control. Polymorphonuclear neutrophils, fibroblasts, and endothelial cells usually showed some degree of immunoreactivity and were considered as internal positive controls. Lymphocytes stained negative and were used as an internal negative control. In addition, p-ERK and p-AKT were preabsorbed with their respective blocking peptide.
Light Microscope Analysis
For p-ERK immunostaining, two independent observers scored the sections by counting 6 to 10 fields (X400) with nuclear stain assessed in the non-tumoral and tumoral tissues. Stained nuclei were considered as positive independently of the intensity of the staining. The counts were done in the fields with the more numerous stained nuclei. These were mostly located at the border of the tumor for NSCLC. All cells were counted in the fields.
Cytoplasmic and/or nuclear localization of p-AKT was reported as positive.
Statistical Analysis
Comparative nuclear expression of p-ERK between the studied groups was estimated using the unpaired two-tailed t-test. Data are shown as box plots representing median and lower and higher values, as well as first and third quartiles. Analysis was performed using the software GraphPrism version 3.00 for Windows (GraphPad Software; San Diego, CA). A p value ≤0.05 was considered statistically significant.
Results
Figure 1 shows that 30.3 ± 4.6% of nuclei were stained in MM as compared with 17.7 ± 3.1% in poorly differentiated NSCLC (p<0.05), 5.4 ± 1.2% in well-differentiated NSCLC (p<0.0001), and 6.4 ± 1.3% in non-neoplastic tissue sections (p<0.0001). Furthermore, expression of p-ERK was significantly increased in poorly differentiated NSCLC as compared with well-differentiated NSCLC (p<0.01) and normal lung tissue (p<0.01). Conversely, the percentage of nuclei stained in well-differentiated NSCLC was not statistically different from control lung tissue sections.
The expression of p-ERK in two types of NSCLC (namely, adenocarcinoma and squamous cell carcinoma) is detailed in Figure 2. Poorly differentiated tumors showed a higher percentage of nuclei stained with p-ERK than well-differentiated tumors in both types (p<0.05), with a more scattered distribution of p-ERK expression in the former. Variation in the levels of activation was observed in three cases of adenocarcinoma. One poorly differentiated case showed 4% staining, whereas two others with a well-differentiated grade resulted in positive staining values of 12% and 16%, respectively. This heterogeneity could reflect transition changes involving the overexpression of other elements in the transduction cascades during carcinogenesis.
Immunohistochemical Localization of p-ERK in MM and NSCLC
The pattern of MM staining, including epithelioid, sarcomatous, and biphasic phenotypes, showed intense, homogeneous staining both in the cytoplasm and in the nucleus, reflecting high levels of p-ERK activation in these cells (Figures 3E and 3F). The qualitative analysis of the NSCLC sections revealed a very heterogeneous pattern of MAP kinase staining in both the cytoplasm and nucleus of the primary tumor, varying from weak, to moderate, to intense (Figures 3G-3J). Frequently, positive clusters of cells localized at the periphery of tumors in the region of the edge of growing zones, whereas the centers of the tumor region were completely negative (Figure 3G). However, in all NSCLC we occasionally found a few positive staining cells in the central regions of the tumor. This cellular staining was extended to both groups of NSCLC (data not shown). Four negative cases among the NSCLC were detected. Non-tumoral cells around the tumor stained more frequently than others located more distantly, possibly as a result of paracrine stimulus (Figure 3D).
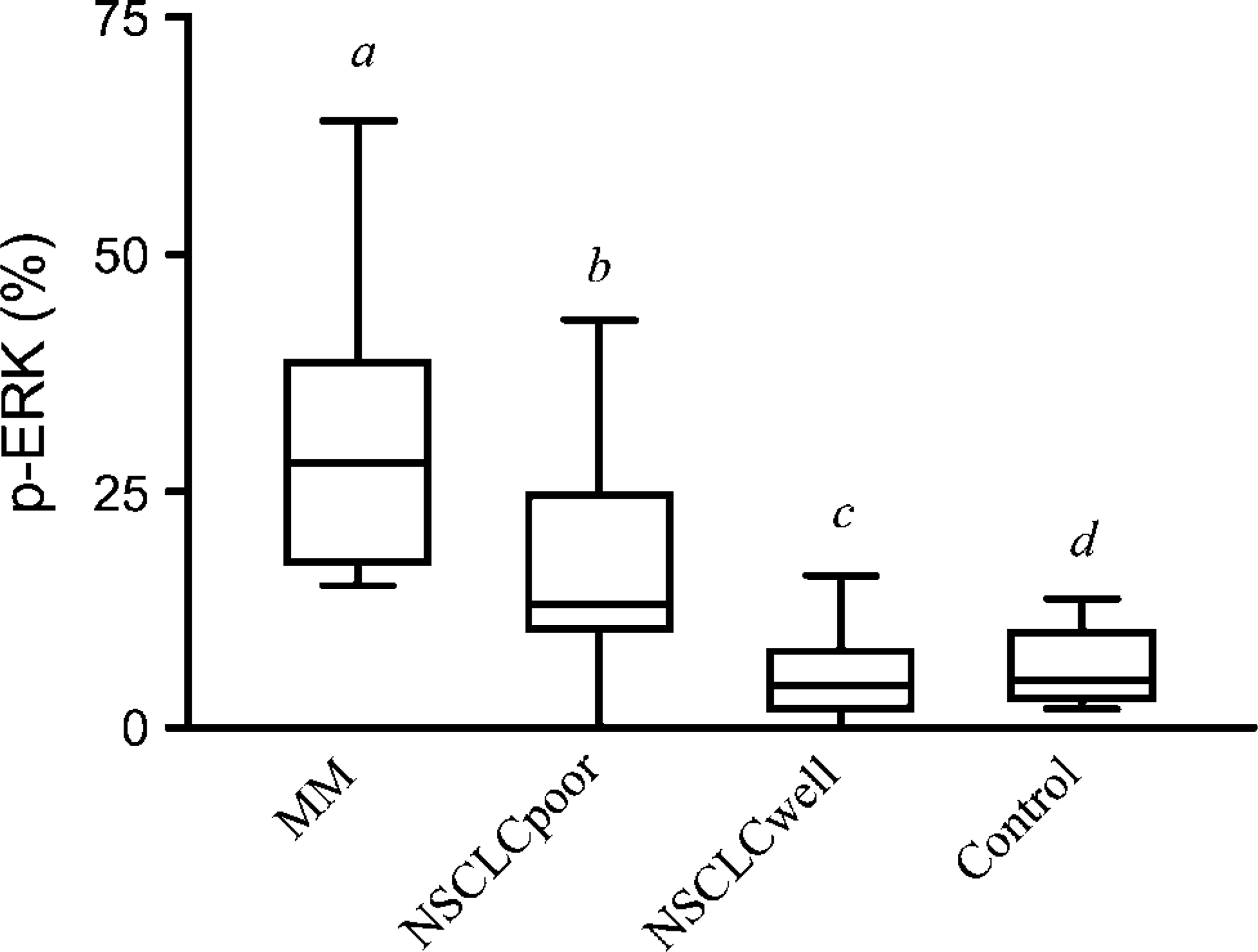
Expression of activated p-ERK nuclear staining (%) in malignant mesothelioma (MM), poorly differentiated non-small-cell lung carcinoma (NSCLCpoor), well-differentiated non-small-cell lung carcinoma (NSCLCwell), and non-tumoral lung tissue (control). Box plots represent median, first quartile (Q1), third quartile (Q3), lower value, and higher value. Statistical significance between different bars: a vs. b, <0.05; a vs. d, <0.0001; b vs. c and b vs. d, <0.01; c vs. d, non-significant.
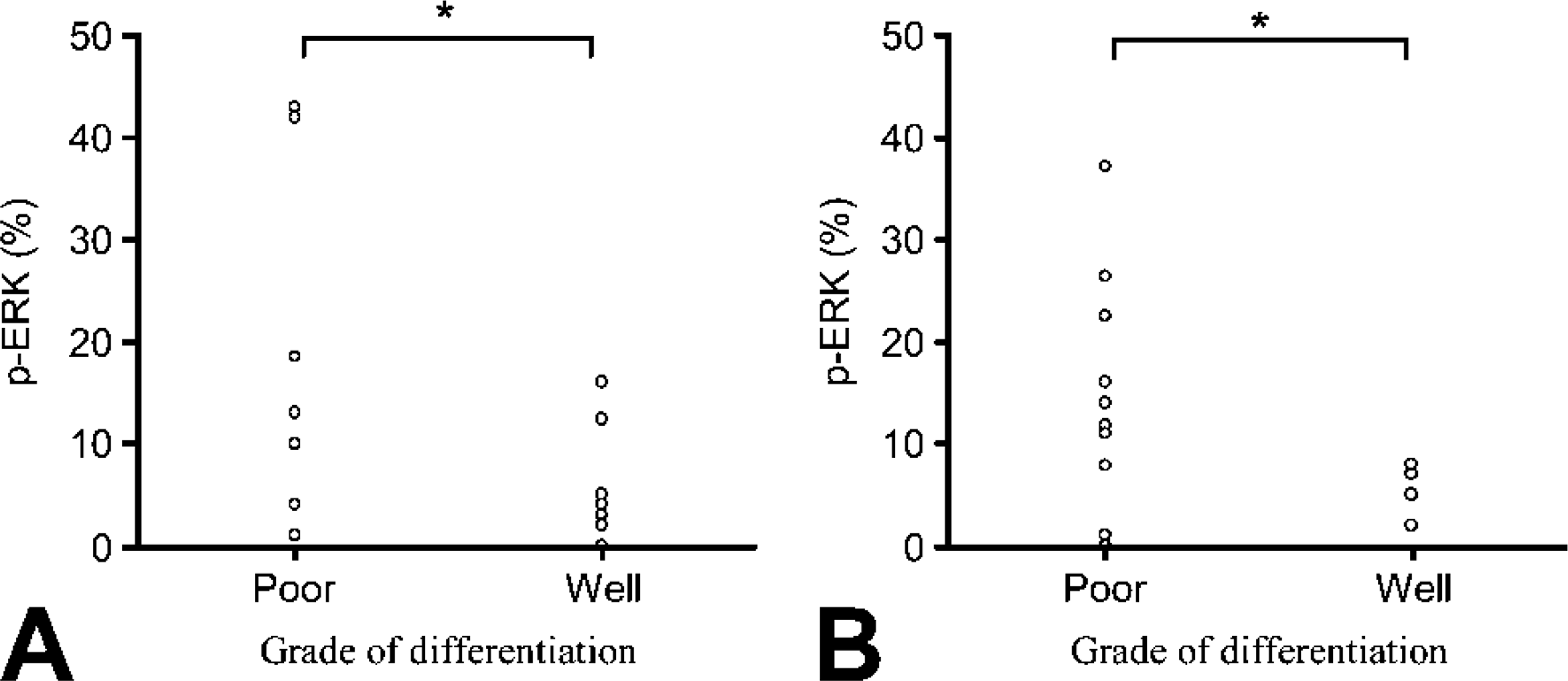
Expression of activated p-ERK nuclear staining (%) in well-and poorly differentiated non-small-cell lung carcinoma. (
In the non-neoplastic sections, p-ERK detection located in rare pneumocytes, fibroblasts, and endothelial cells (Figure 3B). The total ERK antibody showed a uniform cytoplasmic staining distribution. Nuclear positivity with this antibody distributed heterogeneously in relation to tissue distribution (Figure 3C).
After incubation of p-ERK with the blocking peptide, no immunostain was observed (Figure 4B).
Immunohistochemical Localization of p-AKT in MM and NSCLC
p-AKT colored all mesotheliomas. The staining pattern was either nuclear or cytoplasmic (Figure 4C). For NSCLC, the four squamous cell carcinomas were tested and stained in the nucleus and/or in the cytoplasm. For the five adenocarcinomas tested, three were negative and two stained in the nucleus and/or in the cytoplasm.
After incubation of p-AKT with the blocking peptide, no immunostaining was observed (Figure 4D).
Discussion
The results presented in this study demonstrate that the nuclear quantification of p-ERK is significantly increased in MM and in poorly differentiated NSCLC in comparison to control lung. In addition, we show that p-ERK is significantly increased in poorly differentiated NSCLC as opposed to well-differentiated NSCLC. p-AKT stain was observed in all MM and some NSCLC.
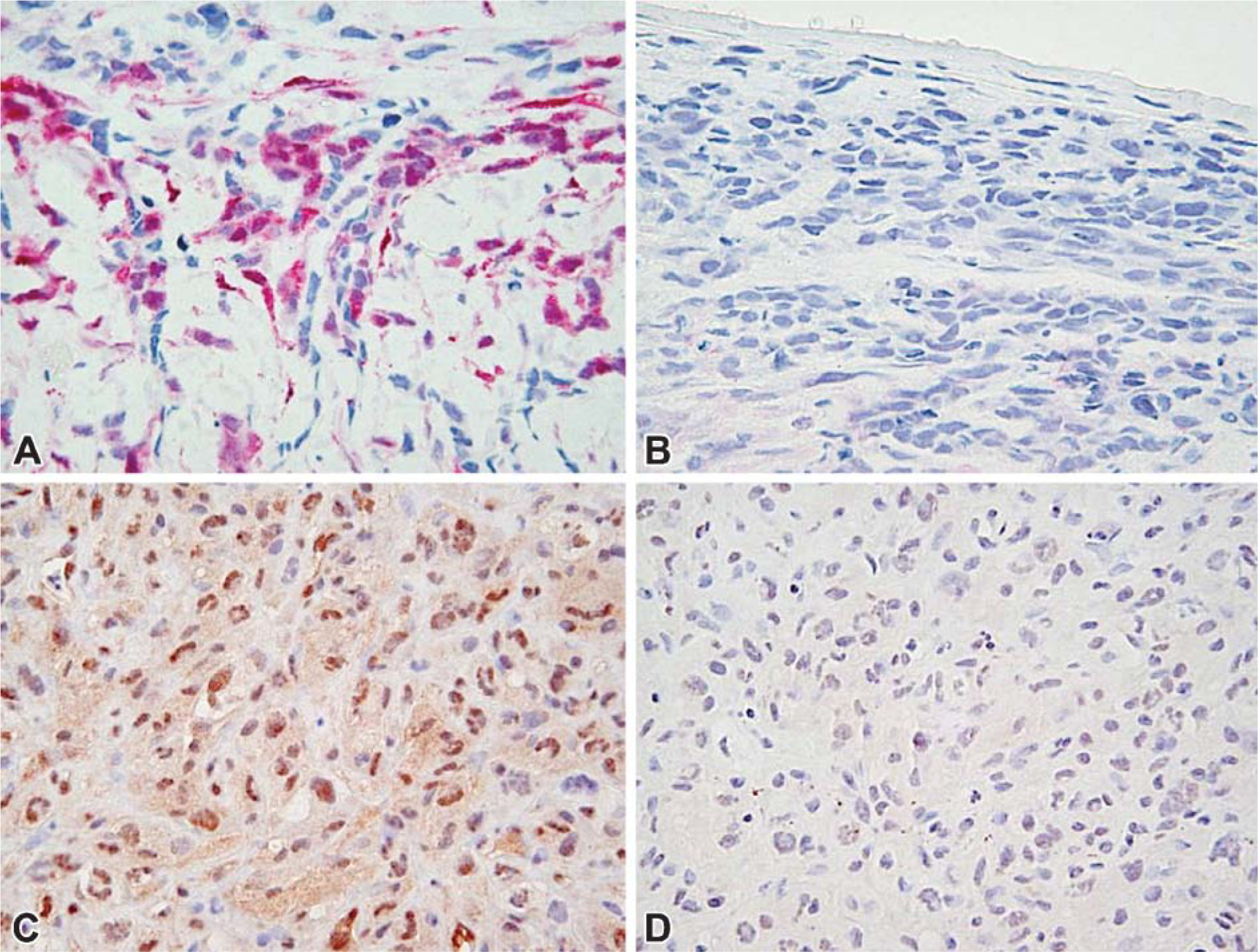
Immunohistochemistry of activated p-ERK in MM (
The use of highly specific antibodies that recognize activated signaling molecules within the cellular context allows the analysis of potential relationships between intracellular phosphorylation mechanisms and tumorigenesis. With this approach, one examines in situ the evidence for aberrant signals, as manifested by a change in the levels of activated proteins and/or their cellular localization. Furthermore, we can observe perturbations in a particular intracellular pathway that may lead to the promotion of abnormal cell proliferation.
Few studies investigated the role of ERK in MM and NSCLC. Ramos-Nino and coworkers (2002) reported that inhibition of ERK activation abrogated mesothelioma cell growth in soft agar and restored contact inhibition. Mukohara et al. (2003) found that, in NSCLC, the level of EGF-r expression was associated with the expression of p-ERK, but staining for p-ERK was not significantly correlated to any clinicopathological factor. In our study, p-ERK was significantly increased for poorly differentiated NSCLC. However, p-ERK expression was significantly decreased in the remaining well-differentiated cases in relation to the totality of NSCLC cases. Earlier work (Mukohara et al. 2003) employed a quantification system based on the intensity of the nuclear staining, an approach somewhat different from that adopted in this study.
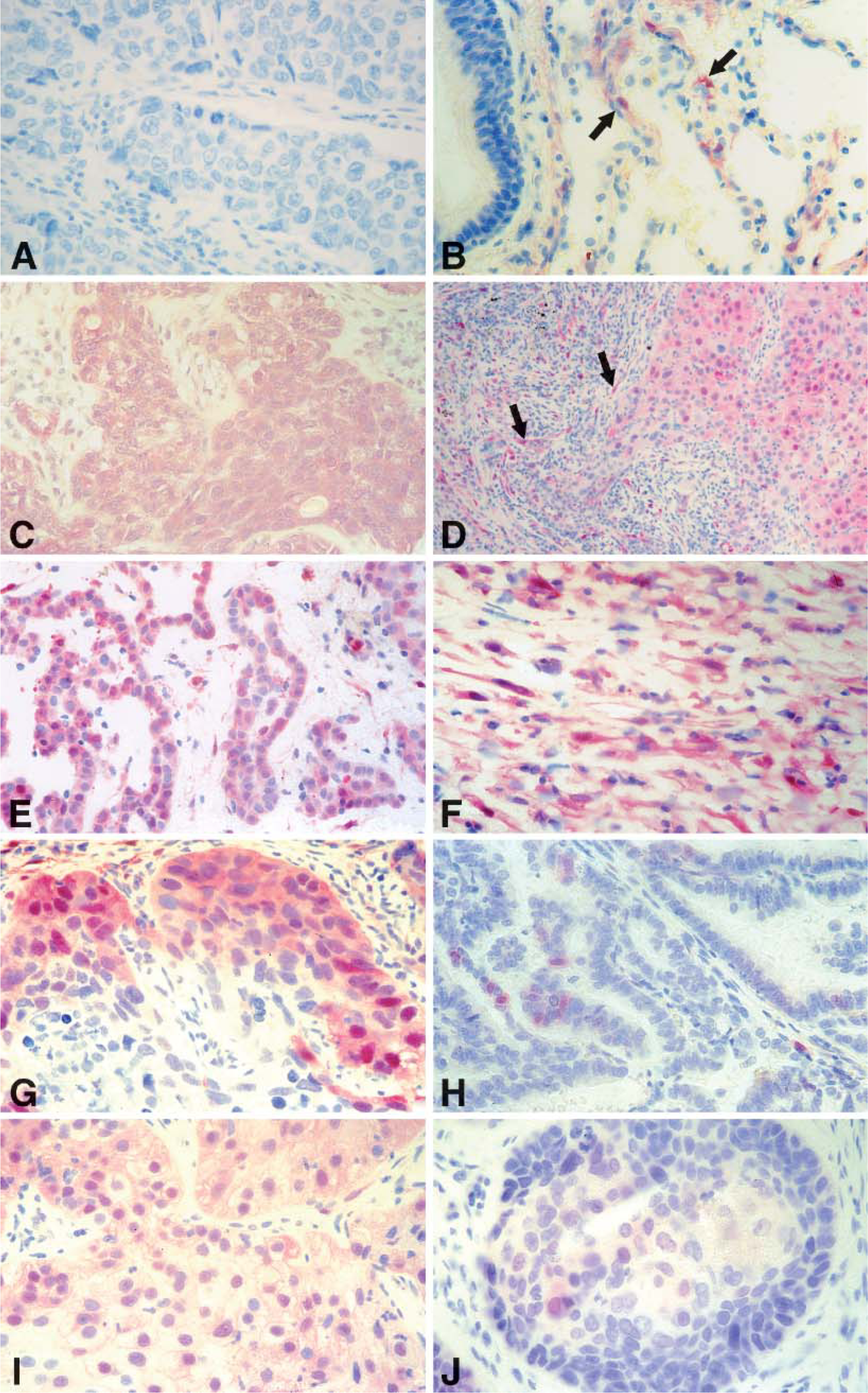
Immunohistochemistry of activated p-ERK in human tissues. (
In a recent report, Vicent and collaborators (2004) observed a strong correlation between nuclear and cytoplasmic p-ERK staining and advanced stages of NSCLC. Our results add to this report. However, methodological differences limit the comparison between studies. The quantification criterion used in their earlier report was different from that employed in our study, in that a cutoff of 15% was chosen for considering a positive p-ERK. Using this approach, all of our well-differentiated carcinoma cases would have been negative for p-ERK with the exception of one adenocarcinoma. Indeed, this could explain the absence of correlation between the nuclear p-ERK expression and the degree of tumor differentiation as reported by authors in the previous study. On the other hand, whereas moderately differentiated carcinoma cases were included in the well-differentiated group by Vicent and co-workers, in our study they were placed in the poorly differentiated NSCLC group.
Our quantitative data show a statistically significant increase in the nuclear expression of p-ERK in MM and in poorly differentiated NSCLC compared with controls. This corroborates previous results by Albanell et al. (2001) who reported similar percentages of cells stained for p-ERK expression in head and neck squamous cell carcinoma. This microscopic approach allows a spatial localization of p-ERK in the tumor tissue as well as in the subcellular compartments. High levels of nuclear p-ERK were equally found at the frontal growth area and at the peripheral region of the tumor in both categories of NSCLC. Others studies have shown the presence of p-ERK preferentially in the invading front of the tumor (Albanell et al. 2001). The fact that activated p-ERK was also detected in normal cells around tumor regions may be explained by the induction of growth factors such as TGF-α and PDGF, which stimulate the proliferation of the tumor in an autocrine fashion and also the surrounding normal cells in a paracrine fashion (Hoshino et al. 1999). Moreover, our study indicates that poorly differentiated NSCLC showed a significantly increased nuclear expression of p-ERK in comparison to well-differentiated NSCLC. In the two categories of NSCLC used in this study, namely, squamous cell carcinoma and adenocarcinoma, the quantitative analysis of the nuclear localization of p-ERK revealed a marked increase in poorly differentiated grades. In addition, MM showed a significantly increased nuclear expression of p-ERK in comparison to poorly differentiated NSCLC. These findings suggest that high levels of MAPK activation occur in MM and in poorly differentiated NSCLC. Additional signaling pathways are also activated in MM and NSCLC as shown by the p-AKT immunostains.
In conclusion, data presented in this study suggest that phosphorylation of ERK may constitute a relevant process in regulating cell proliferation in MM and in poorly differentiated NSCLC. As a consequence, blocking the MAPK pathway by specific inhibition of the EGF-r could have a determinant role in lung cancer treatment (Averbuch 2002; Lonardo et al. 2002). Jänne et al. (2002) reported that in MM cell lines small moecular inhibitors significantly perturbed signaling that depends on activation of EGF-r, including phosphorylation of ERK-1 and −2. Such an approach may have therapeutic applications to the clinical setting. However, to what extent it represents a real target for future drug therapy remains to be demonstrated.
Footnotes
Acknowledgements
The authors thank Professor Anastase Spiliopoulos for his support and Professor Brooke Mossman for insightful discussion of the results. The authors acknowledge the helpful contributions of the Immunopathology Laboratory staff, as well as the editorial support of Alberte Polichouk, Jean-Claude Rumbeli, and Etienne Denkinger.