
Abstract
Efforts to improve severely impaired myocardial function include transplantation of autologous hematopoietic side population (SP) stem cells. The transmembrane ABC-type (ATP binding cassette) half-transporter ABCG2 (BCRP) serves as a marker protein for SP cell selection. We have recently shown that other ABC transport proteins such as ABCB1 and ABCC5 are differentially expressed in normal and diseased human heart. Here we investigated localization and individual ABCG2 expression in 15 ventricular (including 10 cardiomyopathic) and 51 auricular heart tissue samples using immunohistochemistry, confocal laser scanning fluorescence microscopy, and real-time RT-PCR. Individual genotypes were assigned using PCR–restriction fragment length polymorphism (RFLP) analysis and subsequently correlated to ABCG2 mRNA levels. ABCG2 was localized in endothelial cells of capillaries and arterioles of all samples. Ventricular samples from cardiomyopathic hearts exhibited significantly increased levels of ABCG2 mRNA (ABCG2/18S rRNA: 1.08 ± 0.30 × 10−7; p = 0.028 (dilative cardiomyopathy) and 1.16 ± 0.46 × 10−7; p = 0.009 (ischemic cardiomyopathy) compared with 0.44 ± 0.26 × 10−7 in nonfailing hearts). The individual haplotypes were not associated with altered mRNA expression. ABCG2 is variably expressed in endothelial cells of human heart, where it may function as a protective barrier against cardiotoxic drugs such as anthracyclines or mitoxantrone. ABCG2 expression is induced in dilative and ischemic cardiomyopathies.
C
ABCG2 belongs to the G-branch of the superfamily of ABC transporters. After binding and hydrolyzing ATP, these proteins are capable to transport a variety of xenobiotics and endogenous compounds across cellular membranes, thereby providing both protection against toxins and contributing to cellular signaling. Located at chromosome 4q22, the ABCG2 gene encodes a 655 amino acid polypeptide, which is classified as a half-transporter consisting of six putative transmembrane domains and one nucleotide binding domain. Recently, it has been reported that ABCG2 forms homotetramers to create the functional active protein (Xu et al. 2004). Besides the expression of ABCG2 in human cell lines from breast, colon, ovary, and gastric cancer, the distribution of ABCG2 has been located in human colon, small intestine, hepatic bile canaliculi, mammary gland, and the endothelia of veins and capillaries of heart (Maliepaard et al. 2001) and brain (Zhang et al. 2003). Aside from its expression in HSC and subsequent use as selection marker, ABCG2 has been found recently in mice cardiac progenitor cells, which exhibit stem cell characteristics in the adult heart and are distinct from endothelial cells (Martin et al. 2004).
We have recently described expression and function of other ABC transporters in human heart, namely ABCB1 (P-gp) and ABCC5 (MRP5). Although ABCB1 is thought to be involved in cellular export of drugs and toxins (Meissner et al. 2002, 2004), ABCC5 alters intracellular levels of cGMP (Dazert et al. 2003). In contrast, expression of ABCG2 in human heart has not been studied in detail. In view of the ongoing discussion of ABCG2 as a marker for stem cells, characterization of constitutive expression in human heart is of interest. In the present study, therefore, we assessed expression and localization of ABCG2 in human heart as functions of disease processes and genetic polymorphisms. We describe variable expression of ABCG2 in the endothelial wall of cardiac arterioles and capillaries and a significant increase of ABCG2 in patients with cardiomyopathies.
Materials and Methods
Tissue Samples
After approval by the local ethics committee and obtained written informed consent, auricular heart tissue samples were taken from 51 patients undergoing coronary artery bypass graft surgery (Caucasians, 39 male, 12 female, 47–80 years of age). The investigation is in line with the principles outlined in the Declaration of Helsinki. Medical therapy of patients consisted of cardiac glycosides, beta blockers, statins, diuretics, nitrates, and angiotensin-converting enzyme inhibitors. Ventricular samples were taken from excised heart left ventricle during orthotopic heart transplantation as described before (Meissner et al. 2002). In brief, samples from five nonfailing hearts were taken from the excised myocardium of potential donors without medical history or diagnostic evidence of heart disease, whereas the valves were taken and used for human homografts. Samples from patients with ischemic or dilative cardiomyopathy (n = 5, each) were taken from hearts excised because of heart failure. Tissue samples were snap-frozen in liquid nitrogen or fixed in 4% paraformaldehyde. The frozen tissue was homogenized using a vibration grinding mill for RNA isolation (Mikro-Dismembrator; Braun, Germany).
RT-PCR of ABCG2 mRNA
200 ng of total RNA, prepared from 50 mg frozen tissue homogenate (RNEasy mini extraction kit; Qiagen, Hilden, Germany), were reversely transcribed using random hexamers and the TaqMan reverse transcription reagents (Applied Biosystems; Weiterstadt, Germany). PCR of ABCG2 and 18S rRNA was performed using the intron-spanning primers hABCG2-F TATTACCCAT GACGATGTTA CCAAGTA TTA TATTTACCTG TA, hABCG2-R ATGGTGGCTT ATTCAGCCAG TTCCATGG, and the FAM-marked probe hABCG2-P TTCATGTTAG GATTGAAGCC AAAGGCAGAT as well as the predeveloped rRNA control reagents (Applied Biosystems) for 18S rRNA and the universal PCR mastermix (Applied Biosystems). PCR products were amplified (50C, 2 min; 95C, 10 min; followed by 40 cycles of 95C, 15 sec, and 60C, 1 min) and analyzed on a real-time PCR cycler (ABI Prism 7700; Applied Biosystems). Fluorescence threshold cycles (CT) of each sample were compared with the CT values of the standardization series, which consisted of the cloned ABCG2 PCR-fragment in pGem-Teasy (Promega; Mannheim, Germany) resulting in a quantification of mRNA copy numbers. ABCG2 mRNA expression levels were normalized with respect to the expression levels of 18S rRNA.
ABCG2 Genotype
For studying the influence of single nucleotide polymorphisms (SNPs) on the amount of ABCG2-mRNA in human heart, the 51 auricular heart samples were genotyped for previously described hereditary variants. Genomic DNA (gDNA) was extracted using Qiagen Blood Kit as described by the manufacturer (Qiagen; Frankfurt, Germany). 100 ng of the gDNA were amplified using primers flanking the regions of previously described SNPs. Primers BCRPEx2f GATAAAAACTCTCCAGATGTCTTGC and BCRPEx2r AGCCAAAACCTGTGAGGTTCAC were used for amplification of a 287-bp fragment containing 34G>A in exon 2 and BCRPEx5f TGTTGTGATGGGCACTCTGATG and BCRPEx5r ATCAGAGTCATTTTATCCACAC for a 222-bp fragment containing the 421C>A variant in exon 5. PCR was carried out in a volume of 25 μl containing 200 nM of each primer 10 X PCR buffer, 2 mM MgCl2, 0,8 mM of dNTP-Mix, and 0.5 U TaqPolymerase. PCR products were restricted using sequence-depending endonucleases for 34G>A (BsrI) and 421C>A (HpyCH4 III) and visualized after electrophoretic separation in a 2% agarose gel and staining by ethidium bromide.
Immunohistochemistry and Immunofluorescence
Tissue samples were fixed in formalin and embedded in paraffin. After cutting, sections were mounted on slides and dried overnight at 60C. For immunohistochemistry, the sections were deparaffinized using xylene. After rehydratation in decreasing ethanol solutions, the slides were boiled in 10 mM citrate buffer (pH 6.0). The endogenous peroxidase was blocked with 0.3% H2O2, and the biotin-streptavidin immunoperoxidase method was employed using the LSAB HRP detection system (DAKO; Hamburg, Germany) and the monoclonal anti-BCRP antibody BXP-21 (Alexis Biochemicals; Grünberg, Germany) at a dilution of 1:40. For immunofluorescence, the slides were blocked with 5% fetal calf serum (FCS) in PBS after rinsing with PBS (pH 7.4). The slides were then incubated with the BXP-21 antibody (Alexis) at a dilution of 1:50 overnight in a humidified atmosphere. For double staining of endothelial structures, an anti-PECAM antibody was used at a dilution of 1:100 (anti-PECAM; Sigma, Munich, Germany). After several rinses with PBS, the slides were blocked again with 5% FCS in PBS and then incubated with the respective secondary antibodies conjugated to Alexa Fluor 488 or Alexa Fluor 568 (Molecular Probes; Eugene, OR). For staining of the nuclei, DNA was counterstained with TOTO-3 iodide (Molecular Probes) and was added to the mounting medium in a dilution of 1:2000. After several washing steps with PBS, the slides were mounted with anti-fading mounting medium. Fluorescence micrographs were taken with a confocal laser scanning microscope (Chromaphor Analysen-Technik; Duisburg, Germany). Samples were observed with a Nikon inverted microscope and a 100 X oil-immersion objective. A CCD camera and VoxCell scan software from VisiTech International (Sunderland, United Kingdom) were used for analysis.
Immunoblot Analysis
For immunoblotting, purified heart membranes were prepared by differential centrifugation and purification on a sucrose gradient. 50 μg protein were loaded on a 7.5% SDS gel and blotted onto nitrocellulose as described previously (Dazert et al. 2003). Membrane vesicles from human placenta (50 μg) were used as positive control. The expression of BCRP was detected by chemiluminescence using the monoclonal BXP-21 antibody (diluted 1:1000; Alexis Biochemicals) and a horseradish-peroxidase-conjugated goat-anti-mouse IgG (diluted 1:2000; Bio-Rad, Munich, Germany).
Data Analysis
Patient data of heart auricular samples were screened for potential influence of the individual's medication, sex, body mass index, blood pressure, and ABCG2 mRNA expression. The amounts of specific ABCG2/18s rRNA mRNA in relation to ABCG2 genotype as well as to cardiomyopathy were compared by Mann-Whitney U-test using SPSS 11.0 software; p<0.05 was considered significant. All data are presented as mean ± SD as indicated.
Results
Localization of BCRP (ABCG2)
Immunohistochemistry and Immunofluorescence using confocal laser scanning microscopy localized BCRP predominantly in endothelial cells of capillaries and arterioles of human heart tissue for both ventricular and auricular samples. Colocalization experiments displayed simultaneous staining of the endothelial marker PECAM and ABCG2 in the endothelial wall (Figure 1). In contrast to the ABC transport protein ABCC5, the surrounding vascular smooth muscle fibers were devoid of BCRP. However, some staining of the plasma membrane of heart muscle fibers was also detected.
Immunoblot Analysis of BCRP (ABCG2)
Monoclonal anti-BCRP BXP-21 identified BCRP by Western blotting as a 70-kDa protein in both purified auricular and ventricular heart membranes. Samples from human placenta served as controls (Figure 1C, inset).
ABCG2 mRNA Expression in Human Heart Samples
In a series of 51 samples from human heart auriculum, ABCG2 mRNA was detected in all samples with substantial interindividual variability (ABCG2/18S mRNA range from 0.3 to 10.3 × 10−7 mean value 3.6 ± 1.7 × 10−7 Figure 1C, inset).
For investigating the influence of ischemic and dilative cardiomyopathy on the expression of ABCG2 mRNA, 15 ventricular human heart tissue samples were analyzed by means of real-time RT-PCR and related to 18S rRNA expression. Again, ABCG2 was detectable in all samples. ABCG2/18S rRNA expression in samples from cardiomyopathic hearts was significantly increased compared with that of nonfailing hearts for both dilative (ABCG2/18S rRNA: 1.08 ± 0.30 × 10−7; p = 0.028) and ischemic cardiomyopathy (1.16 ± 0.46 × 10−7; p = 0.009) when compared with nonfailing hearts (0.44 ± 0.26 × 10−7, Figure 2).
Influence of Patient Characteristics and Medication on ABCG2 mRNA Expression
ABCG2 mRNA was not influenced by patients' age, sex, or body mass index. Patient data were screened for common medications such as cardiac glycosides, beta blockers, statins, diuretics, nitrates, and angiotensin-converting enzyme inhibitors, which were rather common and prescribed in varying combinations to all individuals included in the study. None of the drugs or drug combinations had any significant influence on ABCG2 mRNA expression.
Influence of the Genotype on ABCG2 mRNA Expression
Variant allele frequencies were 6% for the 34G>A and 14% for the 421G>A SNP (n = 51). None of the SNPs alone or any of the haplotypes was associated with significantly altered ABCG2 mRNA expression (Figure 2).
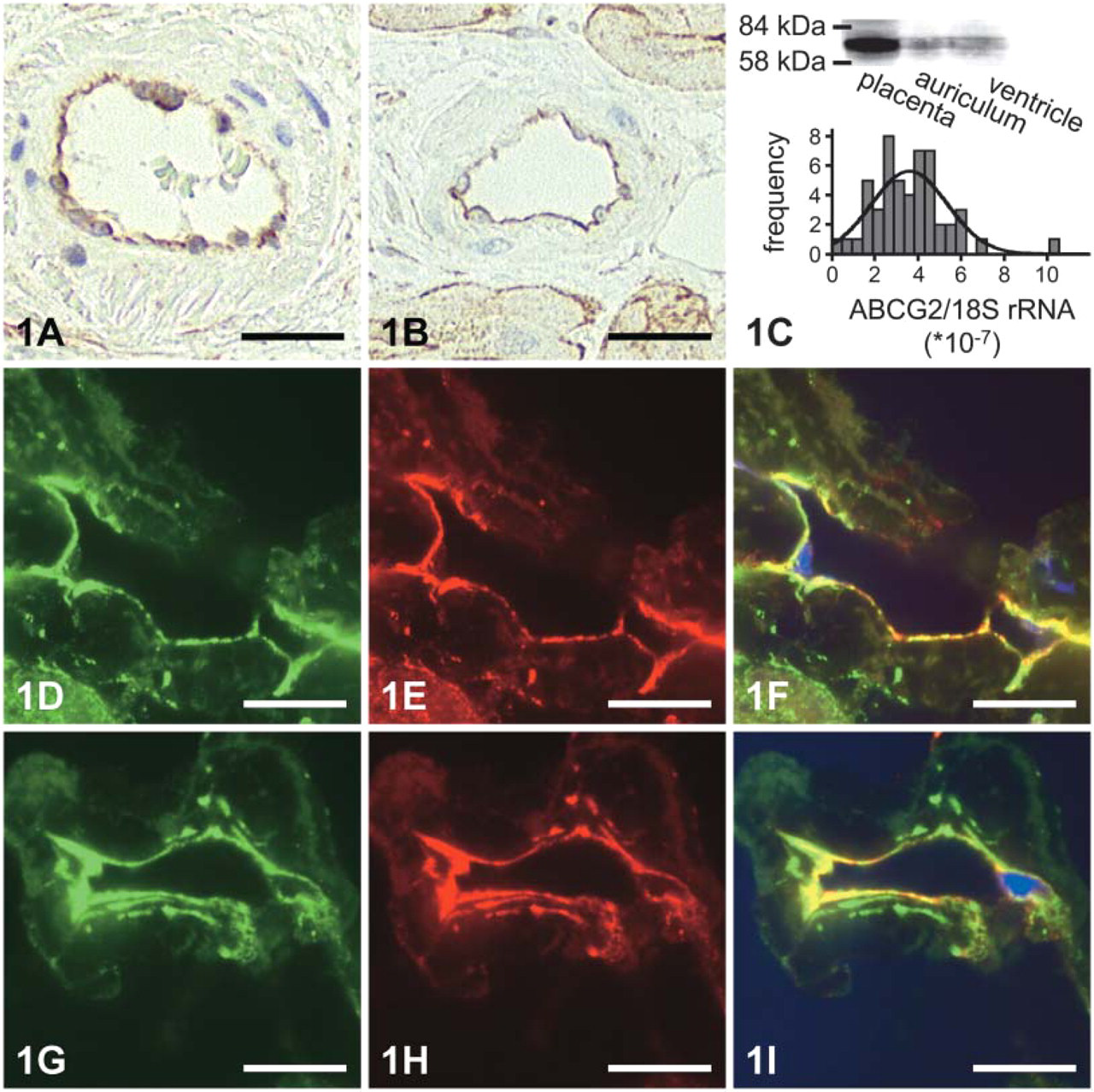
Detection of BCRP (ABCG2) in human heart (bars: 25 μm). (
Discussion
ABCG2 has been suggested as a marker for specific selection of SP stem cells (Zhou et al. 2001). Aside from the selection process, such marker proteins are often used to determine the localization of transplanted cells within the heart. ABCG2 expression in SP stem cells has been shown to significantly decrease during cellular differentiation (Martin et al. 2004). In view of the present study indicating significant expression of ABCG2 in normal human heart, ABCG2 appears not to be suitable as a marker protein for intracardiac localization of such cells. We observed a continuous staining of endothelial cells of arterial and capillary vessels both in diseased and normal heart tissue, along with a staining of plasma membranes of some cardiomyocytes.
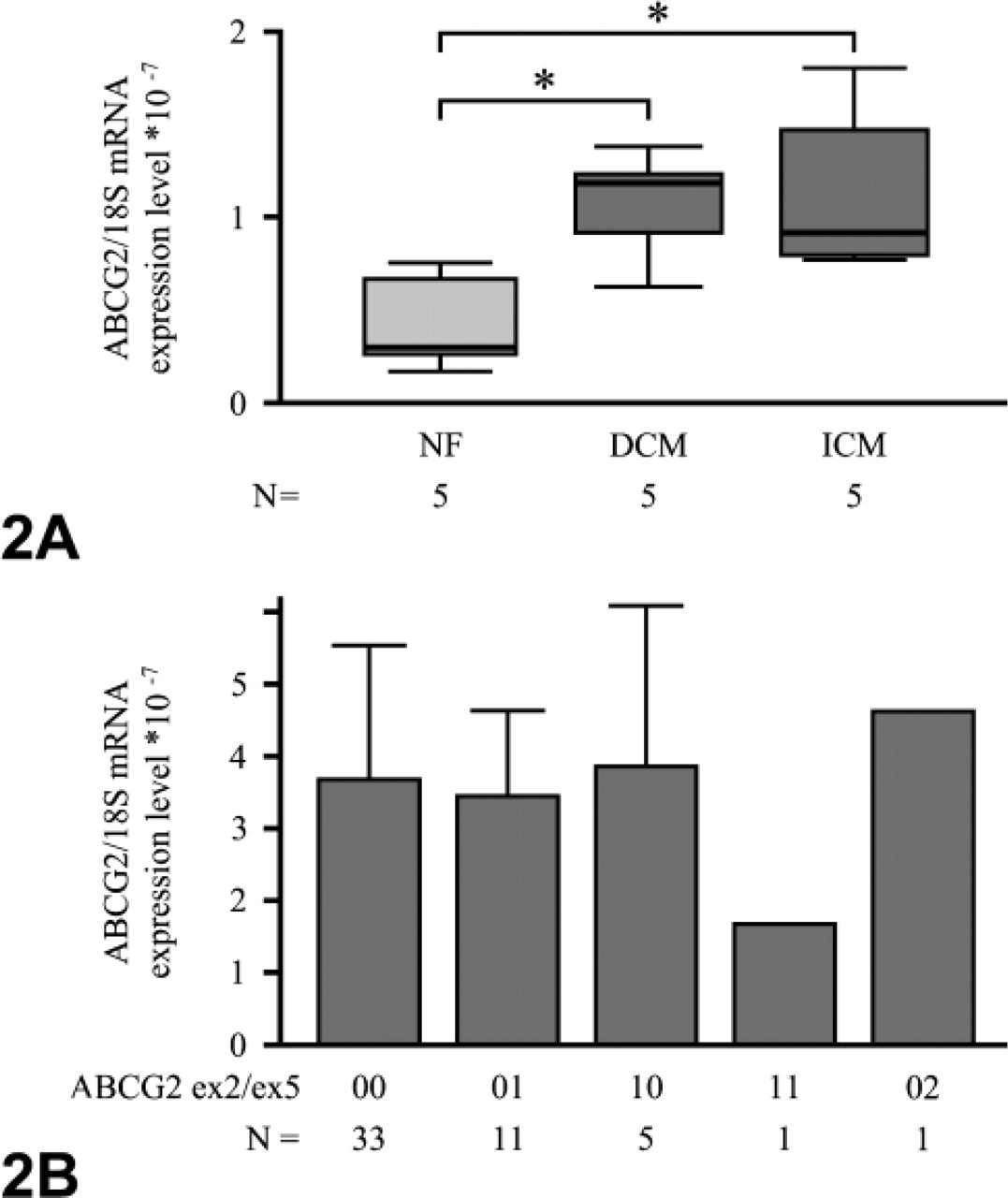
(
Localization of ABCG2 in endothelial cells in human heart is similar to that of ABCB1 (Meissner et al. 2002, 2004) but different from ABCC5, which is expressed in cardiac myocytes, smooth muscle cells, and endothelial cells (Dazert et al. 2003). The question arises whether the putative function of ABCG2 in human heart can be assessed based on its endothelial expression. Expression of ABCG2 has been described in the apical membrane of trophoblast cells of placenta, the apical membrane of enterocytes, the apical membrane of the lactiferous duct in mammary glands, in bile canalicular membranes of hepatocytes, and in the blood–brain barrier. Taken together, these sites of expression suggest a major role in detoxification because ABCG2 contributes to transport across the apical membranes of these cells, thereby protecting cells from drug effects and prohibiting entry of compounds into organ systems (Maliepaard et al. 2001).
In line with these findings, expression in endothelial cells in human heart could contribute to reduce or prevent cardiac uptake of ABCG2 substrates. Among the substrates of ABCG2 are a variety of cardiotoxic drugs such as mitoxantrone, topotecan, SN-38, and the HIV-1 nucleoside RT inhibitors ritonavir and saquinavir (Allen et al. 1999; Volk and Schneider 2003; Wang et al. 2003). Mitoxantrone, which, in addition to cancer chemotherapy, was recently introduced to multiple sclerosis therapy, has been described as cardiotoxic (Ghalie et al. 2002). Cardiac uptake of these substrates for ABCG2 can be prevented by endothelial expression, thereby modulating drug effects.
Our data indicate a pronounced interindividual variability of cardiac ABCG2 expression, which can be due to environmental factors (drugs, disease processes) or genetic influence. Aside from its role in drug resistance, regulation of ABCG2 expression is not well understood. We did not find any association between drugs administered and cardiac ABCG2 levels. A recent article indicated that hypoxia increases ABCG2 mRNA expression and thereby cell survival rates in cell culture experiments of hematopoietic stem cells (Krishnamurthy et al. 2004). Our previous work described a significant influence of disease on cardiac expression of ABCB1 (P-gp). Patients with dilative cardiomyopathy had a significantly reduced expression of this protein. In contrast, ABCG2 was upregulated in patients with both dilative and ischemic cardiomyopathy, indicating a differential effect of heart disease on drug transport proteins. In view of the coexpression of ABCG2 and ABCB1 at the apical site of various cells and some overlap in substrates, a combined role of both proteins in elimination of compounds from human heart can be assumed.
Although increased ABCG2 in vitro activity in cancer cell lines has been shown to be influenced by acquired mutations in the ABCG2 gene (Volk and Schneider 2003), the in vivo contribution of hereditary single nucleotide polymorphisms to ABCG2 expression levels is unclear. Among the four identified naturally occurring single nucleotide polymorphisms, 34G>A (V12M) and 421C>A (Q141K) were most common in diverse populations of different ethnic origin in North America (Zamber et al. 2003). Though the study unveiled a wide range of different individual ABCG2 mRNA expression levels, these data did not correlate to either BCRP protein expression or individual ABCG2 genotypes (Zamber et al. 2003). Transfection experiments in LLC-PK1 cells confirmed altered ABCG2 function for two common ABCG2 SNPs, which could in part be attributed to a different localization of ABCG2 within the cells (Mizuarai et al. 2004). However, the impact of SNPs within the coding region of the ABCG2 gene remains to be discussed controversially (Honjo et al. 2002). In contrast to the described inherited mutations, acquired mutations of tumor cell lines were shown to confer resistance to a number of anticancer agents such as anthracyclines (Volk and Schneider 2003), whereas cells transfected with the wild-type were not (Honjo et al. 2001). Explicit search for the SNP in question (482R>T/G) did not unveil its occurrence in a population of 90 DNAs from the single nucleotide polymorphism discovery resource (Honjo et al. 2002).
Given a defense role of ABCG2 in men, influence on its functional state by anticancer drugs such as imatinib or gefitinib (Houghton et al. 2004; Stewart et al. 2004) may unintentionally lead to increased toxic effects of ABCG2 substrates on organs “protected” by ABCG2 expression in capillaries. Furthermore, ABCG2 expression in heart may be inhibited by antiretroviral drugs such as ritonavir or saquinavir (Gupta et al. 2004), thereby potentially damaging the heart, which was reported to be more susceptible to myocardial infarction after HIV therapy (Friis-Moller et al. 2003).
Taken together, our data indicate variable expression of ABCG2 in human heart endothelial cells, which is influenced by cardiomyopathies. Therefore, cardiac ABCG2 expression may be an obstacle for the detection of side population stem cells in stem cell therapy. Our data, based on 51 auricular heart samples, did not indicate a significant influence of the two studied most common SNPs in the ABCG2 gene on ABCG2 mRNA expression. However, a genetic influence on the transporter activity cannot be excluded. Moreover, cardiac ABCG2 expression suggests a protective role against substances entering the heart. Regulation as a consequence of diseases such as cardiomyopathies may alter susceptibility toward these substrates.
Footnotes
Acknowledgements
Supported by grants from the Deutsche Forschungsgemeinschaft (SFB TR 19–4) to G.J. and H.K.K., German Federal Ministry of Education and Research to H.K.K. (BMBF 01ZZ0103), and from the Karl und Lore Klein-Stiftung, Oy-Mittelberg, Germany, to K.M.
We acknowledge the excellent technical assistance by Tina Brüggmann, Ingrid Geissler, and Anke Wolter, Departments of Pharmacology and Pathology, Peter Holtz Research Center of Pharmacology and Experimental Therapeutics, Ernst-Moritz-Arndt-University Greifswald, Germany.