
Abstract
Ornithine decarboxylase (ODC) is the key enzyme in the polyamine synthesis pathway and is overexpressed in a variety of cancers. We have performed a detailed immunostaining analysis of the expression of ODC in normal, benign prostatic hyperplasia (BPH), and cancerous prostate tissues. We conclude that ODC is overexpressed in both BPH and neoplastic tissues and that ODC overexpression appears to be an early event in prostate carcinogenesis. The extent of overexpression decreases as cancer progresses. Interestingly, ODC overexpression was also detected in patients who underwent androgen ablation therapy, suggesting ODC overexpression may contribute to the androgen-independent survival of prostate cancer cells. ODC is perinuclear localized in BPH samples but is diffusely cytoplasmic in cancer samples. Having shown ODC overexpression in human prostate cancer, we developed prostate-specific ODC transgenic mice to further investigate whether ODC overexpression alone is a causal factor in prostate carcinogenesis. RT-PCR and immunostaining confirmed that ODC was overexpressed in a subset of prostate epithelial cells. Although minor nucleoli enlargements in some tissues were detected, gross morphological changes were not observed in transgenic prostates. Therefore, overexpression of ODC alone in this subset of prostate epithelial cells is not sufficient to induce prostate carcinogenesis.
O
However, although it is clear that ODC overexpression is associated with cancer development, a definitive causal role of ODC overexpression in carcinogenesis has only been shown in NIH/3T3 fibroblast cells (Moshier et al. 1993). In transgenic ODC mice, only a promotive role can be shown in the skin after 7,12-dimethylbenzanthracene (DMBA) treatment (Megosh et al. 1995; Chen et al. 2000).
ODC activity is also increased in prostatic cancer tissues and their respective secretory fluids (Mohan et al. 1999; Bettuzzi et al. 2000; Rhodes et al. 2002). Furthermore, in the widely used transgenic mouse model of prostatic cancer (TRAMP), ODC activity is high in cancer tissues, and DFMO treatment resulted in a significant decrease in tumor incidence, volume, and distant metastasis (Gupta et al. 2000). Despite this, a detailed study comparing the expression patterns of ODC at the cellular level in benign prostatic hyperplasia (BPH) and prostate cancer tissues has not been conducted. Furthermore, it has not been established whether overexpression of ODC is involved in the initiation or promotion of prostatic carcinogenesis. To address these issues, we first conducted a detailed immunohistochemical (IHC) comparison between normal, BPH, prostatic intraepithelial neoplasia (PIN), and prostatic cancer tissues. This revealed distinct expression patterns and confirmed overexpression of ODC in prostatic cancer tissues. Based on the expression pattern obtained from our IHC studies, we developed a mouse model that specifically expressed the ODC transgene in the prostatic epithelial cells by placing the human ODC gene under the control of the minimal promoter of the rat Probasin gene (Pb). Although no specific phenotype has been observed in these mice, involvement of ODC during carcinogenesis cannot be ruled out because consistent overexpression was not achieved in the transgenic mouse prostate.
Materials and Methods
Patients and Specimens
Normal organ donor human prostatic tissues were obtained from the New South Wales (NSW) Red Cross. BPH and prostate cancer biopsy tissues were obtained from the Royal Prince Alfred Hospital (RPAH), Sydney, Australia). Whole organ donor prostate glands were dissected by a urological surgeon into transitional/central and peripheral zones using urethra and ejaculatory ducts as landmarks. The study was approved by the RPAH Human Ethics Committee. Paired ventral mouse prostates were collected from Ketalar-overdosed mice via surgical exploration of the abdominal cavity and dissection. Fresh tissues were fixed in 4% buffered formaldehyde, pH 7, and embedded in paraffin.
IHC on Human Prostatic Tissue
IHC staining was performed on normal (n=7), BPH (n=6), PIN (n=6), and prostatic cancer tissues (n=27). Prostatic cancer tissues could be further separated on the basis of androgen dependence. Cancers were either androgen dependent (n=19) or androgen independent (n=8). The cancers were further classified on the basis of Gleason score as determined by a qualified pathologist.
Tissue slides were antigen retrieved in 500 ml of Tris EDTA (10 mM Tris Base, 1 mM EDTA solution, 0.05% Tween 20, pH 9.0) and microwave treated twice for 10 min each at medium high in a 650-W microwave oven. The mouse anti-human ODC monoclonal antibody (ODC-29, mouse ascites fluid) was obtained from Accurate Chemicals (Westbury, NY), and immunoreactivity was visualized by indirect peroxidase staining with Vectastain ABC kit (Vector Laboratories; Burlingame, CA), following manufacturer's instructions.
Following immunodetection, a pathologist attributed a relative stain intensity to the section. The relative stain intensity could be anywhere from 0 to 3 (3 being the most intense). The relative stain intensity is a measure of difference between the normal and abnormal cells in each section. Therefore, the comparison of ODC levels between cancer and normal, BPH and normal, and PIN and normal was made from tissues found within the one section, thus minimizing the need for inter-patient comparisons.
Transgene Construction and Preparation of Transgenic Mice
Pb promoter (−426 to +28) was PCR amplified from rat genomic DNA with primers containing BamHI and XhoI sites (5′-TGGATCCAGCTTCCACAAGTG CATTT-3′ and 5′-TCTCGAGTCTGTAGGTATCTGGACCT-3′). ODC was amplified from human prostate cDNA with primers containing XhoI and NotI sites (5′-ACTCGAGCATGAACAACTTTGGTAATGA-3′ and 5′-CGCGGCCGCAAAT CCCTTAATTCAAGCTA-3′). Both Pb and ODC were cloned into pBK-RSV vector (Stratagene; La Jolla, CA) through the intermediate TA cloning vector pGEM-T EASY (Promega; Madison, WI) using standard cloning techniques. Twenty μg of the final vector was digested with BamHI and MluI and microinjected into fertilized C3H mouse oocytes as described previously (Allan et al. 2001). DNA from founder mouse toes was extracted, and transgenic mice were confirmed by PCR amplification of the DNA with primers PbF1 (5′-GACACTGCCCATGCCAATCA) and ODCR1 (5′-ACAGCGCTCAACAATCC GATC-3′). Routine screening of mice was carried out by amplifying toe DNA.
Confirmation of Transgene Expression by RT-PCR
Total RNA was extracted from mouse prostates harvested from heterozygous and non-transgenic mice with Tri-Reagent (Invitrogen; Carlsbad, CA). Two μg of the total RNA were treated with DNase I (Invitrogen) and reverse transcribed with Superscript II reverse transcriptase (Invitrogen). The resulting cDNA (1 μl) was either subjected to 40 cycles PCR with primers ODCVF (5′-TATTACAATTCACTGGCCGTCGTTT-3′) and ODCVR (5′-AATGAGCCTTGGG ACTGTGAATCTA-3′) for ODC transgene or 30 cycles with primers ACTF (5′-CCGGGACCTGACTGACTACCT-3′) and ACTR (5′-GCATTTGCGGTGGACGA-3′) for β-actin.
Confirmation of Transgene Expression by IHC
To confirm overexpression of the ODC transgene at the protein level, IHC was performed on prostate harvested from ODC transgenic mice at 33 weeks (n=2), 24 weeks (n=6), 18 weeks (n=3), and 12 weeks (n=2) of age, together with age-matched wild-type prostates (n=8). The antibody was diluted 1:200 and used in conjunction with the Animal Research Kit (DAKO; Carpinteria, CA) following manufacturer's instructions. All images were taken with Olympus BX51 microscope (Olympus; Tokyo, Japan) under the same conditions. Hematoxylin and eosin staining was performed on the same samples, and the morphology was evaluated by a qualified pathologist without knowing the transgenic status.
Results
ODC Expression Pattern in Normal, BPH, PIN, and Cancer Tissues
ODC protein expression was examined by IHC in normal, BPH, PIN, and cancer tissues of human prostates (Figure 1; Table 1). Cytoplasmic immunoreactivity was detected in the secretory epithelial cells and endothelial cells in normal prostates (n=7) and was distributed evenly within the cells. No expression was detected in basal epithelial cells, fibroblasts, or smooth muscle cells. There was no significant difference between expression patterns of the peripheral zone and transitional/central zone of the normal prostates (Figure 1). In BPH samples (n=7), strong perinuclear staining was detected mainly at the basal side of the secretory epithelial cells. Compared with adjacent normal tissue, ODC expression in BPH secretory epithelial cells is stronger, and the cellular location is also unique (Figure 1).
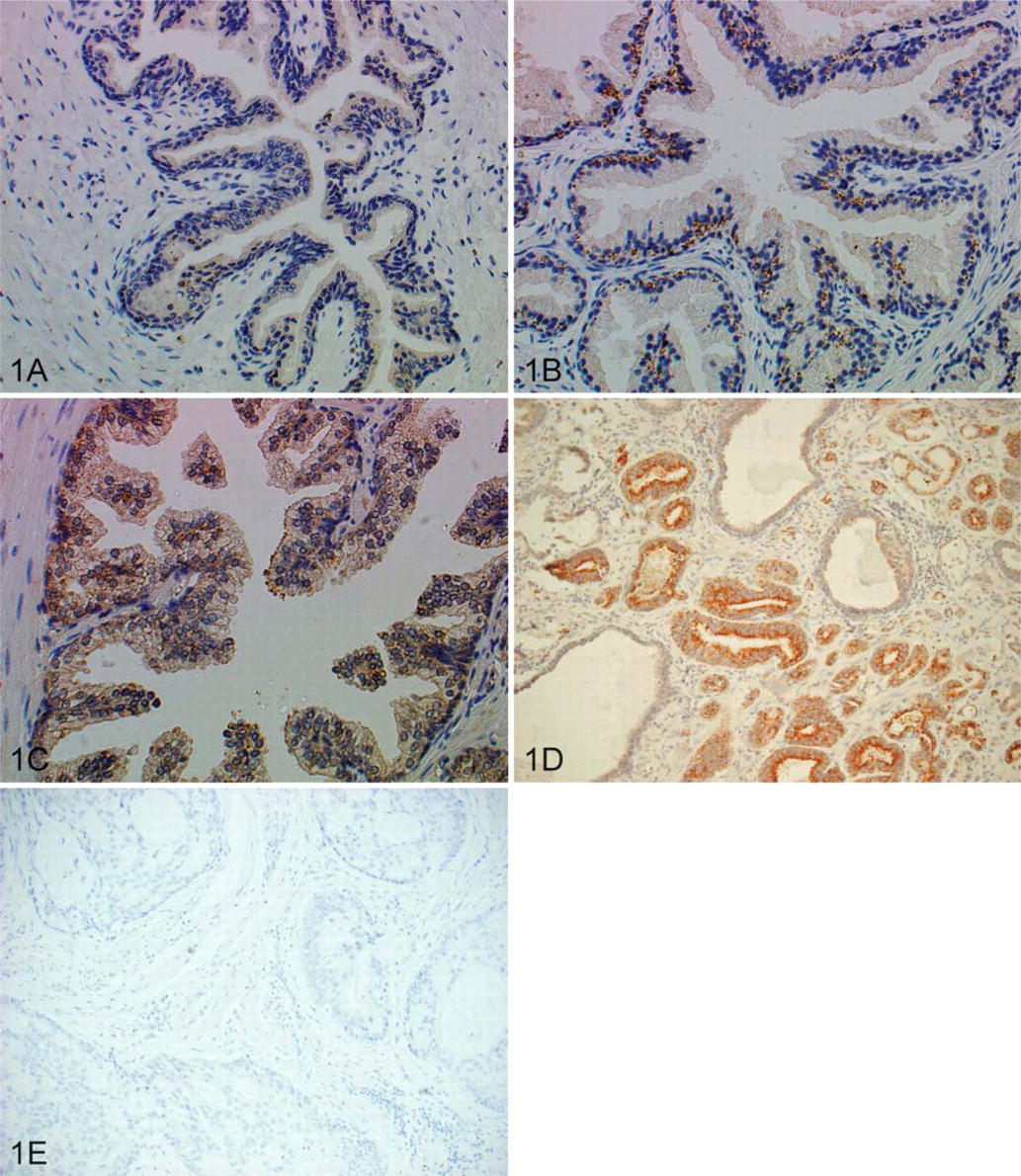
Protein expression pattern of ornithine decarboxylase (ODC) in human prostate. (
High-grade PIN has been shown to be linked to prostate cancer (Ashida et al. 2004) and is recognized as having a significant predictive value for prostatic cancer. We therefore analyzed ODC expression in PIN from radical prostatectomy patients. As shown in Figure 1C, overexpression (compared with normal tissue) of ODC in the cytoplasm of secretory epithelium for high-grade PIN can be detected in all samples (n=6). Expression in adjacent normal glands is comparable to that in the organ donor prostates. ODC expression in PIN is more diffuse compared with BPH, and expression levels are similar between the two.
All prostate cancer tissue analyzed (with the exception of one specimen) showed an increase in ODC expression between the cancerous and adjacent normal tissue, and immunoreactivity was strongest at the apical side of the epithelial cells (Figure 1D). Although this was true for both androgen-dependent and androgen-ablated sections, ODC expression was markedly higher in the non-androgen-ablated sections. Analysis (Student's t-test) between sections having a relative stain intensity of 1, 2, or 3 showed that there was no statistical correlation between Gleason score and relative stain intensity.
Confirmation of Presence of Transgene mRNA in the Mouse Prostate
To investigate whether ODC overexpression alone can be a causal factor in prostatic cancer, we generated a transgenic ODC mouse line with ODC expression driven by prostate-specific rat Pb promoter. Because mouse and human ODC catalyze the same reaction and show 90% identity in amino acid sequence, human ODC cDNA was used as the transgene. To confirm the presence of transgene-derived ODC mRNA in the prostate, we amplified cDNA with primers specific for the vector sequence section of the transgene so that any amplification detected should have arisen from the transgene. As shown in Figure 2, ODC mRNA expression was detected only in the heterozygote ODC transgenic mouse prostate, whereas β-actin quantities remained constant in both transgenic and non-transgenic mouse prostates. This confirms ODC transgene expression in the prostates of transgenic mice at the mRNA level.
Ornithine decarboxylase relative intensity in human prostate cancer tissues compared to adjacent normal tissues
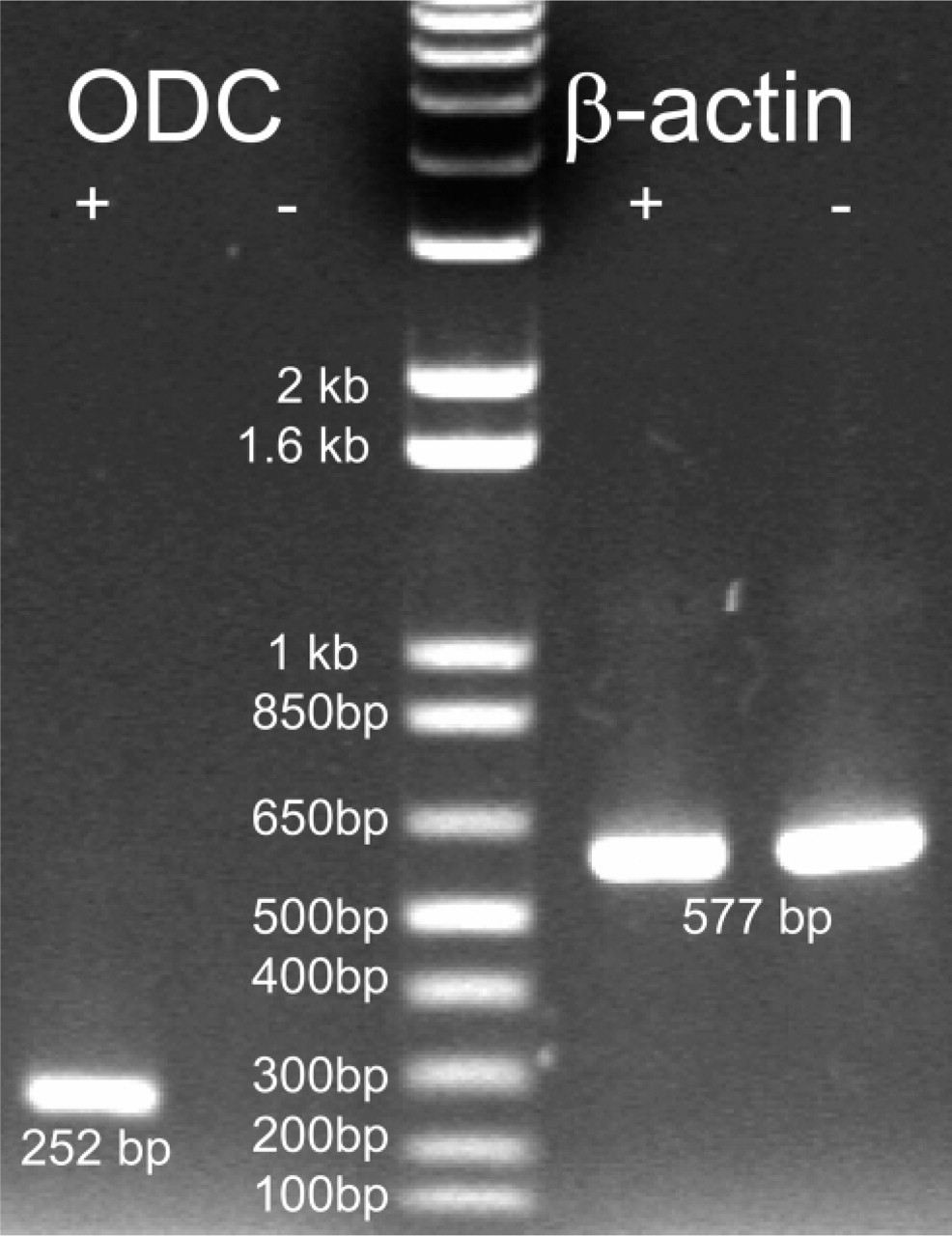
RT-PCR analysis showing expression of ODC transgene mRNA in mouse prostate. Lane 1, transgene was expressed in heterozygous mouse prostate; Lane 2, transgene expression was not detected in wild-type mouse prostate; Lane 3, 1 kb plus DNA marker (Invitrogen); Lanes 4 and 5, β-actin expression in heterozygous and wild-type mouse prostates.
Protein Levels of the ODC Transgene in the Mouse Prostate
To evaluate the expression level of ODC protein in the transgenic mouse prostates, we performed IHC staining on the ventral lobes of both transgenic and non-transgenic prostates between 12 and 33 weeks of age. Ventral lobes are selected because previous reports have shown that the minimal Pb promoter is more active in these lobes (Greenberg et al. 1994). Because the monoclonal antibody used was of mouse origin, the ARK kit (DAKO) was utilized to avoid nonspecific binding of the secondary antibody to endogenous mouse IgGs. As shown in Figure 3, the antibody stained epithelial cells exclusively; staining was not detected in the stromal compartment. Within the same transgenic prostate, there were areas with high ODC expression (Figure 3) and areas where ODC expression was barely detectable (Figure 3). Similarly, in non-transgenic prostates, there were also areas with moderate (Figure 3) or no (Figure 3D) ODC expression. However, all 13 transgenic prostates contained areas with ODC expression levels above those detected in the non-transgenic prostates, indicating that ODC overexpression was at least achieved in a subset of cells. No significant difference in expression pattern was observed in prostates of different ages. Tissue sections of the same prostates were also H–E stained and evaluated blindly by a pathologist. Although significant changes in morphology or histology were not detected, nucleoli enlargements were detected in some prostatic cells of the transgenic mice.
Discussion
In the present work we examined in detail the protein expression pattern of ODC in normal, BPH, and prostatic cancer tissues and clearly revealed overexpression of ODC at protein level in both BPH and cancer samples. Interestingly, apart from the changes at the protein level, BPH samples showed perinuclear localization patterns distinct from normal and cancer samples. In keratinocytes, ODC localization has been shown to correlate with changes in the cytoskeleton network. In normal human epidermal keratinocytes, ODC exhibited perinuclear expression patterns, which became diffusely cytoplasmic after cytoskeleton disruption by TPA, cytochalasin B/D, or DFMO (Pomidor et al. 1995). The different localization patterns thus may indicate a more stable cytoskeleton in BPH and disruption of the cytoskeleton in PIN and prostate cancer cells. This difference serves as further evidence that PIN rather than BPH may be the precursor of prostate cancer.
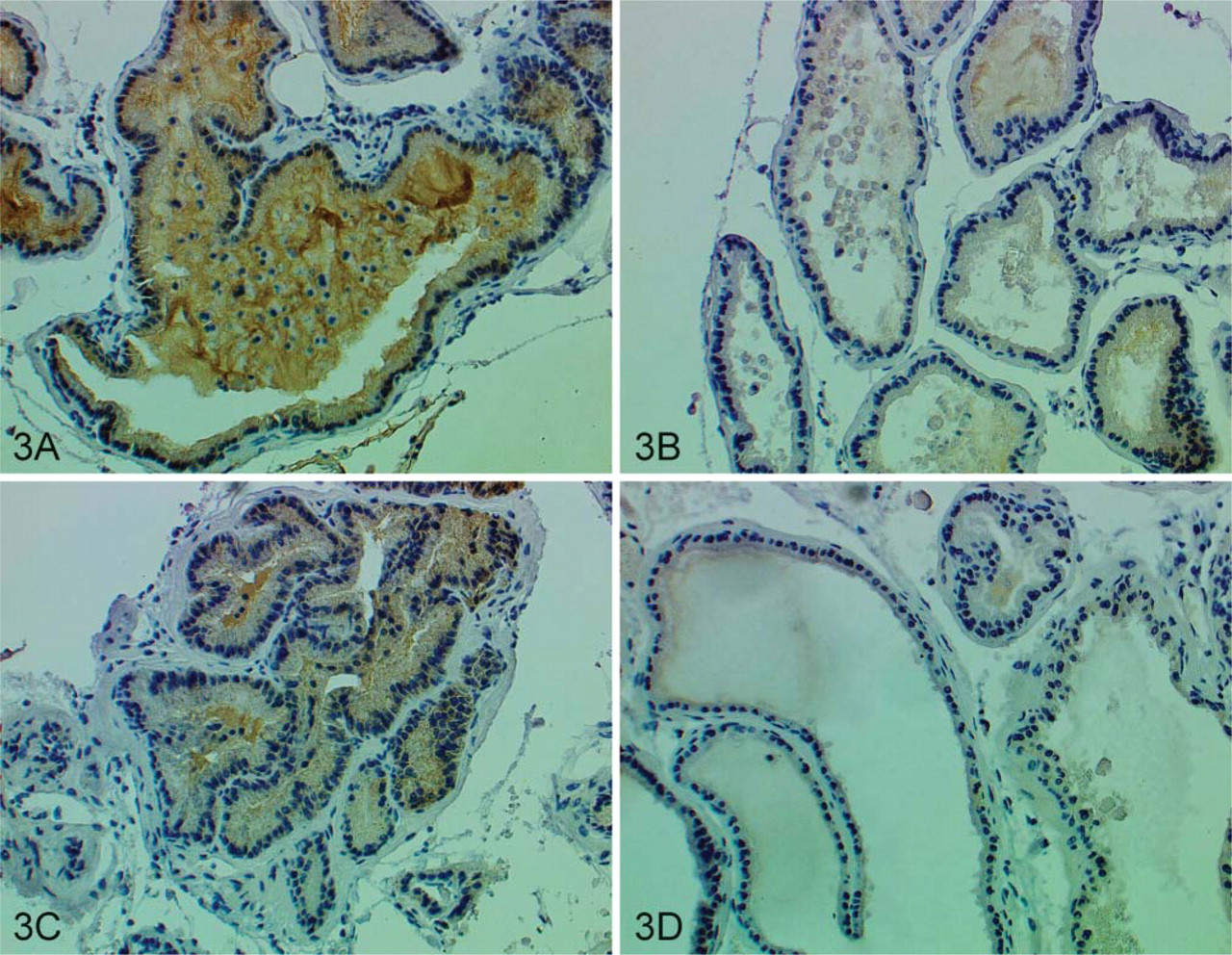
ODC protein expression in mouse ventral prostates. (
ODC overexpression was detected as early as in PIN, suggesting ODC could be involved in the early stages of the carcinogenesis process and could be involved in promoting the progression of PIN to invasive cancer. Overexpression of ODC can be detected in all cancer tissues examined, and although the staining patterns were heterogeneous, the increase of ODC can be concluded to be an early event in prostate carcinogenesis.
To investigate the role of ODC overexpression in prostatic cancer, we developed a prostate-specific ODC transgenic mouse model. Immunostaining results suggest that ODC was achieved at least in a subset of the prostate epithelial cells. The reason that overexpression of ODC was not achieved in all the prostate epithelial cells may be due to the tight posttranslational regulation of ODC by ODC antizyme (Murakami et al. 1992), as the normal transcriptional (promoter and enhancer) regulatory sequences have been replaced by the Pb promoter, and the translational regulatory sequences, the 5′ and 3′ untranslated regions (UTR) (Manzella et al. 1991), have been eliminated in the transgene. Prostate epithelial cells display different ODC regulation levels intrinsically, as heterogeneous ODC expression has also been shown in wild-type prostates in previous studies (Muller et al. 1986). Their study has revealed the existence of three types of prostatic epithelial cells displaying high, medium, or low levels of ODC. Therefore, the increased ODC levels detected in some regions of the transgenic mouse prostate may reflect true overexpression in cells that have minimal antizyme regulation (those cells that have high wild-type ODC levels).
Despite that overexpression of ODC was achieved in a subset of the transgenic prostate epithelial cells, cell transformation was not observed, which means that overexpression of ODC alone in this subset of cells could not serve as a causal factor in prostate carcinogenesis. However, a conclusion on the role of ODC in prostate cancer cannot be made without the overexpression of ODC in all of the transgenic prostate epithelial cells, which requires the escape from antizyme regulation. It should be noted that in the only transgenic ODC model that developed skin tumors after chemical carcinogen initiation, the ODC transgene is a C-terminal truncated copy (Megosh et al. 1995), which could no longer be degraded by the endogenous antizyme. We have chosen to use the full human ODC gene because in an earlier study the full human ODC including its promoter has been used as the transgene to achieve overexpression in most mouse tissues (Halmekyto et al. 1991). We reasoned that because the ODC used was of human origin, it may be able to escape the mouse antizyme control without C-terminal truncation. However, our result indicates this may not be the case. Therefore, in future studies the C-terminal truncated ODC gene should be used instead to achieve more stable overexpression of ODC.