
Abstract
Rupture of vulnerable atherosclerotic plaques that are characterized by extensive neointimal accumulation of lipid is a cause of acute coronary syndromes. To identify whether insulin resistance alters atherogenesis, we characterized the composition of atherosclerotic lesions in the proximal aortas in mice deficient in apolipoprotein E (ApoE−/-) and in ApoE−/- mice in which insulin resistance was intensified by a concomitant heterozygous deficiency in insulin receptor substrate type 2 (IRS2+/- ApoE−/- mice). In addition, we characterized the effect of an insulin sensitizer, pioglitazone, on the atherogenesis in IRS2+/- ApoE−/- mice. The extent of the aortic intima occupied by lesion was increased in the IRS2+/- ApoE−/- compared with ApoE−/- mice (79 ± 3% compared with 68 ± 8%, p<0.05). Treatment with pioglitazone decreased the neointimal content of lipid in 20-week-old mice from 50 ± 6% to 30 ± 7%, p=0.005 and decreased the cellularity reflected by the multisection cross-sectional areas of lesions comprising cells in atheroma from 24 ± 1% to 19 ± 3%, p=0.018. Accordingly, genetically induced intensification of insulin resistance increases atheroma formation. Furthermore, attenuation of insulin resistance by treatment with pioglitazone decreases accumulation of lipid in the neointima.
Keywords
Myocardial infarction is two- to fourfold more common in persons with diabetes than in age- and gender-matched persons without diabetes (Haffner et al. 1998). It also occurs more frequently in persons without diabetes but with insulin resistance reflected by increased concentrations in blood of insulin under fasting conditions (Miettinen et al. 1998). One factor potentially contributing to the greater incidence of myocardial infarction in persons with diabetes and in those with insulin-resistant states is a propensity to generate atherosclerotic plaques that are particularly prone to rupture. Accordingly, this study was undertaken to determine whether insulin resistance alters the composition of atheroma by characterizing the composition of atherosclerotic lesions in a 300-μm region of the proximal aorta in hyperlipidemic mice that were genetically deficient in apolipoprotein E (ApoE−/-) and in ApoE−/- mice in which insulin resistance had been intensified genetically by concomitant heterozygous deficiency of insulin receptor substrate type 2 (IRS2+/-) (Previs et al. 2000). Mice with heterozygous deficiency of IRS2 were used because homozygous IRS2-deficient mice exhibit only limited survival unless treated with exogenous insulin (Withers et al. 1998).
A second objective of the present study was to determine whether an intervention designed to increase insulin sensitivity—treatment with the thiazolidinedione, pioglitazone—would alter the composition of atherosclerotic plaques in hyperlipidemic mice with intensified insulin resistance (IRS2+/- ApoE−/- mice). Treatment with pioglitazone decreases the concentration in blood of both insulin and glucose in mice, rats, and human subjects with type 2 diabetes reflected by fasting concentrations of both in blood and a decrease in the concentration in blood of glycosylated hemoglobin (HbA1c) (Hofmann et al. 1992; Gerber et al. 2003; Larsen et al. 2003). The drug alters the lipid profile as reflected by decreased concentrations in blood of total cholesterol, increased concentrations of HDL cholesterol, modestly increased concentrations of LDL cholesterol, and decreased concentrations of triglycerides. In the present study, the effect of pioglitazone on the composition of atherosclerotic plaques was determined in 10- and 20-week-old IRS2+/- ApoE−/- mice given daily pioglitazone orally compared with that in IRS2+/- ApoE−/- mice given vehicle alone.
Materials and Methods
Use of Animals
Transgenic mice were used in a protocol approved by the University of Vermont Institutional Animal Care and Use Committee. ApoE−/- mice, congenic with respect to the C57Bl6 background, were purchased from Jackson Laboratories (Bar Harbor, ME). IRS2+/- mice (Withers et al. 1998) used for breeding were kindly provided by the Howard Hughes Medical Institution and the Joslin Diabetes Center (Dr. Morris White, investigator). IRS2+/- mice were congenic, having been backcrossed 10 times into the C57Bl/6 line. The congenic IRS2+/- mice were crossed with ApoE−/- mice to generate IRS2+/- ApoE−/- mice. To avoid potential influences of gender and estrus cycles, only male animals were used in the experiments described.
We compared the composition of atherosclerotic lesions in the aortas of two groups of mice; (a) in 20-week-old ApoE−/- mice compared with IRS2+/- ApoE−/- mice, and (b) in 10- and 20-week-old IRS2+/- ApoE−/- mice treated daily with 20 mg/kg of pioglitazone compared with IRS2+/- ApoE−/- mice given vehicle alone (Hofmann et al. 1992; Larsen et al. 2003). The pioglitazone was mixed in peanut butter (100 mg) leading to its complete consumption daily.
All mice studied were fed a high-fat diet from weaning (20% fat, 1.5% cholesterol; Teklad, Harlan Laboratories, Madison, WI). The heart and aorta were removed and immersed into 3% formaldehyde (freshly prepared from paraformaldehyde) in PBS (0.01 M phosphate buffer, 0.15 M NaCl, pH 7.3), and processed for sectioning with a cryostat at a thickness of 10 μm in a 300-μm region of the aorta beginning at the aortic valve, as previously described (Taatjes et al. 2000; Wadsworth et al. 2002).
Histochemical Staining
Sections were stained with oil red O (Pearse method) to detect lipid, and picrosirius red to distinguish collagen present in the lesions, as previously described (Taatjes et al. 2000; Wadsworth et al. 2002; Schneider et al. 2004). 4',6-Diamidino-2-phenylindole (DAPI) staining (Roche Diagnostics Corporation, Indianapolis, IN) of cell nuclei was used to demonstrate overall cellularity in lesions. Following oil red O staining, slides were incubated with a 5-μg/ml DAPI/PBS solution for 15 min.
Imaging
Sections were imaged with an Olympus BX50 upright light microscope (Olympus America, Lake Success, NY) with an attached Optronics MagnaFire digital camera (Optical Analysis, Nashua, NH), and images were obtained with the use of MagnaFire software (version 2.0), as previously described (Wadsworth et al. 2002). Sections stained with oil red O were illuminated with transmitted light and imaged with a 4X objective lens, as previously described (Wadsworth et al. 2002). Autofluorescence images were captured as well to reveal the vessel wall better and ultimately to aid in image processing. Autofluorescence images were captured subsequent to oil red O imaging of the same section by turning off bright field illumination and changing the filter cube turret from the ultraviolet/transmitted light position to the green emission position (blue excitation ex: 460–490, dichromatic beamsplitter DM 500, bandpass barrier filter BA 515–550 IF). A constant exposure time was set, and images were saved in 1280 × 1024 pixel 8-bit grayscale format.
DAPI-stained sections were imaged by epifluorescence microscopy with the use of a 10X objective lens. An exposure was set with the “clip detect” application to avoid pixel saturation (Wadsworth et al. 2002). Images were saved as 1280 × 1024 pixel 8-bit grayscale files for further analysis.
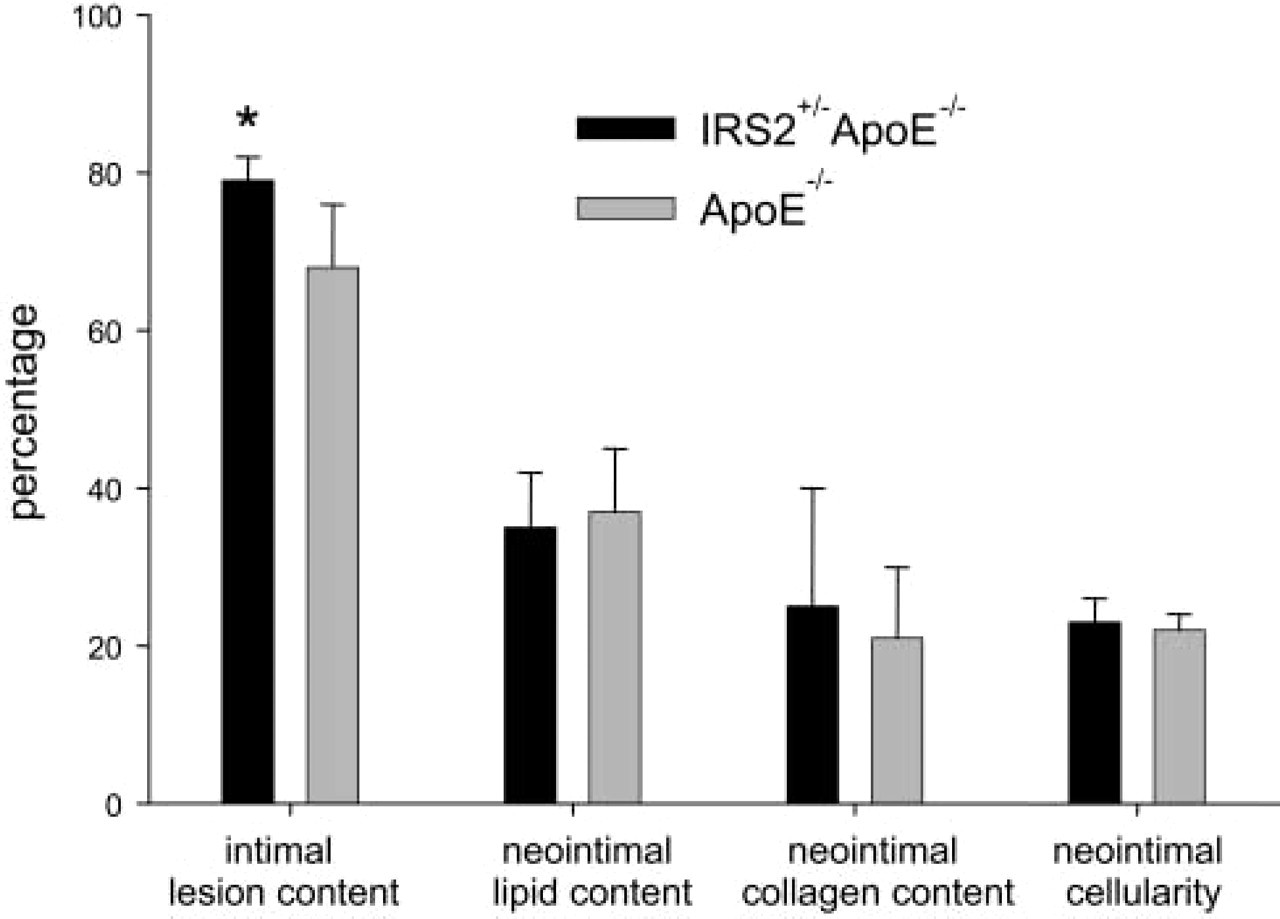
Features of the neointima in hyperlipidemic ApoE−/- mice compared with those in hyperlipidemic mice with intensified insulin resistance (IRS2+/- ApoE−/- mice).
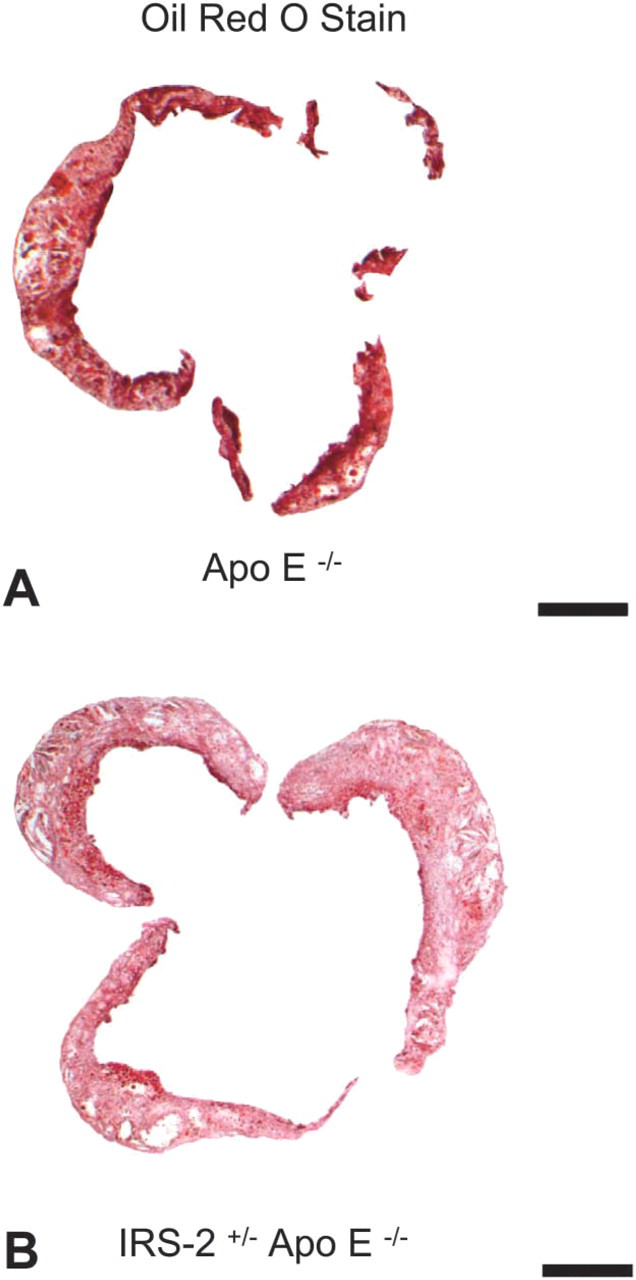
Representative images (transmitted light, bright field microscopy images captured with X4 objective lens) of oil red O–stained aorta; 10-μm-thick sections demonstrate total lesion area. Surrounding cardiac tissue, including media, has been cropped away with the use of Adobe Photoshop to facilitate quantitative analysis. In each section, the percentage of the intima is determined as total intimal area (μm2) occupied by atheroma (μm2). (
Images stained with picrosirius red were obtained with polarized light microscopy according to our previously published protocol (Wadsworth et al. 2002).
Computer-assisted Image Processing
Digital images were opened in Adobe Photoshop (version 7.0; Adobe Systems, San Jose, CA) for cropping to isolate the lesion. Autofluorescent images were cropped to isolate the artery. With the use of the cropped autofluorescent image as a guide, oil red O stained images were cropped to separate the lesion from the surrounding vessel wall, as previously described by Wadsworth and coworkers (Wadsworth et al. 2002).
Captured 10X DAPI images were cropped in the same manner, with the additional use of the rectangular marquee tool to ensure minimal overlap when cropping consecutive images of each vessel. Images cropped with the use of the oil red O images as guides showed isolated lesions with black backgrounds.
The 4X picrosirius red polarized light microscopy images were cropped as previously described (Wadsworth et al. 2002).
Qualitative Analysis
A qualitative visual assessment (scale of 1–5) of lesion composition was performed as previously described on each uncropped image by an individual who was blinded with respect to the source of the image (Schneider et al. 2004). We have demonstrated previously that differences in lesion composition can be identified accurately with this analysis (Schneider et al. 2004). When differences were identified, quantitative analysis was performed as described later.
Quantitative Analysis
The processed images were opened in MetaMorph Image Analysis software (Universal Imaging, West Chester, PA) and analyzed quantitatively as previously described (Wadsworth et al. 2002). Measurements were logged onto a Microsoft Excel spreadsheet (MicroSoft; Redmond, WA) for later comparison.
Statistical Analysis
Values were expressed as means ± SD. Significances of differences were assessed with the use of Student's t-tests.
Results
Atherosclerotic Lesions in ApoE−/- Mice Compared with IRS2+/- ApoE−/- Mice
The percentage of the aortic intima that comprised atheroma (Figure 1) was characterized in IRS2+/- ApoE−/- mice and compared with that in ApoE−/- mice. The results were based on analysis of all of the serial sections in the 300-μm regions of the proximal aortas as previously described (Taatjes et al. 2000; Wadsworth et al. 2002). Thus, the cross-sectional area measurements were integrated to yield an estimate of the entire volume of the atheroma in the neointima in the 300-μm region of aorta in each animal. The percentage of the aortic tunica intima occupied by atheroma was greater in the IRS2+/- ApoE−/- mice than in the ApoE−/- mice. Thus, IRS+/- ApoE−/- mice with genetically intensified insulin resistance exhibited lesions occupying 79 ± 3% of the intima, compared with 68 ± 8% in ApoE−/- mice (n=5 in each group, p=0.018). Despite the increase in extent of the tunica intima that was occupied by atheroma, the composition of lesions with respect to percentage of lipid content, cellularity, and collagen content was similar in the two groups (Figure 1). Figure 2 shows representative images used in characterizing the lesions.
Effects of Pioglitazone on Atherosclerotic Lesions in IRS2+/- ApoE−/- Mice
The percentage of the aortic intima in the 300-μm region studied that was occupied by atheroma was similar in IRS2+/- ApoE−/- mice treated with pioglitazone and in IRS2+/- ApoE−/- mice treated with peanut butter alone (control). In 20-week-old mice, the extent of atheroma averaged 74 ± 5% (n=5) and was not significantly different from the percentage in control animals given vehicle alone (70 ± 7%, n=5).
Qualitative analysis of the atheroma from 10-week-old mice demonstrated similar composition both with respect to cellularity (pioglitazone = 2.7 ± 0.4; control = 3.1 ± 0.3) and lipid (pioglitazone = 3.7 ± 0.3; control = 3.4 ± 0.4). By contrast, qualitative analysis of atheroma from 20-week-old mice demonstrated less neointimal lipid (pioglitazone = 2.3 ± 0.4; control = 3.8 ± 0.3, p<0.001) and a trend toward decreased cellularity (pioglitazone = 3.0 ± 0.5; control = 3.5 ± 0.5, p=ns).
Because of the observed qualitative differences, quantitative analysis was performed on atheroma from 20-week-old mice (Figure 3). The mice treated with pioglitazone exhibited significantly less neointimal lipid in the atheroma than control mice (pioglitazone = 30 ± 7%; control = 50 ± 6%, n = 5 in each group, p = 0.005) (Figure 4). Furthermore, the IRS2+/- ApoE−/- mice treated with pioglitazone exhibited decreased neointimal cellularity (19 ± 3%) compared with the IRS2+/- ApoE−/- peanut butter control group (24 ± 1%, p=0.018) (Figure 5). Neointimal collagen content was similar in the two groups.
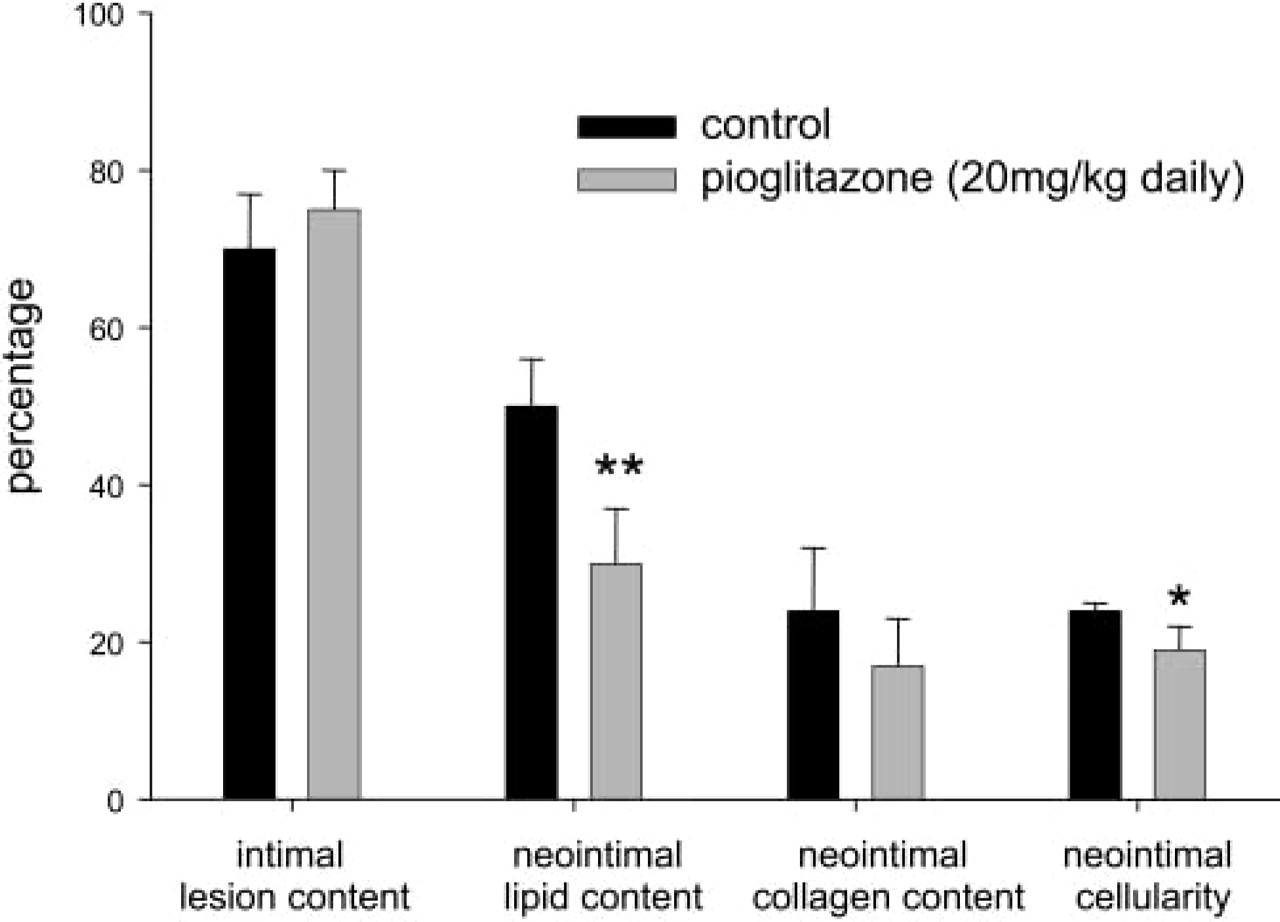
Effects of pioglitazone on features of the neointima in hyperlipidemic mice with intensified insulin resistance (IRS2+/- ApoE−/- mice). ∗,p=0.018; ∗∗,p=0.005.
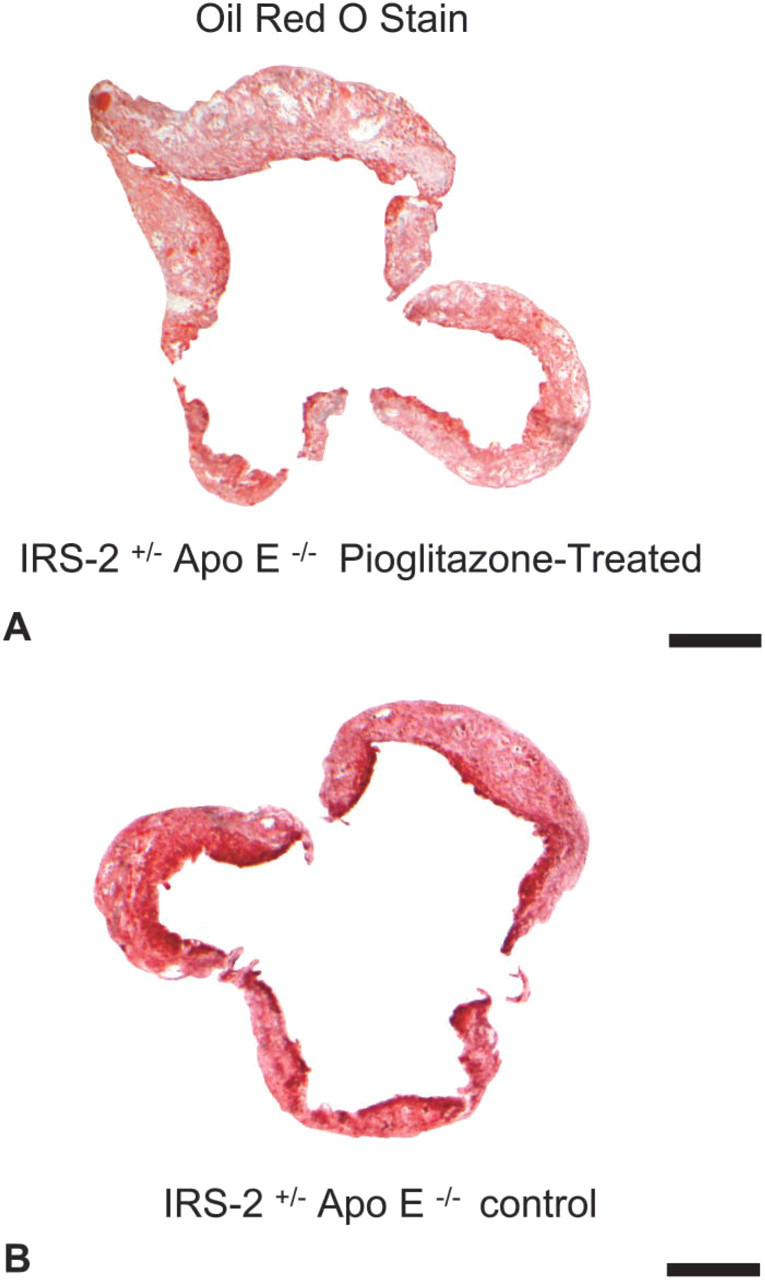
Representative images (transmitted light, bright field microscopy images captured with X4 objective lens) of oil red O–stained aorta; 10-μm-thick sections delineate lipid composition. Surrounding cardiac tissue, including media, has been cropped away with the use of Adobe Photoshop to facilitate quantitative analysis. The percentage of lipid was determined as the fraction of the total area of the atheroma in the neointima (μm2) occupied by lipid (μm2). (
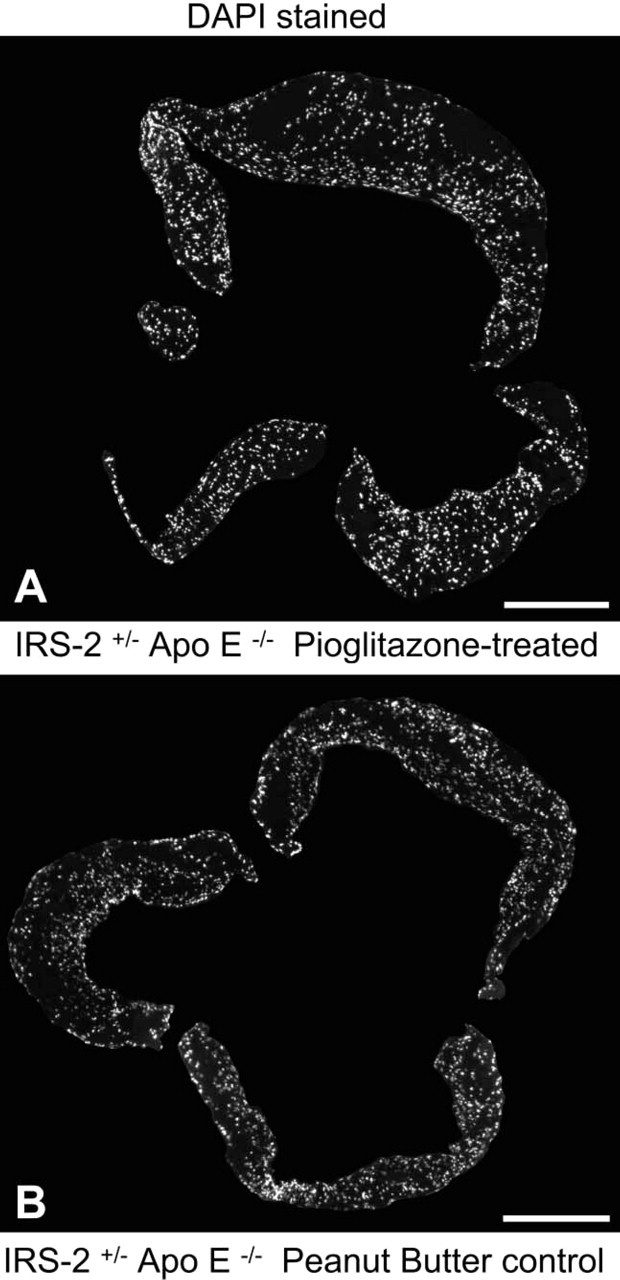
Cellularity in atherosclerotic lesions demonstrated by staining 10-μm-thick cryostat sections with DAPI. Images of the entire aorta in cross-section were created by digitally stitching together several individual images from a vessel. Images were captured with epifluorescence microscopy at a 10X magnification. (
Discussion
Our results demonstrate that genetically induced intensification of insulin resistance conferred by heterozygous deficiency of IRS2 increases the extent of atherosclerosis in ApoE−/- mice. We have shown previously that ApoE−/- mice exhibit insulin resistance of modest extent (Cefalu et al. 2004). Heterozygous deficiency in IRS2 is known to induce insulin resistance (Withers et al. 1999; Previs et al. 2000). The present results are consistent with the hypothesis that intensification of insulin resistance can accelerate plaque formation in the setting of atherogenesis. Our results in IRS2+/- ApoE−/- mice are consistent with observations that have been made in patients with type 2 diabetes who are insulin resistant for years before the onset of frank diabetes and throughout much of its course. Coronary angiographic studies have shown that the extent of vascular disease is increased markedly in patients presenting with diabetes (Natali et al. 2000; Ledru et al. 2001).
To determine whether attenuation of insulin resistance affected the nature of atheroma, we studied IRS2+/- ApoE−/- mice in which the intensity of insulin resistance is greater than in ApoE−/- mice without heterozygous deficiency of IRS2. Atheroma in mice treated with pioglitazone daily for 6 and 16 weeks were compared with those from mice given vehicle alone. Pioglitazone did not change the characteristics of lesions in 10-week-old mice fed drug for 6 weeks. Although the extent of vascular disease was not altered in 20-week-old mice, the amount of lipid in the neointima was decreased. These changes are consistent with a potential decrease in the vulnerability of atherosclerotic plaques to rupture (Davies and Thomas 1985; Falk et al. 1995).
The decreased neointimal cellularity and lipid content that we observed is consistent with results of others who treated ApoE−/- mice with thiazolidinediones. Phillips and coworkers (2003) found that ApoE−/- mice fed a high-fat diet and treated with rosiglitazone after exogenous injury to a carotid artery exhibited a reduction in the number of macrophages in the neointima. Troglitazone has been shown to reduce the size of atherosclerotic lesions in the aorta of ApoE−/- mice fed a high-fat diet (Chen et al. 2001; Collins et al. 2001). Previously, however, we did not observe an effect of troglitazone or rosiglitazone on lesion size or composition in ApoE−/- mice fed a high-fat diet (Cefalu et al. 2004). This difference may reflect differences in the duration and intensity of dyslipidemia in the two studies, gender-related differences, or other differences in experimental design. The present study differed from our previous work with respect to the characterization of the effect of pioglitazone on plaque composition in IRS2+/- ApoE−/- mice rather than ApoE−/- mice and the known greater affect of pioglitazone on lipid metabolism compared with rosiglitazone and troglitazone (Kipnes et al. 2001; Khan et al. 2002; Derosa et al. 2004; Peters Harmel et al. 2004).
Thiazolidinediones activate peroxisome proliferator activated receptor gamma (PPARγ) that increases expression of CD36, a scavenger receptor commonly found on the surface of monocyte-derived macrophages (Li et al. 2000; Han et al. 2003). CD36 enhances uptake of LDL and particularly that of oxidized LDL. Accordingly, increased expression of CD36 would be anticipated to increase foam cell formation and thereby increase the lipid content of atherosclerotic lesions (Li et al. 2000; Han et al. 2003; Plutzky 2003). In contrast to rosiglitazone, pioglitazone exhibits a more beneficial effect on the concentration of lipid components in blood (Kipnes et al. 2001; Khan et al. 2002; Peters Harmel et al. 2004). Thus, our results suggest that the effect of pioglitazone on the concentration of lipids in blood is the predominant factor accounting for its influence on the composition of atherosclerotic plaques in intensely insulin-resistant, dyslipidemic, and atherogenic mice.
Treatment with pioglitazone decreased not only the lipid content but also neointimal cellularity in the IRS2+/- ApoE−/- mice. These results are consistent with the direct effect of glitazones on PPARγ that have been seen in vascular smooth muscle cells in culture. Glitazone-induced activation of PPARγ has been shown to decrease proliferation of VSMC (Goetze et al. 2000; Law et al. 2000; Bruemmer et al. 2003; de Dios et al. 2003). The decreased proliferation of VSMC may have contributed to the decreased neointimal cellularity we observed in the IRS2+/- ApoE−/- mice treated with pioglitazone. Subsequent studies are planned to characterize the effect of pioglitazone on specific cellular constituents of the neointima.
Conclusions
Consistent with coronary angiographic and ultrasonic observations made in patients with type 2 diabetes, genetically induced intensification of insulin resistance in genetically dyslipidemic, atherogenic, transgenic mice increases the extent of vascular disease. Furthermore, attenuation of insulin resistance by treatment with pioglitazone decreases the lipid and cellular content of atheroma and therefore may be capable of decreasing the risk of rupture of vulnerable atherosclerotic plaques in patients with insulin resistance.
Footnotes
Acknowledgements
This research was conducted with support from Takeda Pharmaceuticals North America, Inc. This work was supported in part by a Clinical Research Award from the American Diabetes Association (to BES).
We would like to thank Heidi Taatjes for excellent technical assistance.