
Abstract
Facial and calvarial bones form intramembranously without a cartilagenous model; however, cultured chick calvarial mesenchyme cells may differentiate into both osteoblasts and chondroblasts and, in rodents, small cartilages occasionally form at the sutures in vivo. Therefore, we wanted to investigate what factors regulate normal differentiation of calvarial mesenchymal cells directly into osteoblasts. In embryonic mouse heads and in cultured tissue explants, we analyzed the expression of selected transcription factors and extracellular matrix molecules associated with bone and cartilage development. Cartilage markers Sox9 and type II collagen were expressed in all craniofacial cartilages. In addition, Msx2 and type I collagen were expressed in sense capsule cartilages. We also observed that the undifferentiated calvarial mesenchyme and the osteogenic fronts in the jaw expressed Co∗∗∗l2A1. Moreover, we found that cultured mouse calvarial mesenchyme could develop into cartilage. Of the 49 explants that contained mesenchyme, intramembranous ossification occurred in 35%. Only cartilage formed in 4%, and both cartilage and bone formed in 4%. Our study confirms that calvarial mesenchyme, which normally gives rise to intramembranous bone, also has chondrogenic potential.
T
Head mesenchyme mostly derives from ectomesenchymal tissue of neural crest (NC) origin (Couly et al. 1993,1998), but some seems to be of mesodermal origin (Jiang et al. 2002). In mice, mesenchymal condensations at the sites of future bone appear at embryonic day (E) 10 and 11, but determinants of this early patterning are only partly understood. The NC cells obtain signals from neuroectoderm and from facial ectoderm both before and after entering the future sites of cartilage and bone formation (Schneider et al. 2001; Lee et al. 2001; Depew et al. 2002). The formation of skeletogenic condensations requires epithelial-mesenchymal interactions and is regulated by the cellular microenvironment. This regulation includes diffusible signal molecules, extracellular matrix molecules, their receptors, and cell adhesion molecules. For example, fibronectin, tenascin, syndecan, and neural cellular adhesion molecules are associated with the initiation and boundary setting of the condensations, whereas the proliferation of cells within them is modulated by diffusible signal molecules (reviewed by Hall and Miyake 2000).
Morphological studies of the developing head indicate that cells in the mesenchymal condensations are strictly regulated to form either cartilage or bone, and recent experimental studies suggest that the information required for this selection could reside in the neighboring epithelial structures. The picture seems to be more complicated, however. Characterization of “chondroid bone,” found in human mandible and in cranial sutural areas and differing from both bone and cartilage, suggests that an intermediate type of tissue exists (Goret-Nicaise et al. 1988). It has even been stated that in chick, calvarial development starts with deposition of such chondroid tissue at the sites of future frontal, parietal, and squamosal bones, and that bone is deposited on the external side of this primitive, mineralized desmocranium (Lengelé et al. 1990). Moreover, cultured mesenchymal cells from rat and chick calvariae can give rise to both chondroblasts and osteoblasts (Bellows et al. 1989; Wong and Tuan 1995), and cultured calvariae from calcium-deficient chick embryos form cartilage (Jacenko and Tuan 1995).
Sox9 is required for cartilage differentiation and expression of chondrocyte-specific genes that code for cartilage extracellular matrix components, including collagen types II, IX, and XI and aggrecan. In humans, dominant negative mutations in Sox9 lead to campomelic dysplasia, a severe dwarfism syndrome (reviewed by Lefebvre and de Crombrugghe 1998, de Crombrugghe et al. 2000). Cells in skeletal condensations express Runx2 (Cbfa1), a transcription factor required for bone development. It is critical for osteoblast differentiation and positively regulates gene expression of bone extracellular matrix components such as type I collagen, osteopontin, osteocalcin, and bone sialoprotein (Bsp). In the absence of Runx2, bones are lacking, but a cartilagenous skeleton forms. Heterozygosity for this mutation in mice and in humans leads to cleidocranial dysplasia. The skeletal phenotype includes short stature, hypoplastic clavicles, and delayed closure of calvarial sutures and fontanels (reviewed by Ducy 2000). Transcription factor Msx2 is also required for normal bone development. Mice overexpressing Msx2 and humans with a gain-of-function mutation in Msx2 show craniosynostosis (premature fusion of the calvarial sutures; reviewed by Wilkie 1997), whereas Msx2 deficiency leads to defective frontal bone ossification, resulting in a persistent anterior fontanel in mice and parietal foramina in humans (Satokata et al. 2000).
Most of the studies suggesting a chondrogenic potential for the calvarial mesenchymal cells have been performed in chick. In mice, formation of the calvarial bones starts from mesenchymal condensations dorsally close to the endochondrally formed caudal part of the interparietal bone and bilaterally above the eye region. After initiation, the forming bone spreads, in vivo and in vitro, to cover the brain until the bony fronts meet in sutures, where their fusion is strictly regulated (Kim et al. 1998). Calvarial bone development in the mammalian head has widely been used as an example of intramembranous ossification.
Our purpose was to study the potential of the calvarial mesenchymal condensations to form bone and cartilage and its molecular basis. We hypothesized, based on the existing knowledge, that calvarial mesenchyme initially has potential to differentiate into either chondroblasts or osteoblasts. Therefore, we analyzed in embryonic mouse heads the expression of transcription factors Sox9, Msx2, and Runx2 as well as cartilage and bone extracellular matrix molecules Col1A1, Col2A1, Bsp, types I, II, III, and V collagen, laminin, fibronectin, syndecan, and tenascin. These molecules have previously been associated with hard tissue formation, but their expression patterns have been less well described in the developing calvaria than in the endochondrally forming long bone. Therefore, we performed parallel analyses in limb as control. Moreover, we studied bone and cartilage formation in tissue explants comprising early embryonic mouse calvarial mesenchyme to test the hypotheses that (a) differentiation of calvarial mesenchyme into hard tissue-forming cells is dependent on paracrine signals from epithelial tissues and/or from the already formed bone, and (b) in the absence of such signals, the cells, already committed to hard tissue formation, may take a more primitive, chondrogenic pathway.
Materials and Methods
Preparation of Tissue
We used mouse embryos (NMRI × NMRI), whose age was assessed by the appearance of the vaginal plug (day 0) and confirmed by morphological criteria. The heads from E9 to E17 embryos and front limbs from E15 and E17 embryos were dissected in Dulbecco's PBS under a stereomicroscope, fixed in 4% paraformaldehyde overnight at 4C, dehydrated in an ascending ethanol series, and routinely embedded in paraffin. Serial sections, 5-μm thick, were cut in a frontal plane, mounted on silane-coated slides, and stored at 4C until use. The small heads were cut throughout, and, starting from E13, the part anterior to developing incisors and the posterior third of the head were excluded. Selected slides were stained with hematoxylin and eosin (H&E) for morphological analysis or were subjected to radioactive in situ hybridization or immunohistochemistry. These analyses were performed at different anteroposterior levels.
In Situ Hybridization
The preparation of the following RNA probes has previously been described: Msx2 (Jowett et al. 1993), proα1(I) collagen and proα1(II) collagen (Metsäranta et al. 1991), and Bsp (Rice et al. 2000). The 500-bp Sox9 plasmid and the 600-bp 5’ PstI-HindIII fragment of murine Cbfa1 (Runx2) were kindly provided by Peter Koopman and Seppo Vainio, and by Toshihisa Komori, respectively. Radioactive in situ hybridization, with appropriate sense controls, was carried out under high stringency, as previously described (Vainio et al. 1991).
Immunoperoxidase Staining
We used polyclonal antibodies raised in rabbits against the C-terminal telopeptide region (included in the triple-helical collagen molecule) of human type I collagen (Risteli et al. 1993), the N-terminal propeptide of human type III procollagen (Niemelä et al. 1985), and human type V collagen (Odermatt et al. 1983); all of these antibodies were gifts from Leila Risteli and Juha Risteli. Polyclonal antibodies raised in goat against mouse type II collagen were from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA). Rabbit antiserum to rat tenascin (Mackie et al. 1988) was a gift of Eleanor Mackie. A monoclonal antibody raised against syndecan from epithelial cells of mouse mammary glands (Jalkanen et al. 1985) was from Markku Jalkanen. Antti Vaheri provided the rabbit antisera against human plasma fibronectin (Vuento and Vaheri 1979). Rabbit antiserum to rat laminin was from Telios Pharmaceuticals, Inc. (San Diego, CA). Control sections were treated with corresponding dilutions of normal rabbit serum, normal rabbit IgG, and normal goat IgG. The sections were immunostained with Vectastain ABC Elite kit (Vector Laboratories, Burlingame, CA), and the precipitate was stained with 3-amino-9-ethylcarbazole dissolved in N,N-dimethylformamide, as described before (Lukinmaa and Waltimo 1992). Triton-X (0.01–0.1%) was added to the blocking solution (5% normal serum from the species in which the secondary antibodies were raised) as well as to the primary and secondary antibodies. Before immunostaining, selected sections were pretreated with hyaluronidase (5000 U/ml PBS, pH 6.0) for 30 min at 37C and/or autoclaved in 0.1% sodium citrate, pH 6.0, for 20 min at 120C.
Tissue Recombinant Studies
Mouse embryos, aged E12.5 to E14, were dissected in Dulbecco's PBS (Gibco/Invitrogen; Paisley, Scotland, UK) under a stereomicroscope. A piece of approximately 1 mm2 was sectioned with injection needles from selected areas of the head (Figure 2A), and four tissue layers (surface epithelium, skeletogenic mesenchyme, leptomeningeal layer, and the developing brain) were identified (Figure 3). Selected layers, separated mechanically without any enzyme pretreatment, were cultured alone or in different combinations on Nuclepore filters (Whatman; Maidstone, UK) in Trowell-type organ cultures at the medium-gas interface for 8 to 21 days at 37C. The medium was changed every third day and contained 10% normal calf serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 100 μg/ml ascorbic acid in Dulbecco's modified Eagle's medium. Thereafter, explants were fixed in ethanol and routinely stained with Alcian blue and alizarin red to detect cartilage and bone formation, respectively. Alternatively, they were fixed for 1 min in cold methanol and then with 4% paraformaldehyde, dehydrated, embedded in paraffin and serially sectioned. Adjacent sections were mounted on slides, routinely stained with H&E for morphological analysis, or subjected to immunohistochemistry or in situ hybridization.
Some tissue explants were transplanted under kidney capsules of adult male nude mice (Harlan; Horst, The Netherlands), anaesthetized with tribromoethanol, and left for 10 to 23 days. The mice were then killed, and the tissue transplants were dissected out, stained with Alcian blue and alizarin red, or prepared into sections and treated as described previously. All experiments were approved by the Institutional Ethical Committee of Animal Experiments of Helsinki University, Finland.
Results
Expression of Bone and Cartilage Extracellular Matrix Molecules
To test markers for bone and cartilage development, we analyzed the expression patterns of a number of extracellular matrix molecules in our embryonic mouse material. These markers were used later to analyze the results of the tissue recombinant studies. We also looked at the gene and protein expression in different parts of the developing head and limb to find differences in the composition of the bones and cartilages that could be related to their different formation patterns and developmental role.
As expected, type I collagen expression at mRNA and protein levels was widespread but was most abundant in the matrix of both endochondral and intramembranous bones (Figures 1B, 1D–1J). It was also detected in the preosteogenic calvarial and jaw mesenchyme (Figures 1A–1D) and in the dental papilla (Figure 1D). We observed notable differences in type I collagen expression in the different cartilages: in the developing long bones its expression was confirmed in peripheral cartilage cells in the zones of high proliferative activity (not shown), and uniform expression was found in the dorsal cartilages of the braincase wall (Figures 1G and 1H). No reactivity was observed in Meckel's cartilage (Figures 2I–2J).
Antibodies recognizing the amino terminal propeptide of type III collagen yielded an expected positive reaction in all connective tissues. In the head, it was weaker in the cartilage than in the bone (not shown). Type V collagen reactivity was relatively weak but uniform in cartilage (Figures 1K–1L). Syndecan and fibronectin were observed in both bone and cartilage. As expected, tenascin expression followed the pattern of type I collagen in cartilage and bone, and was almost absent in other tissues. Notably, the staining results in the different cephalic cartilages with all these antibodies were uniform (not shown).
As documented earlier, Bsp was expressed in all areas that were morphologically identified as bone. In the limb, reactivity was limited to the ossifying central core areas of long bones and to a nearby layer beneath the perichondrium, making it the most specific of our markers for bone formation. We expected to find a similar correlation between cartilage morphology and type II collagen. We found this correlation at the level of protein expression, but Col2A1 was also detected in the calvarial preosteogenic mesenchyme (Figure 1M) and at the periphery of forming jawbones (Figures 1P and 1R), although not in the dental mesenchyme (Figure 1Q). The morphology of the developing calvaria is best visualized in the H&E-stained section (Figures 1S and 1T).
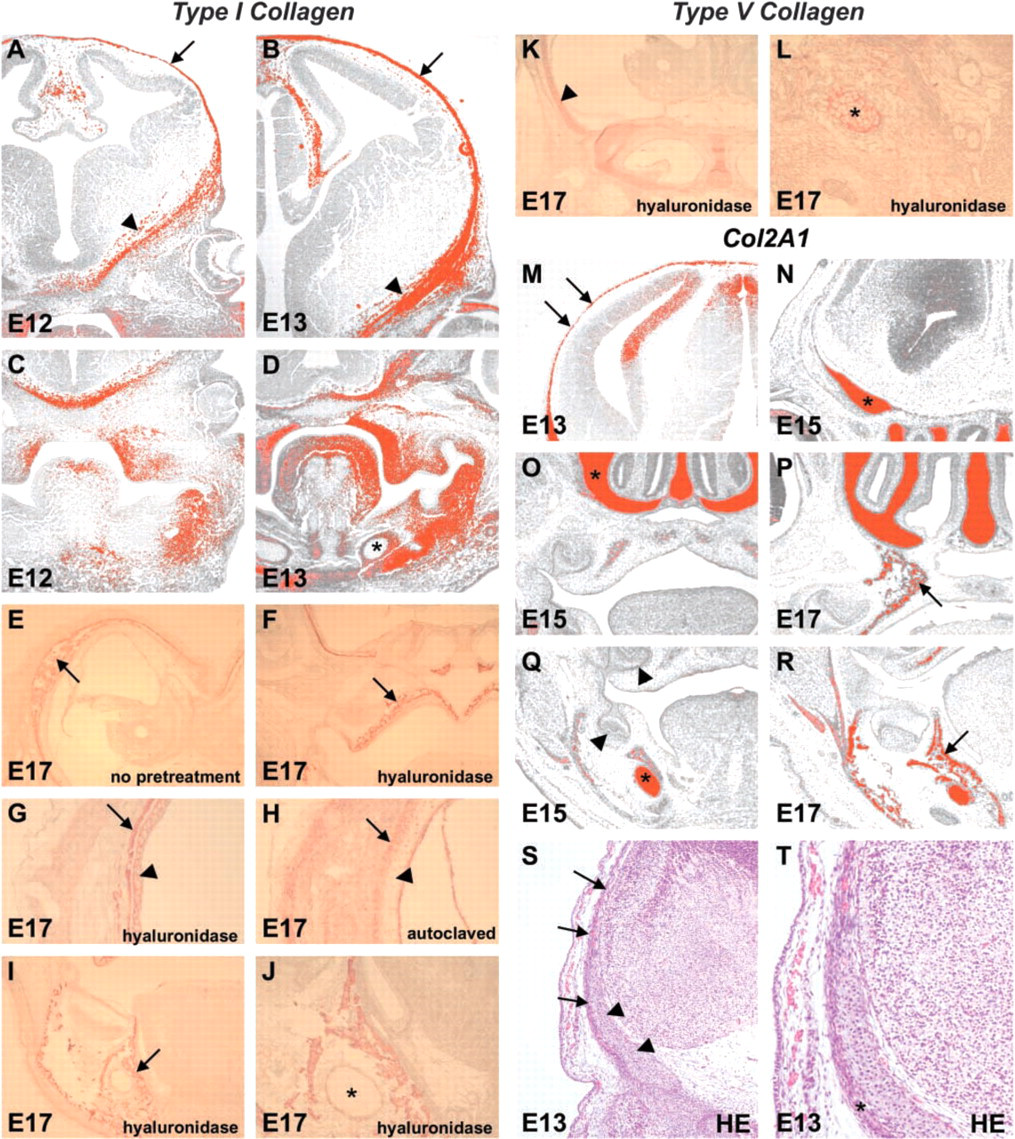
Expression of types I, II, and V collagen in embryonic craniofacial tissues. (
Expression of the Transcription Factors Runx2, Msx2, and Sox9 in Relation to Skeletogenic Development
At E12.5, bone started to form in the lower jaw (laterally to Meckel's cartilage and the nerve-blood vessel bundle), lateral and inferior to the nasal cartilage, and lateral to the primary wall of the braincase. Before bone matrix was laid, the mesenchyme became condensed at these sites and started to express Runx2 at E10.5. Similarly, in the ossifying long bones Runx2 expression preceded the advancement of bone formation. Consequently, Runx2 expression was used to define the osteogenic part of the mesenchyme (Figures 2A and 2B). Msx2 expression was more widespread, but a slight increase in reactivity was seen in the ossifying areas. Moreover, we found that type I collagen and Msx2 were coexpressed in the developing bone and the dorsal cartilages of the primary braincase wall (Figure 2F), but neither was found in Meckel's cartilage (Figures 2I and 2J). Msx2 expression was strong throughout the calvarial skeletogenic mesenchyme (Figures 2C–2F) and in the dental mesenchyme (Figure 2J), but only weak or moderate levels of expression were seen in other bone-forming areas and in the neuroepithelium (Figures 2C–2J).
Sox9 was localized in all morphologically identified cartilages, including the primary braincase, Meckel's, and nasal cartilages. In addition, Sox9 was expressed in the neuroepithelium (Figures 2K–2P). In the limb, Sox9 was expressed evenly in all cartilages, and it gradually disappeared from the ossifying areas. Col2A1 expression was identical to that of Sox9. In all cartilages the increased expression of Col2A1 was associated with Sox9 expression. This association was, however, less clear in the (skeletogenic) mesenchyme in the area of future calvarial bone and at the sites of the maxillary and mandibular bone expansion.
Differentiation of Calvarial Mesenchyme in Tissue Recombination Explants
We first made sure that the culture conditions allowed skeletogenic differentiation by culturing the thickened mesenchyme above the eye from two E12.5 heads. This mesenchyme was positive for Runx2 and was therefore expected to undergo intramembranous ossification under permissive conditions. Brain tissue was removed, but surface epithelium was not. After 15 days of culture, we detected hard tissue separated from the epithelium by a clear zone. Central parts of the hard tissue islets were alizarin red-positive, and ossification was confirmed in sectional in situ hybridization by strong Bsp expression. Notably, a zone surrounding the bony plates stained with Alcian blue and showed some Col2A1 expression. Col2A1 was also detected in some osteocytes within bony lacunae close to the sites of bone expansion (not shown).
We defined the area of the future calvaria, where no known signs of bone formation could yet be observed, based on our morphological and gene expression studies (Figures 2A, 2B, and 2Q–2T). At E12.5 to E14, the central top area of the embryonic mouse head was devoid of markers of hard tissue differentiation and therefore was used in the in vitro experiments. It turned out to be technically possible to cut minute pieces of tissue from the area next to the future anterior fontanel and to separate them into four delicate tissue layers (surface epithelium, skeletogenic mesenchyme, leptomeningeal sheet, and brain) (Figure 3). When these layers were cultured alone or in different combinations, it became obvious that hard tissue formed only within the mesenchyme. When the epithelium was cultured alone, it showed hyperkeratinization, and no bone or cartilage developed subepithelially. The leptomeningeal sheet covering the brain was thin, highly vascularized, and, when cultured alone, formed a single layer of cubical cells. In explants that contained parts of the brain, neuronal overgrowth occurred unless the leptomeninges had been left in place. Blood vessels continued to proliferate between the leptomeningeal sheet and mesenchyme (not shown).
Hard tissue formation was analyzed by Alcian blue and alizarin red staining of whole mounts (Figures 4A–4D) and by routine histological and molecular analyses of transverse serial sections (Figures 4E–4J). When the explant contained surface epithelium and mesenchyme (n=23), bone developed in two explants, and cartilage in other two. In the absence of epithelium (n=14), mesenchyme differentiated into bone in 50% of the explants and into separate islands of cartilage and bone in one. In some of the explants, where formation of plates of bone but not cartilage was evident, the alizarin red-positive bony particles were surrounded by a narrow bluish rim, indicating cartilage extracellular matrix (Figure 4C). Areas that were morphologically identified as bone always expressed type I collagen and Bsp (Figure 4G, I). The bone either expressed no Col2A1, or Col2A1 was detected around the edges of the bony plates and in some osteocytes within the peripheral bone (Figures 4H and 4J). All areas with a cartilagenous phenotype uniformly expressed Sox9 and Col2A1; the probes for type I collagen showed no reactivity different from the dorsal cartilages of the primary braincase wall. In one explant with cartilagenous plates of uniform structure, weak expression of Runx2 and a stronger expression of Col1A1 were evident in selected areas of the perichondrium, suggesting appositional bone formation.
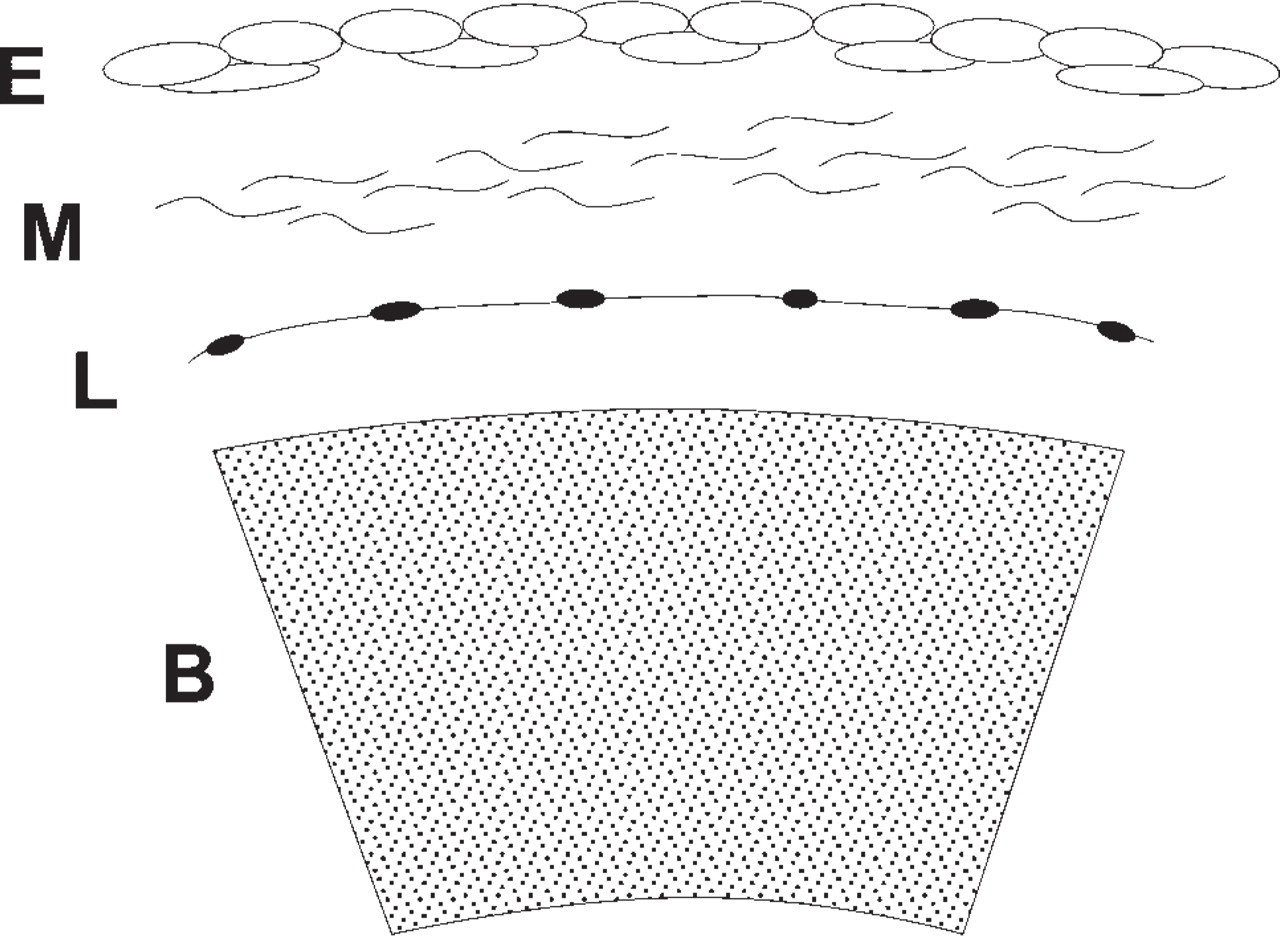
The tissue layers cultured in isolation or at different combinations. B = developing brain, E = surface epithelium, L = leptomeningeal sheet, M = mesenchyme.
To ensure conditions as close to the in vivo situation as possible, some E13 and E14 tissue explants were transplanted under the kidney capsules of adult male nude mice. In both groups, comprising of epithelium and mesenchyme combined (n=6) and mesenchyme alone (n=6), intramembranous bone formed in 50% of the explants. One explant additionally showed extensive plates of cartilage, which, surprisingly, seemed to undergo endochondral ossification (Figure 4K). Signal for Col2A1 was uniform in the cartilagenous end (Figure 4L), whereas the opposite end, which had transformed into bone, expressed Bsp (Figures 4M and N). Type II collagen protein was present throughout the cartilage. Antibodies to type I collagen gave a strong immunoreaction in the perichondrium and, notably, also in the chondrocyte cytoplasm in autoclaved sections, reminiscent of the type I collagen staining pattern seen in the cartilages of sense capsules in vivo (not shown).
In the combined experiments, intramembranous ossification occurred in 35% of the 49 explants that contained mesenchyme. Only cartilage formed in 4%, and both cartilage and bone formed in 4%. No common nominators were found for the sections that failed to form hard tissue or for those that developed bone or cartilage.
Discussion
Alterations in the Balance of Msx2 and Sox9 Expression Are Associated with Different Patterns of Hard Tissue Formation
Typical of intramembranous ossification both in vivo and in vitro were the absence of cartilage morphology but expression of Col2A1 at the edges of the developing bone. This pattern occurred in the presence of Runx2, a low level of Sox9, and a moderate level of Msx2. In the cephalic cartilages, Sox9 was uniformly expressed, whereas Msx2 was not detected in most of them. We observed, however, that Msx2 and type I collagen were both expressed in the lateral wall of the braincase, similar to the peripheral cartilage zone in developing long bones. We suggest that Msx2 and type I collagen might be expressed in rapidly proliferating chondroblasts. Msx2 expression in calvarial mesenchyme indicates a regulatory role in calvarial skeletogenesis and fits well with the mutant phenotypes.
Dental mesenchyme, which gives rise to dentin and periodontal tissue and possibly also to alveolar bone, displayed strong Msx2 expression but no Col2A1. This expression is probably regulated through early signaling between oral epithelium and the underlying mesenchyme, a reciprocal interaction that is crucial for tooth morphogenesis and cellular differentiation (Thesleff and Nieminen 2001). Tissue recombination studies, in line with our own unpublished observations, have shown that dental mesenchyme lacks chondrogenic potential (Kollar and Baird 1970). Moreover, periodontal ligament cells that line tooth roots have not been shown to form cartilage as a reparative tissue after tissue damage (e.g., damage caused by excessive orthodontic force). Nested expression of Dlx homeobox genes patterns the maxillary and mandibular arches, including teeth (Depew et al. 2002). Knocking out Dlx1 and Dlx2 leads to agenesis of certain teeth and to ectopic cartilage formation in the position of the missing maxillary molar teeth. This process occurs along with other aberrant features in the skull hard tissues that resemble those in phylogenetically more primitive vertebrates (Qiu et al. 1997). It seems, therefore, that jaw mesenchyme looses its potential for cartilage development when it acquires potential for tooth formation through epithelial induction.
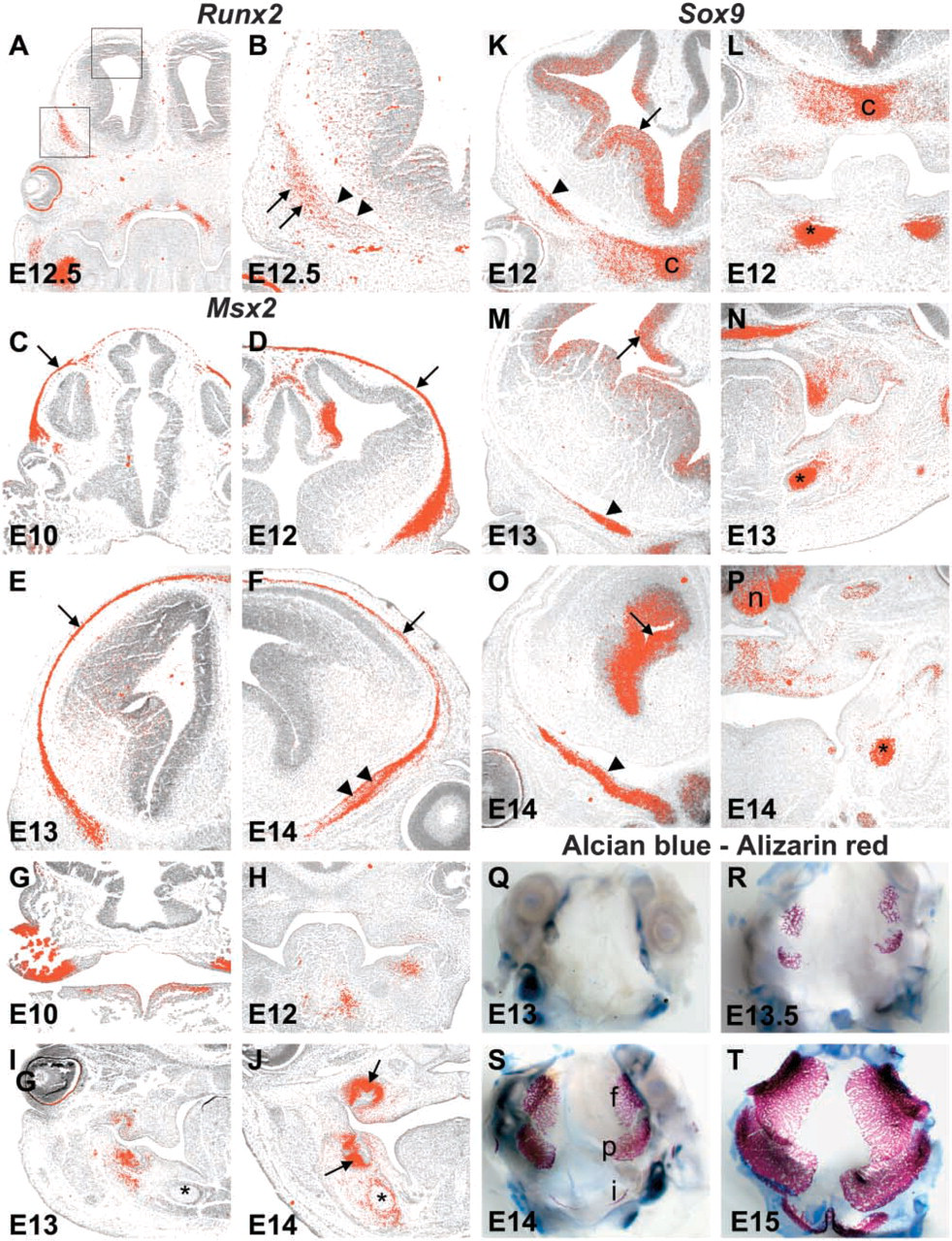
Expression of transcription factors Runx2, Msx2, and Sox9, and calvarial cartilage and bone development. (
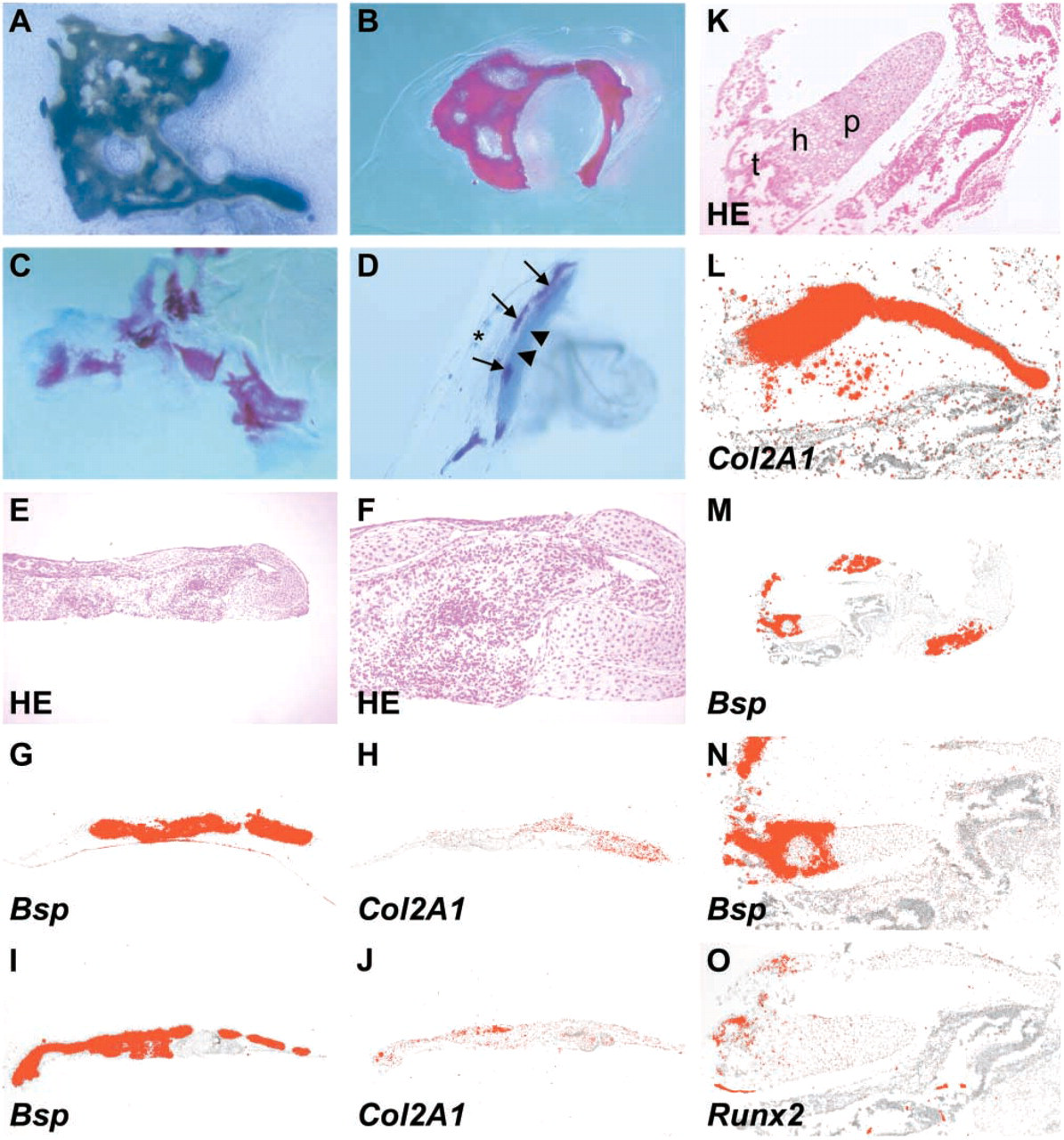
Bone and cartilage formation in the cultured calvarial mesenchyme. (
In Figure 5, we illustrate how the model of hard tissue development in the embryonic skull might be associated with the mutual balance of Sox9 and Msx2. In the absence of direct mechanistic data, this proposal remains a preliminary hypothesis. It is, however, in line with experimental work indicating that chondrogenesis only takes place in areas where Sox9 expression is high relative to that of Msx2 (Semba et al. 2000). In a subpopulation of migrating cranial NC cells, inhibited Msx2 function leads to accelerated chondrogenesis (Takahashi et al. 2001). Hence, Msx2 as a transcriptional repressor may inhibit Sox9-induced cartilage formation.
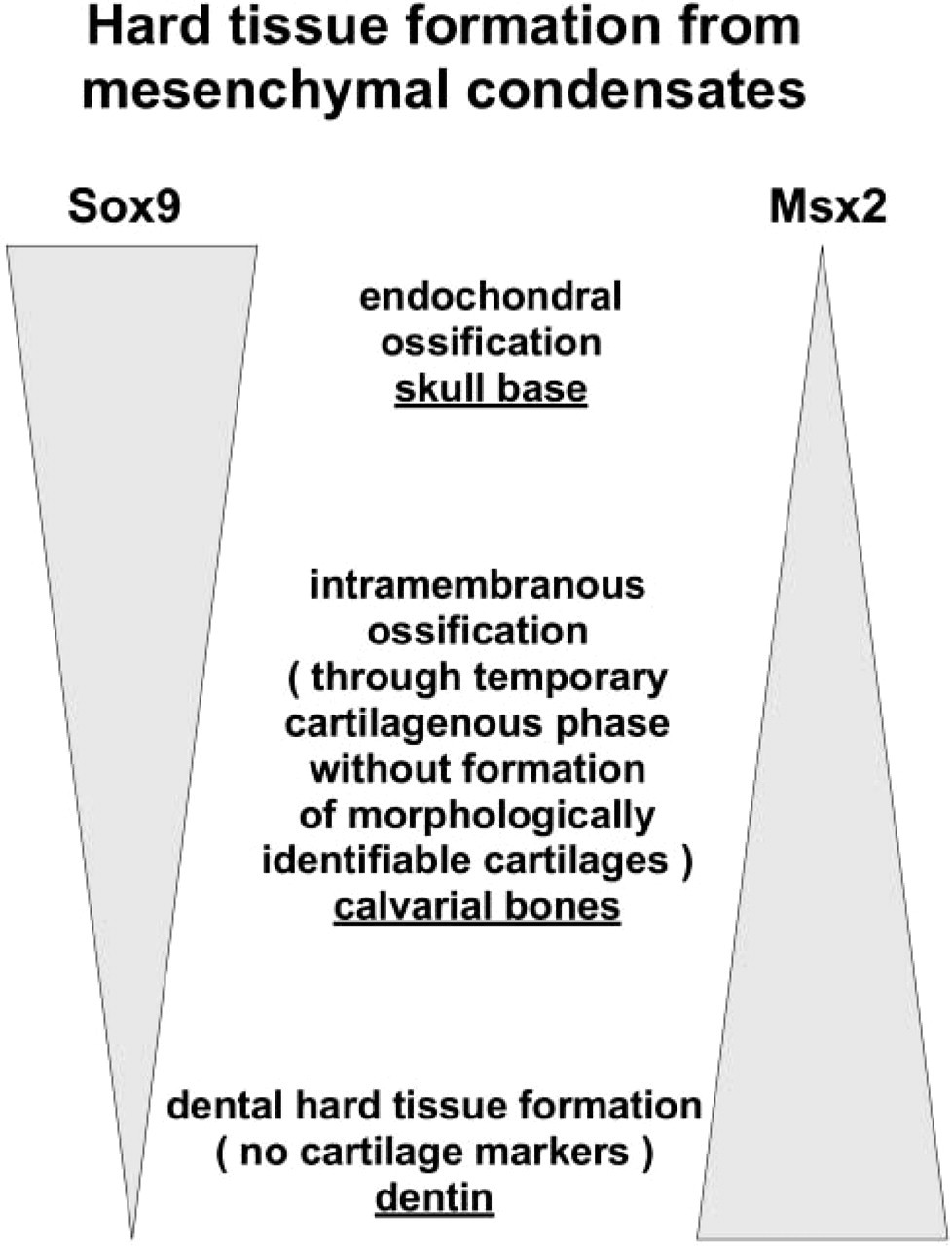
Hypothesized association of the balance between the transcription factors Msx2 and Sox9 and different models of hard tissue formation.
Intramembranous Ossification Occurs in Isolated Calvarial Mesenchyme
In our tissue recombination studies, calvarial mesenchyme at E13 and E14 retained its capacity to develop into intramembranous bone. When the mesenchyme, surface epithelium, and neuroepithelial tissues were cultured in isolation or at various combinations, only mesenchyme gave rise to hard tissue and did so independently of the presence of the neighboring surface and neural epithelia. Of our 49 explants that contained mesenchyme, intramembranous ossification occurred in 35%, and chondrogenesis alone occurred in 4%. In another 4%, both cartilage and bone formed. The absence of bone or cartilage formation in 61% of the explants can reflect either lack of skeletogenic competence of the mesenchyme or inadequate culture conditions. We speculate that if the mesenchyme obtains the signal for skeletal differentiation before tissue preparation, the osteogenic or chondrogenic pathway is taken in response to microenvironmental factors that remain to be elucidated.
The presence of surface epithelium had a negative affect, if any, on skeletogenic differentiation. Cultured isolated premigratory NC cells from early chick embryo, however, do not differentiate into bone or cartilage but do so when combined with epithelium. This finding indicates that interaction of NC with epithelium is crucial for skeletal commitment (reviewed by Fang and Hall 1997). In our explants this permissive interaction may have already occurred. As occurs in vivo, a subepithelial connective tissue layer in our explants always separated the bones and cartilages from surface epithelium. The apparent negative regulatory effect of surface epithelium has been proposed to result from epithelial short-range inhibitory signals that counteract the bone-inductive signals from the same epithelium (Mallo et al. 2000).
Intramembranous Ossification May Take Place Through a Short Interphase During Which Genes Associated with Chondrogenesis Are Expressed
We observed that Col2A1 was expressed in vivo in the murine mesenchyme that underwent intramembranous bone formation, and that some of the ossified explants in vitro showed peripheral Alcian blue staining as well as Col2A1 reactivity. Transient expression of type II collagen mRNA and protein in developing craniofacial tissues other than cartilage has been reported previously and is hypothesized possibly to be associated with skeletal patterning (Thorogood 1988; Cheah et al. 1991; Wood et al. 1991). The finding that targeted inactivation of Col2A1 results in a skeleton with membranous and periosteal bone but no endochondral bone strongly challenges that idea (Li et al. 1995). Chondrocytes do provide morphogenic signals that selectively induce osteogenic differentiation of mesenchymal stem cells, but their identity remains to be elucidated (Gerstenfeld et al. 2002).
A capacity of calvarial cells to express cartilage markers was reported earlier in chick (Toma et al. 1997; Nah et al. 2000), and a transient chondrogenic phase has been suggested as part of the normal intramembranous pathway (Nah et al. 2000). Expression of cartilage markers in chick calvarial cells diminishes with age of the donor animal, however. Thus, it has been concluded that the osteogenic differentiation of the calvarial mesenchyme results from negative selection against chondrogenesis in the absence of a permissive or inductive environment (Toma et al. 1997). Nevertheless, chondrogenic potential is still present in the periosteum covering the intramembranous bones. This potential becomes evident during secondary cartilage formation and fracture healing. In the dura beneath rat calvaria, this chondrogenic potential has been shown to be restricted to the sutural area (Yu et al. 1997).
We interpret the Col2A1 expression as a marker of a stage when the mesenchymal cells have developed the potential to form hard tissue, but final selection for osteogenesis or chondrogenesis has not been completed. The function of the transient Col2A1 expression during intramembranous ossification is not understood, but it is notable that mice with mutated type II collagen gene show retarded ossification of intramembranously forming bones (Savontaus et al. 2004).
Calvarial Mesenchyme Has a Potential to Differentiate into Cartilage
Our experiments were the first to show that mouse calvarial mesenchyme, when cultured in Trowell-type tissue culture or under the kidney capsule, can give rise to extensive plates of cartilage. This finding was confirmed morphologically and by gene expression. Cartilage formation within the thin, undifferentiated calvarial mesenchyme in culture conditions reflects absence of the final osteoblastic determination of the tissue. It was proposed earlier that cells making up the mesenchymal condensations are determined only for skeletogenesis, and that determination for osteogenesis, chondrogenesis, or chondroid tissue formation occurs later and seems to be linked tightly to the influence of local or regional factors acting in their microenvironment (Hall 1983; Lengelé et al. 1990).
The finding that calvarial mesenchyme can form cartilage but does not do so under normal development leaves open the question whether chondrogenic capacity is fulfilled only under special, permissive conditions or occurs unless actively repressed. Previous work on chick calvaria suggests that calcium deficiency creates a microenvironment permissive for chondrogenic differentiation and that normally calcified matrix is repressive to chondrogenesis (Jacenko et al. 1995). The finding that cranial NC cells retain the capacity to differentiate into cartilage when transplanted ectopically to the trunk level supports site-specific regulation of cell differentiation (Couly et al. 1998). Moreover, the cranial NC cells that contribute to the formation of mesenchymal dental components, Meckel's cartilage, and mandibular bone are of common origin (Chai et al. 2000). In the skeletal condensations model, hard tissue formation seems to be associated with the mutual levels of Sox9 and Msx2. Downstream of Sox9 and Msx2, Runx2 expression marks the final selection of skeletogenic precursors for osteoblastic differentiation, whereas Col2A1 marks the absence of differentiation.
Footnotes
Acknowledgements
This work was supported by Academy of Finland (JW-S, IT).
We thank Jukka Jernvall and Han-Sung Jung for help with kidney capsule transplantation experiments and Oxana Arsenieva, Merja Mäkinen, Riikka Santalahti, and Ludmila Razzkozova for excellent technical help.