
Abstract
Murine arylamine N-acetyltransferase 2 (NAT2) is expressed in the developing heart and in the neural tube at the time of closure. Classically described as a xenobiotic metabolizing enzyme, there is increasing evidence for a distinct biological role for murine NAT2. We have characterized the expression of arylamine N-acetyltransferase 2 during cardiogenesis, mapping its expression in vivo, using a lacZ insertion deletion, and also in vitro, by measuring NAT2 enzyme activity. These findings show that cardiac Nat2 expression is both temporally and spatially regulated during development. In neonatal mice, cardiac Nat2 expression is most extensive in the central fibrous body and is evident in the atrioventricular valves and the valves of the great vessels. Whereas Nat2 expression is not detected in ventricular myocardial cells, Nat2 is strongly expressed in scattered cells in the region of the sinus node, the epicardium of the right atrial appendage, and in the pulmonary artery. Expression of active NAT2 protein is maximal when the developing heart attains the adult circulation pattern and moves from metabolizing glucose to fatty acids. NAT2 acetylating activity in cardiac tissue from Nat2−/- and Nat2+/- mice indicates a lack of compensating acetylating activity either from other acetylating enzymes or by NAT2 encoded by the wild-type Nat2 allele in Nat2+/- heterozygotes. The temporal and spatial control of murine Nat2 expression points to an endogenous role distinct from xenobiotic metabolism and indicates that Nat2 expression may be useful as a marker in cardiac development.
A
Mouse and human NAT genes exist as a cluster of polymorphic genes found at orthologous loci on chromosome 8 (Blum et al. 1990; Hickman et al. 1994; Matas et al. 1997; Fakis et al. 2000). Two of these gene products show distinct but overlapping substrate specificity in vitro, whereas in mice there is a third gene (Nat3) that encodes a protein of unknown substrate specificity. In humans, there is also a pseudogene (NATP1)(Blum et al. 1990). On the basis of substrate specificities and tissue distribution, human NAT1 (hNAT1) is judged to be the functional homolog of murine Nat2 (mNAT2) (Hein et al. 2000).
The tissue distribution of each isoform is characteristic; mNAT1/hNAT2 expression is largely confined to tissues classically described as responsible for xenobiotic metabolism (Chung et al. 1993), whereas that of mNAT2/hNAT1 is more widespread (Stanley et al. 1996; Rodrigues-Lima et al. 2003). Although many of the sites of mNAT2/hNAT1 expression are compatible with a xenobiotic role, in some tissues mNat2 expression is enhanced in specific cell types and particular developmental stages not generally associated with xenobiotic metabolism, including the developing heart and neural tube (Mitchell et al. 1999; Cornish et al. 2003).
Mice lacking both Nat1 and Nat2 genes appear aphenotypic (Cornish et al. 2003; Sugamori et al. 2003), like null mutants of other xenobiotic metabolizing enzymes (Gonzalez 2002). However, the viability of NAT overexpression mutants is severely compromised (Sim et al. 2003), and human NAT1 allelic variation is also associated with developmental defects (Lammer et al. 2004). To allow us to distinguish the endogenous role of mNAT2 from that of its isoform, mNAT1, we have used mice specifically lacking mNAT2, generated by targeted insertion of lacZ into the Nat2 coding region (Cornish et al. 2003). Nat2 null mutants lack an obvious phenotype when reared under standard laboratory conditions even though the main site of NAT2 activity, the liver, lacks the ability to acetylate NAT2-specific substrates in vitro (Cornish et al. 2003).
The mammalian heart develops from lateral plate mesoderm via a simple tube whose spontaneous rhythmic contractions direct the fetal circulation from as early as the 10-somite stage. Development of the heart progresses through a continuum of morphological transformations involving looping of the tube heart, septation of the cardiac chambers, and differentiation of the multi-component conduction system (reviewed by Kirby 2002; Anderson et al. 2003). Integration with the cardiovascular centers of the autonomic nervous system, as well as some of the signals for the morphogenetic transformations, is accomplished by cells from the neural crest (Gitler et al. 2003; Hutson and Kirby 2003). To assess whether NAT would be a useful marker for cardiac development, we describe the expression pattern of mNat2 in the developing heart using histochemical and biochemical methods.
Materials and Methods
Except where stated otherwise, all chemicals were obtained from Sigma-Aldrich (Dorset, UK). All work involving animals was carried out according to the Animals (Scientific Procedures) Act of 1986 and was subject to local and national ethical review.
Nat2 Mutant Mice Maintenance and Breeding
The generation of a stable Nat2 knockout line of mice by targeted insertion of a lacZ-containing cassette has been described (Cornish et al. 2003). Essentially, a TAG3/IRES/ lacZ/loxP/neo/loxP reporter ablation cassette was inserted into the BglII site in the Nat2 coding region, an MCl-thymidine kinase dimer-negative selection cassette was appended, and the construct used to generate a null allele of mouse Nat2 by homologous recombination in 129/Ola embryonic stem cells. The mutant gene was maintained in the heterozygote state on a rapid acetylator (C57Bl/6) background by backcrossing Nat2+/- males to C57Bl/6 mice obtained from Harlan (Bicester, Oxfordshire, UK) as 8-week-old adults. Nat2−/-, Nat2+/-, and Nat2−/- animals used for analysis were generated by intercrossing. Routine genotyping of adult mice was by PCR using DNA isolated from ear clips as described in Cornish et al. (2003). Primers used for genotyping were mNat2–1, mNat2–10 (Estrada et al. 2000), and Neo-T as described in Cornish et al. (2003). Material for genotyping fetuses and neonates was obtained from yolk sac and tail tips, respectively. To obtain fetuses and neonatal pups, timed matings were set up, with noon on the day of the vaginal plug designated as embryonic day (E) 0.5. Pregnant dams and neonates were killed by cervical dislocation. Fetuses were dissected from the uterus into ice-cold 10 mM potassium phosphate, pH 7.5, 145 mM NaCl (PBS).
Preparation of Protein Samples
Tissues were dissected from adult animals immediately following cervical dislocation, trimmed, washed briefly in PBS, and snap frozen in liquid nitrogen. Tissues taken from adult mice were homogenized using an Ultraturax T25 (Janke and Kunkel; IKA Labortechnik, Germany) tissue homogenizer, and tissues taken from neonatal and fetal mice were homogenized in glass Dounce (Jencons; East Sussex, UK) homogenizers. Samples were prepared for Western blotting or acetylation activities as described previously (Smelt et al. 2000). Briefly, samples were centrifuged for 15 min at 12,000 rpm at 4C before recovering the supernatant and snap freezing in aliquots for storage in liquid nitrogen. Protein concentrations were determined as previously described (Smelt et al. 2000).
For each genotype and each sex, tissues were dissected from at least six adult C57Bl/6 mice weighing between 22 and 28 g, derived from intercrosses of backcross generations three and four. Tissue homogenates were assayed for their ability to acetylate para-aminobenzoic acid (pABA) essentially as described (Sinclair et al. 1998). Homogenates were diluted into 20 mM Tris-HCl, pH 7.5, 1 mM DTT, to a protein concentration such that the acetylation of substrate was linear over 10 min at 37C. The acetylation reaction was started by addition of acetyl CoA, and routinely the reaction was terminated by addition of ice-cold trichloro-acetic acid. An aliquot of liver homogenate made from pooled tissues taken from six adult male C57Bl/6 Nat2+/+ mice was included in each assay as an internal standard as previously described (Smelt et al. 1998). For Western blotting, cytoplasmic extracts from the hearts of six adult male mice of each Nat2 genotype were blotted after SDS-PAGE and probed with rabbit polyclonal antiserum 184, raised against the C-terminal dodecapeptide of human NAT1 conjugated to bovine serum albumin, as described (Stanley et al. 1997,1998; Cornish et al. 2003). The C-terminal dodecapeptide of human NAT1 is identical to that of mouse NAT2.
Whole-mount Staining for β-Galactosidase Activity
Hearts were isolated, trimmed, and fixed in formaldehyde/ glutaraldehyde buffer (2% formaldehyde, 0.2% gluteraldehyde, 0.02% NP40, 0.01% sodium desoxycholate in PBS) at room temperature for 10 min. After fixation, tissues were rinsed four times in PBS, then stained in the dark overnight at 37C in X-gal stain solution [5 mM K3Fe(CN)6, 5 mM K4Fe(CN)6, 1 mg/ml 5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-gal), 2 mM MgCl2, 0.02% NP-40, 0.01% sodium desoxycholate in 0.1 M phosphate buffer, pH 7.3, and 20 mM Tris-HCl, pH 7.3]. Tissues were then washed in PBS, dehydrated, and made transparent according to the protocol given by Rentschler et al. (2001). E11.5 fetuses were collected in ice-cold PBS, then stained exactly as above, and further dissected to isolate the hearts after staining and dehydration. To observe X-gal staining at a cellular level, tissue was fixed, stained with X-gal as above, dehydrated in a graded series of methanol, cleared with Histoclear (Lamb; Eastbourne, East Sussex, UK), and paraffin embedded. Sections were cut at 10-μm thickness, mounted on superfrost slides (BDH; VWR International, Lutterworth, Leicestershire, UK), heated to 60C for 1 hr, and stored at room temperature.
Results
NAT2 Acetylating Activity and Protein Levels Correlate with Nat2 Genotype
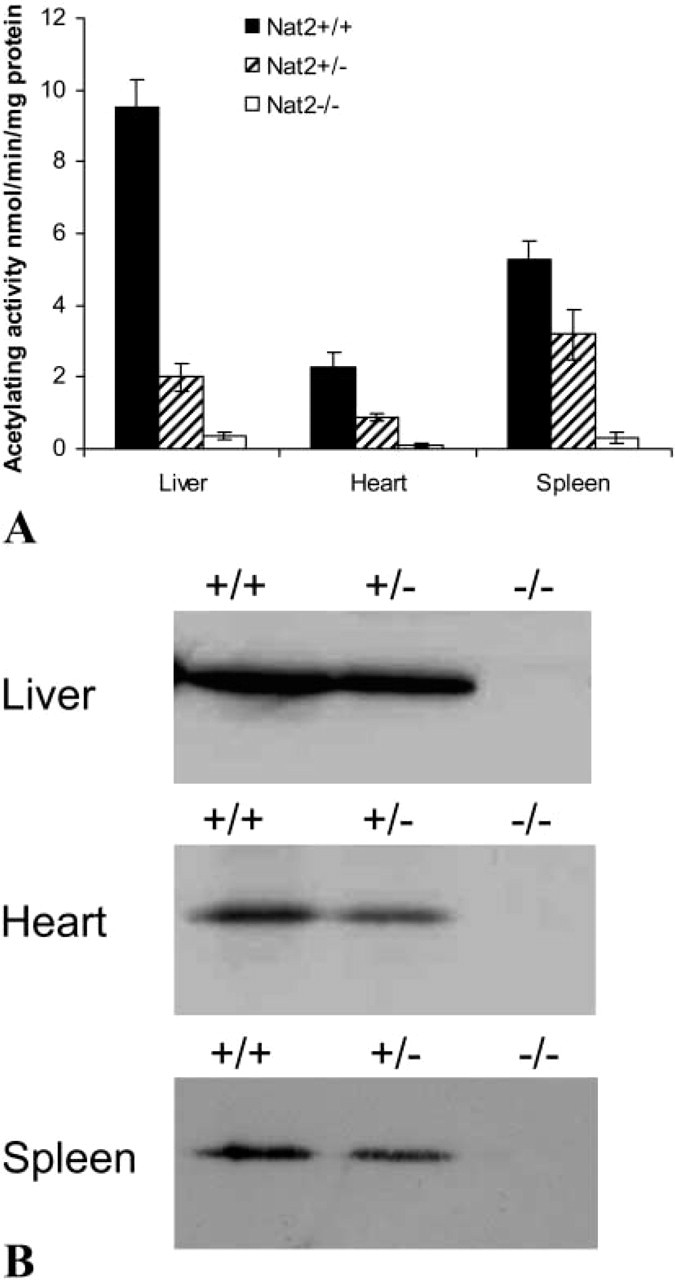
The expression of mouse Nat2 was investigated in mice of Nat2+/+, +/-, and −/- genotypes using a substrate known to be specific for mouse NAT2, pABA. Homogenates from tissues isolated from C57Bl/6 mice were assayed for their ability to acetylate pABA. Differences in the proportion of extracellular protein in different tissues (van den Hoff et al. 1997) were reduced by fractionating the tissue homogenates, removing intact cells and fibrous material, allowing more meaningful comparison of enzyme activity levels per unit mass.
Homogenates of heart tissue from adult Nat2+/+ animals show measurable pABA acetylating ability, at 25% that of liver homogenates from the same Nat2+/+ adults (Figure 1A). Although NAT2 activity in hearts from Nat2+/+ males (2.6 ± SD 0.4 nmol/min/mg protein; n=12) was greater than in hearts from Nat2+/+ females (2.1 ± SD 0.1 nmol/min/mg protein; n = 12), this difference was not statistically significant. Heart tissue shows good genotype/phenotype correlation comparing tissues from Nat2+/+ and Nat2+/- animals, indicating that the wild-type allele was not overexpressed to compensate for the null allele.
(
Specific substrates have been identified for mNAT1 and mNAT2 enzymes. pABA is a specific substrate for murine NAT2 and is not acetylated by murine NAT1 (Estrada et al. 2000). Hence, NAT1 activity does not contribute to the observed pABA acetylation. A NAT3-specific substrate has yet to be defined Kelly and Sim 1994; Fretland et al. 1997; Estrada-Rodgers et al. 1998). As Nat3 transcripts have been detected in the spleen (Boukouvala et al. 2002), it was of interest to establish whether the activity of NAT3 might be able to compensate for the loss of NAT2 function in spleens of heterozygotes or animals with the Nat2 genes ablated. The acetylation activity of the adult spleen was found to be 50% that of adult liver, with heterozygotes showing slightly higher than expected values; however, null mutants showed very little detectable acetylation of pABA, arguing against a contribution from Nat3. Activity levels in null mutants ranged from 0.04 to 0.5 nmol/min/mg protein. These figures lie within the variation in background activity seen in liver extracts incubated in the absence of acetyl CoA. The Western blot analysis confirms the lack of NAT2 in the spleen of Nat2−/- animals (Figure 1B).
To investigate whether the activity difference in Nat2+/+ and Nat2+/- mice arises from a difference in NAT2 protein levels, we tested the samples for NAT2 protein by Western blotting (Figure 1B), using a polyclonal antiserum raised against the C-terminal do-decapeptide from human NAT1 (Stanley et al. 1996) and previously shown to be specific for mouse NAT2 and human NAT1(Stanley et al. 1997). Densitometric analysis of Western blots suitably exposed for quantitative analysis shows that the amount of NAT2 protein in the heart of Nat2+/+ adults is 29 ± 6% (n=6) that in adult liver (Figure 1B). Similar densitometric analyses indicate that the amount of NAT2 protein in hearts of wild-type animals (n=6) is 1.9 ± 0.3 times greater than in heterozygotes (n=6), demonstrating that there is no compensation in expression of the remaining wild-type allele in Nat2+/- animals. Prolonged exposure of these blots showed no immunore-activity in the position of NAT2 in Nat2−/- samples.
Nat2 Expression Is Specifically Localized within the Neonatal Heart
Nat2−/- mice were generated by directed insertion of lacZ into the coding region of the murine Nat2 gene (Cornish et al. 2003). Using β-galactosidase expression driven by the Nat2 promoter, we have mapped Nat2 expression in neonatal hearts of 15 pups taken from three independent intercross matings (Figures 2A-2C). Hearts from heterozygotes and homozygous null mutants clearly show intense staining in the area of fibrous continuity between the mitral and aortic valve leaflets (Figures 2B and 2C); staining is also evident in the pulmonary artery. By contrast, staining is barely detectable in the ventricles. Analysis of sectioned X-gal-stained hearts suggests that the faint staining visible in the ventricles of wholemounts may be attributable to staining of coronary vessels. Heterozygotes show the same distribution of X-gal staining as homozygotes, but in heterozygotes the staining is less intense than in Nat2−/- animals that bear two copies of the NAT2/lacZ fusion gene.
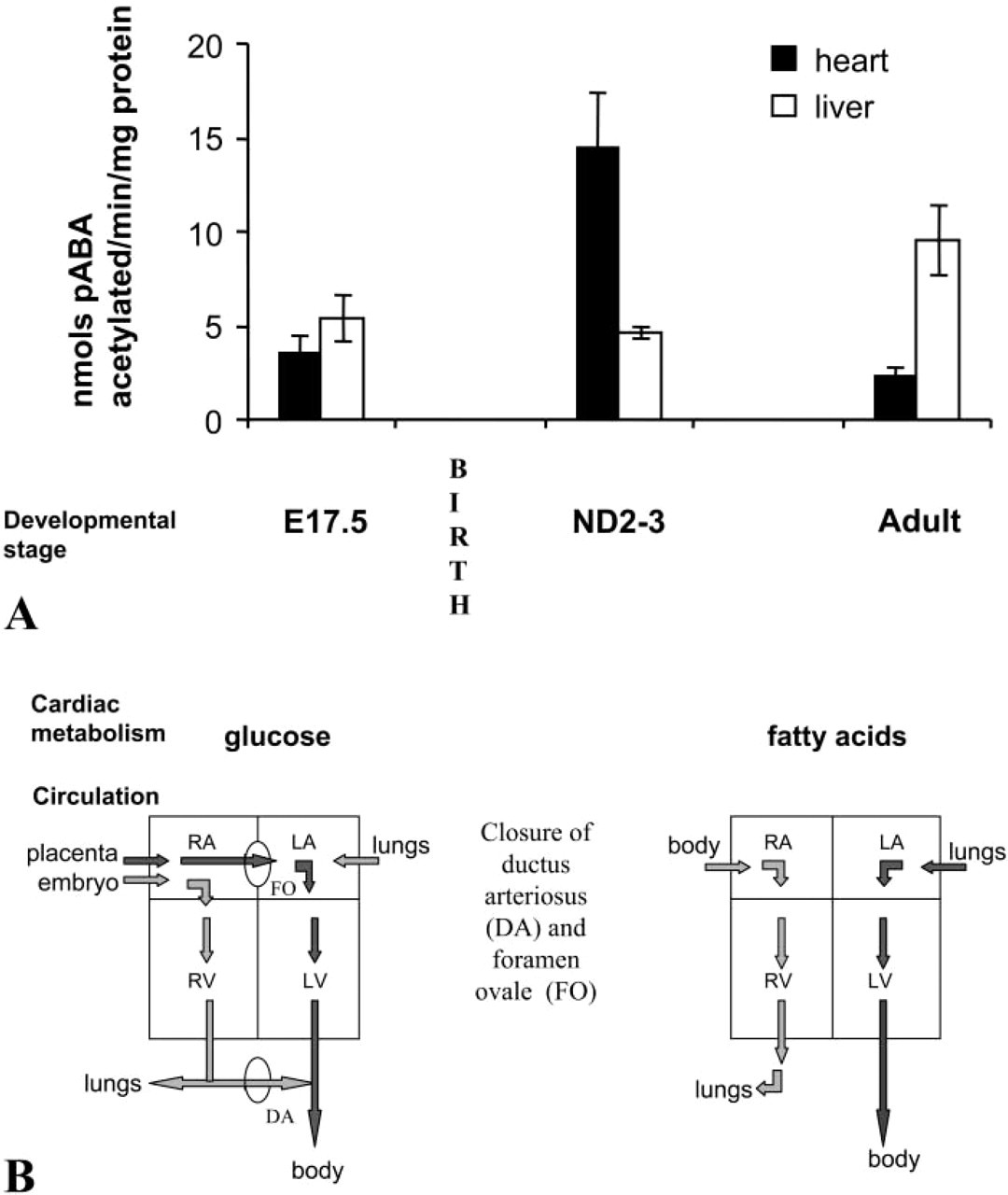
Nat2 Activity in Heart Is Developmentally Controlled and Is Highest around Parturition
To test whether there is a postpartum developmental change in cardiac NAT2 activity, we examined acetylating activity in vitro of prenatal (E17.5) and neonatal day (ND) 2–3 hearts, comparing the NAT2 acetylating activity of heart homogenates before, during, and after the postpartum metabolic and circulation changes (Figure 3A). Maximum NAT2 activity is seen in the protein homogenates prepared from the hearts of neonatal pups (n = 11), at which time it is at least 5-fold greater than that of adult heart homogenates (Figure 3A). NAT2 activity of fetal hearts (n=6) shortly before parturition (E17.5) is as low as in the adult heart. These data are consistent with those obtained from X-gal staining in that staining is maximal within 3 days of birth. To establish whether this developmental change in NAT2 activity is common to other tissues, we measured the activity of livers taken from the same animals. In contrast to the heart, the specific activity of NAT2 in the liver increases slightly between E17.5 and ND3 and is highest in the adult.
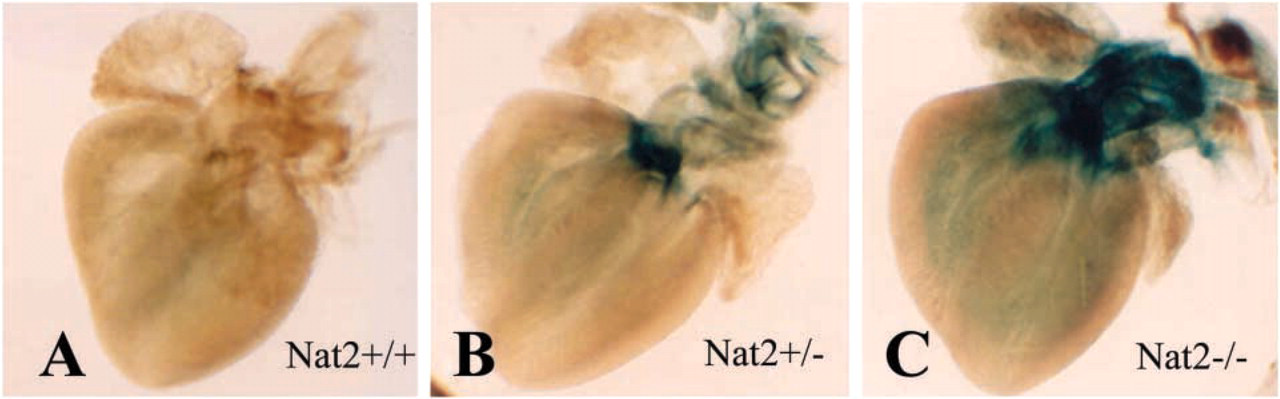
Location of Nat2 expression in whole-mount neonatal mouse hearts. β-galactosidase staining in hearts from Nat2+/+, Nat2+/-, and Nat2−/- neonatal day 3 (ND3) mice. Neonatal pups arising from three independent intercross C57Bl/6 matings were dissected and prepared as whole-mount preparations.
(
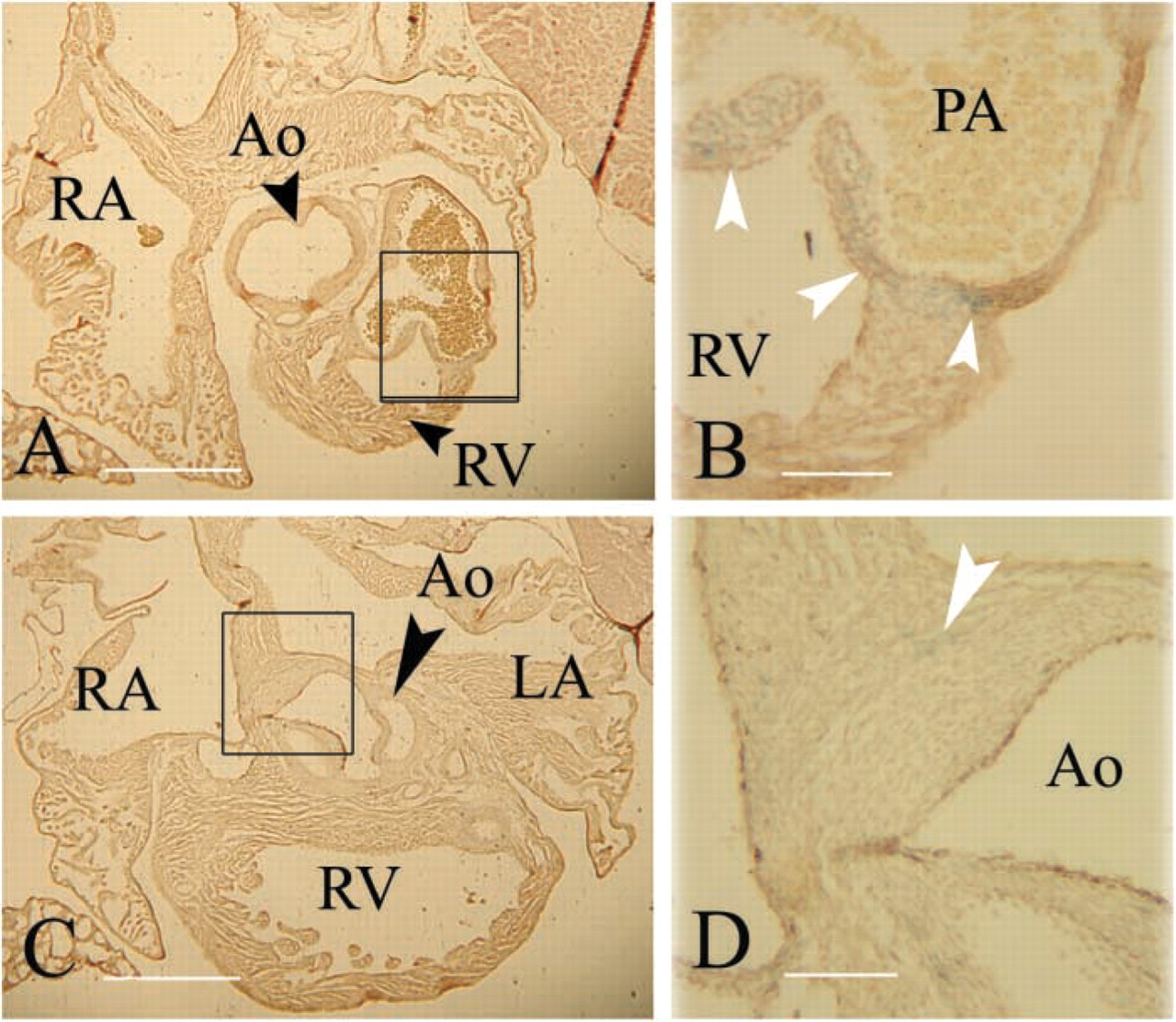
In Neonates, Nat2 Expression Is Widespread in the Central Fibrous Body and Heart Valves and Is Localized in Clusters of Cells within the Atria and Pulmonary Artery
Localization of Nat2 expression was examined in more detail in serial sections of X-gal-stained hearts dissected from ND2 pups. Hearts isolated from Nat2-/- neonates show no obvious morphological abnormalities. X-gal staining, indicative of NAT2 expression, is most prominent in the cells of the fibrous continuity (Figure 4A). A field of cells from the mitral to the tricuspid valve shows uniform staining (Figures 4A and 4C). Scattered cells in the epicardial tissue of the atria and a cluster of cells in the region of the sinoatrial node show robust staining (Figure 4B). Staining is visible in the leaflets of the atrioventricular valves (Figures 4A and 4C) and in the valves and walls of the great vessels (Figures 4D and 4E). The walls of the pulmonary artery have interspersed but strongly staining cells (Figure 4E) within the smooth muscle layer. Neither the working myocardium nor the His bundle shows detectable staining. X-gal staining of Nat2+/+ hearts revealed no evidence of endogenous β-galactosidase expression. By ND5, X-gal staining in the trigone has diminished, as has staining in the atrioventricular valves. Nat2 gene activity, as evidenced by X-gal staining, remains in the pulmonary valve and in the walls of the pulmonary artery (Figures 5A and 5B). A few cells at the level of the aortic valve leaflets show β-galactosidase activity (Figures 5C and 5D).
Nat2 Expression in Hearts of E11.5 Fetuses
Expression of β-galactosidase driven by the Nat2 promoter from the Nat2 knockout allele in stage E11.5 mouse fetuses has been investigated in whole-mount heart preparations, shown in Figure 6. Strong X-gal staining is visible in the right ventricle and in the wall of the primary heart tube. Expression of mNat2 in the right ventricle has previously been identified immuno-histochemically in E11.5 fetal mice and can be seen in the right atrium at E13.5 (Stanley et al. 1998), reinforcing the assertion that X-gal staining in mice carrying the Nat2 null allele can be used to map Nat2 expression. Although NAT expression in small groups of cells within the walls of the primary heart tube has not previously been observed in mice by immunohistochemical methods, these scattered NAT-expressing cells would not be expected to be present in all sections. In the absence of lineage analysis data, we are unable to say at the present time whether the cells staining positive for NAT in different regions of the developing heart share a common ontogenetic origin.
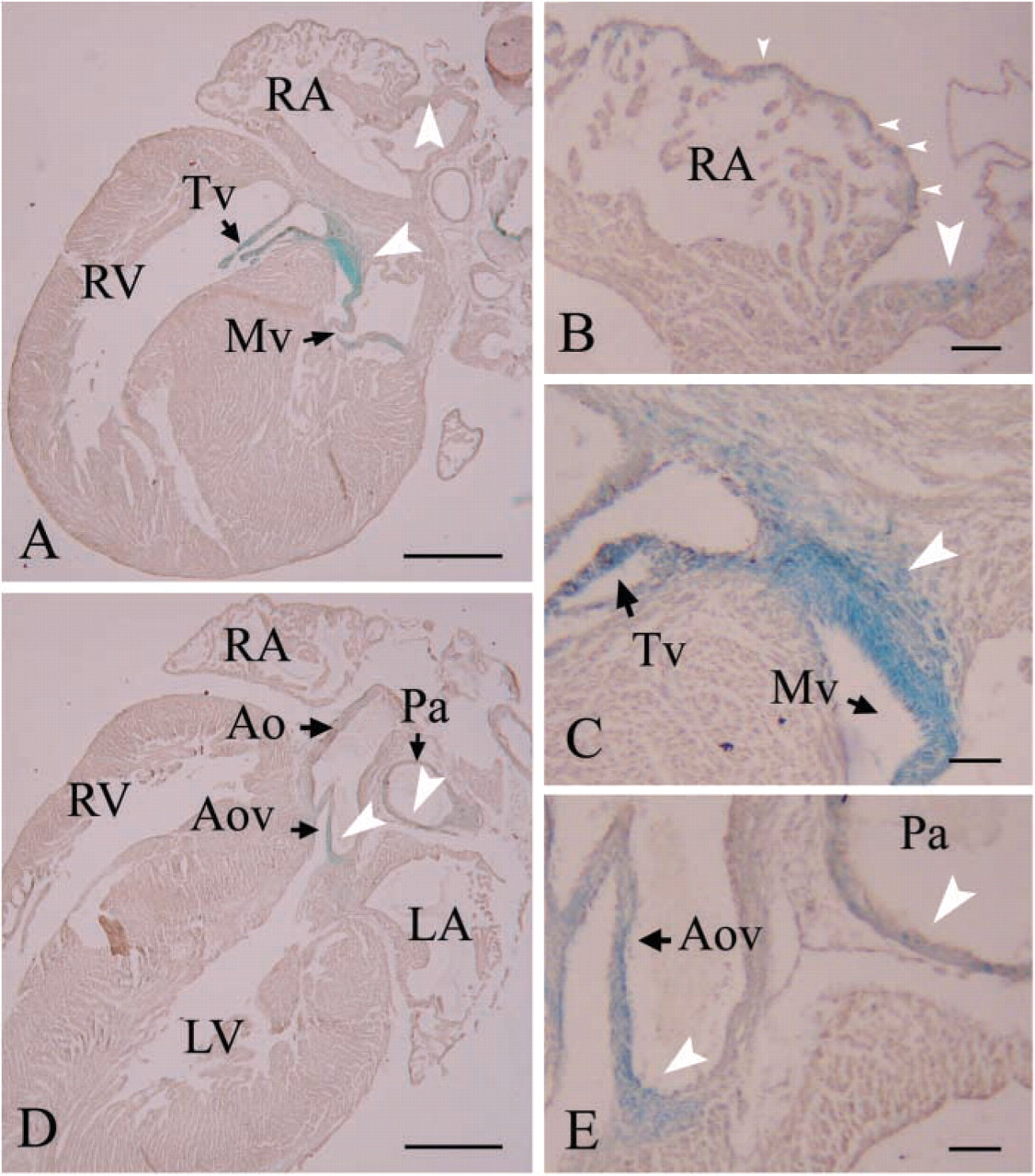
Nat2 expression profile in ND2 hearts from C57Bl/6 Nat2−/- mice. X-gal-stained neonatal hearts isolated from ND3 C57Bl/6 pups generated by intercross mating were dehydrated, embedded in paraffin wax, sectioned at 10 μm, and counterstained with Meyer's hemalum. Expression of Nat2 is indicated by blue X-gal staining (white arrowheads); black arrows in
Discussion
The expression of mouse NAT2 is both spatially and temporally controlled over different stages of cardiac development, encompassing fetus, neonate, and adult. The expression of the Nat2 gene has been assayed in heart tissue by monitoring the expression of the lacZ gene, driven by the Nat2 control elements. Previous studies support the lacZ gene expression as a realistic indicator of Nat2 gene expression in this strain of mice compared with laboratory-inbred strains (Stanley et al. 1998; Cornish et al. 2003). In addition, it has recently been demonstrated, using real-time PCR, that the expression of the mNAT2 gene is not affected by the introduction of the lacZ cassette (Loele et al. 2005).
Cardiac Nat2 expression has diminished by ND5. Coronal sections of an X-gal-stained heart isolated from a ND5 C57Bl/6 Nat2−/- pup.
The activity of the NAT2 enzyme has been assayed in mice carrying two, one, or zero copies of the Nat2 null allele. In heart tissue from the rapid acetylating strain C57Bl/6, the levels of NAT2 protein correlate with activity levels in Nat2−/-, Nat2+/-, and Nat2+/+ genotypes, indicating that there is no compensation either from the remaining wild-type allele, in the case of the heterozygotes, or from the other Nat genes. Similar results have been obtained observing expression of each individual hNAT1 allele in heterozygous and homozygous individuals (Butcher et al. 2004).
The work of Boukouvala et al. (2003) shows that Nat2 gene expression is differentially regulated in adult mouse heart compared with other tissues. These authors demonstrated the inclusion of a non-coding exon in Nat2 RNA isolated from most tissues, mapping a transcription start site 5’ to the non-coding exon, and detecting use of alternative splice sites in a variety of non-cardiac adult mouse tissues. In contrast, transcripts isolated from adult heart tissue lack the non-coding exon; thus, in heart, transcription is likely to be driven by an alternative promoter, proximal to the initiating codon (Boukouvala et al. 2003). The use of a proximal promoter has also been described in the kidney, together with modulation of transcription through androgen and glucocorticoid receptors, acting on an upstream hormone-response element (Estrada-Rodgers et al. 1998). Transcriptional regulation of murine Nat2 gene expression in the kidney is gender specific (Estrada et al. 2000), as is the case with the phase 1 drug-metabolizing protein CYP2J5 (Ma et al. 2004). A variety of genes expressed in cardiac tissue give rise to gender-related differences in cardiac function when ablated; these include proteins with structural (Du et al. 2003), metabolic (Nohammer et al. 2003), and signaling roles (Takahashi et al. 2003; Li et al. 2004), and insulin-resistant diabetic mice show gender-selective attenuation of cardiac potassium currents (Shimoni et al. 2004). In all these instances, the phenotype is more pronounced in males. Although a significant difference in the acetylating activity of NAT2 from male and female heart homogenates is not detected, preliminary proteomic analysis of Nat2+/+ and Nat2−/- animals suggests differential changes in the profile of cardiac proteins in male and female mice. These studies will form the basis for subsequent investigation of the effects of ablating Nat2 expression in the heart.
Changes in the Nat2 gene activity in murine fetal tissue and postpartum liver have been examined in terms of both RNA levels and acetylation activity (McQueen et al. 2003; McQueen and Chau 2003). These authors find that hepatic NAT2 activity increases gradually from parturition to adult. The results described here are in good agreement, showing hepatic NAT2 acetylation levels of 4 nmol/min/mg protein at ND2 and 3. In contrast to the acetylating activity of the liver, in the heart, NAT2 activity is highest in neonates, decreasing 7-fold between neonate and adult. We have considered the possibility that low levels of acetylation in adult hearts arise from the increased proportion of myocardial cells and fibrous tissue, reducing the concentration of NAT protein in the preparation, but with NAT activity remaining at the neonatal level on an individual cell basis. Examination of 5-day-old neonates suggests this is not the case, inasmuch as very little X-gal staining remains in the fibrous trigone or the cardiac valves. At this stage, only the pulmonary valve shows extensive staining. Thus, the lower activity in adult hearts is likely to arise from the loss or reduction of NAT2 activity from a subset of cardiac cells.
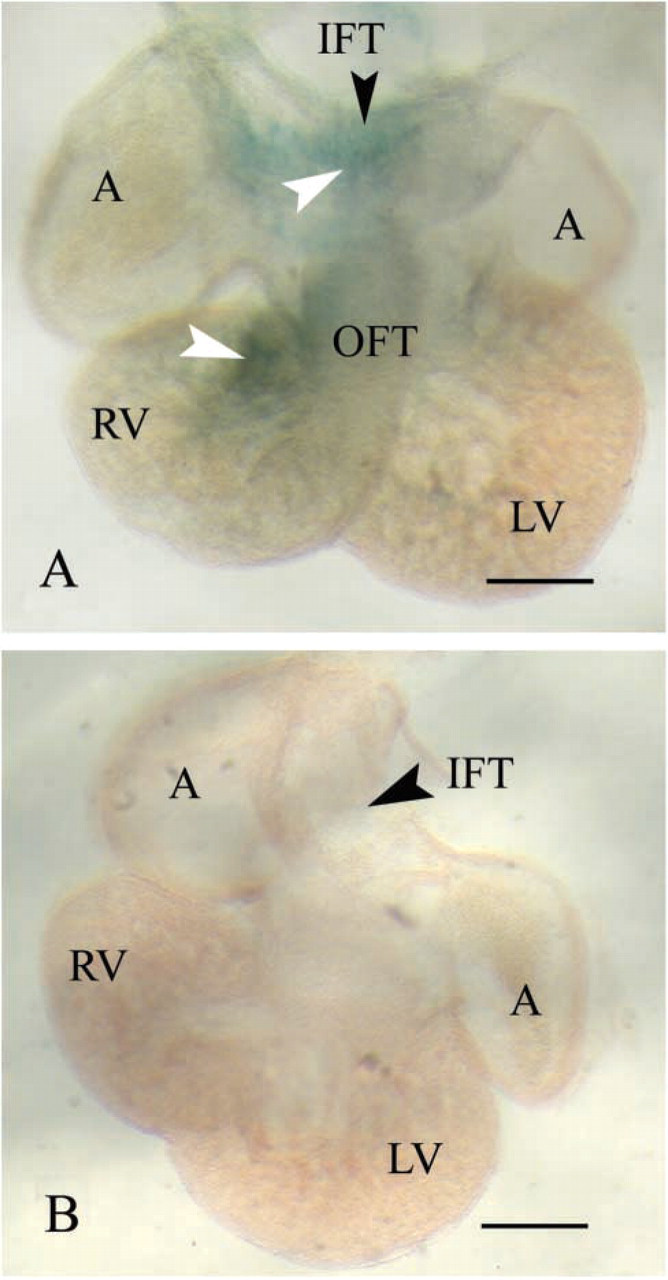
Cardiac NAT2 expression in E11.5 fetuses, visualized by X-gal staining. X-gal staining is indicated by white arrowheads; black arrowheads indicate the position of named structures. Embryos taken from three independent intercross matings were stained with X-gal, dehydrated, and cleared, as before (Cornish et al. 2003), and the hearts dissected and photographed as whole-mount preparations. Nat2−/- fetuses expressing lacZ from the Nat2 null allele (
Major changes in the structure and metabolism of the heart occur shortly after parturition. During fetal development, when the lungs are functionally inactive, the ventricles of the heart operate in parallel and some protection from hypoxia is afforded by use of glucose as opposed to fatty acid as source of myocardial energy. On parturition, the pattern of circulation changes, with the closure of the foramen ovale and ductus arteriosus, largely in response to pressure changes arising from occlusion of the umbilical vessels and decreased pulmonary vascular resistance (shown schematically in Figure 3B). Postnatally, the use of fatty acids predominates over that of glucose.
Oxidative stress is an important determinant of homocysteine levels, production of toxic cellular byproducts, and xenobiotic metabolism. Increased generation of free radicals accompanies reoxygenation of isolated hearts after hypoxia (Damerau et al. 1993). Cellular redox status has been shown to modulate human NAT1 function (Atmane et al. 2003). The observation that murine NAT2 is largely absent from the working myocardium argues against a generalized role for NAT2 in energy metabolism within myocardial cells, although it does not exclude the possibility of a role modulating hypoxic effects in other cell types within the heart.
Murine Nat2 expression is most marked in the fibrous continuity of the neonatal heart. It is, however, also expressed along the length of the valves that regulate the flow of blood into both the ventricles and the major vessels, and in the wall of the right atrium in close proximity to the sinoatrial node. Highest levels of autonomic innervation are to be found in the region of the sinus node (Chow et al. 2001). Neuronal expression of murine Nat2 and its homolog, human NAT1, has been found both in mice and in humans (Stanley et al. 1998). In this context, the presence of Nat2-expressing cells in the wall of the right atrium in the region of the sinus node and in the region of the atrioventricular node, from the looped heart stage, is of particular interest.
Although Nat2−/- mice are overtly aphenotypic, our results showing highly specific temporal and spatial control during cardiac development are indicative of an endogenous role distinct from that of xenobiotic metabolism. Preliminary proteomic studies suggest that the Nat2−/- mutation is complemented by changes in other cytoplasmic proteins. The expression of polymorphic murine Nat2/human NAT1 in locations associated with pacemaker activity and autonomic innervation suggest that mNat2 activity should be investigated in relation to cardiac function.
Footnotes
Acknowledgements
The authors thank the Wellcome Trust for financial support.
We thank Hilary Long for outstanding technical assistance, Matt Anderton for reading an early draft of the manuscript, Robert Anderson and Shuomo Bhattacharya for help with cardiac morphology, and Jennie Charlton for secretarial support.