
Abstract
The MDR1 gene product, P-glycoprotein (P-gp), was shown to confer multi-drug resistance to cancer cells, but its overexpression is also suggested to be involved in pharmacoresistance of epilepsy by acting as an energy-dependent drug-efflux pump in the blood-brain barrier (BBB). In normal brain tissue, P-gp is almost exclusively expressed by capillary endothelial cells (EC) of the BBB, whereas little or no expression is detected in other cell types. Increased P-gp expression was observed after seizures, but localization of this increase, i.e., within brain capillary EC or within parenchymal or perivascular astrocytes, which contribute to the BBB function, is controversial. To test whether these antithetic data arise from unusual properties of the antigen itself, we compared different immunohistochemical techniques and monoclonal or polyclonal antibodies to P-gp in normal rat brain and rat brain after kainate-induced seizures. Using acetone-fixed cryostat sections of snap-frozen tissue, strong P-gp labeling was detected in EC and, after seizures, in hippocampal neurons, but not in astrocytes. In contrast, EC and neuronal P-gp immunolabeling were not seen in paraformaldehyde-fixed sections, whereas both perivascular and parenchymal astrocytes exhibited strong P-gp labeling after seizures. The lack of P-gp labeling in EC by paraformaldehyde fixation, was reversed by treatment of the sections with acetate/ethanol. These experiments demonstrate that various fixation conditions have a striking effect on the immunohistochemical localization of P-gp in rat brain and detection of its increased expression by seizures. When data obtained from different immunohistochemical techniques are taken together, seizures seem to induce overexpression of P-gp in four different cell types, i.e., EC, perivascular astrocytes, parenchymal astrocytes, and neurons.
T
In the brain, P-gp is thought to be primarily located in the apical (luminal) membrane of capillary endothelial cells that form the blood-brain barrier (BBB) and to be one of the mechanisms involved in protecting the brain from xenobiotics (Schinkel 1999; Bendayan et al. 2002; de Boer et al. 2003). Various Western immunoblotting and immunohistochemical studies, using confocal and electron microscopy in different species including humans, have demonstrated the luminal expression of P-gp in brain capillary endothelial cells with little or no expression in other brain cells (e.g., astroglia or neurons) under normal conditions (Schinkel 1999; Lee et al. 2001; Abbott et al. 2002). However, based on immunocytochemical studies on human and monkey brain capillaries, Pardridge and colleagues questioned this view but proposed that P-gp is primarily localized at the astrocyte foot processes that invest the brain microvascular endothelium (Pardridge et al. 1997; Golden and Pardridge 2000). This proposal was criticized (e.g., Schinkel 1999; Abbott et al. 2002) because it was essentially based on findings with only one P-gp antibody (MRK16). It is known that the extracellular epitope of human P-gp for the monoclonal antibody MRK16 can be completely shielded in some cell types by heavy N-glycosylation (Schinkel 1999). P-gp immunohistochemistry is further highly sensitive to preparation and fixation of samples. Suboptimal fixation may lead to lack of recognition by obliteration of epitopes or increased nonspecific binding of the primary antibody (O'Brien and Cordon-Cardo 1996; Schinkel 1999). Based on these obstacles, immunolocalization of P-gp can only be considered reliable when consistent results are obtained with at least two, and preferably three or more, different antibodies (Schinkel 1999; Scheffer and Scheper 2002). In a recent study using a rabbit polyclonal anti-serum [mdr-(Ab-1)] against P-gp and different tissue fixation protocols, Schlachetzki and Pardridge (2003) demonstrated dual expression of P-gp at astrocytes and endothelium of normal primate brain, thus obviously resolving the previous conflicting results. However, most other groups did not detect P-gp in astrocytes of normal brain tissue, indicating that fixation and staining variables might be critical. Clear expression of P-gp in astrocytes is seen in certain pathological states such as epilepsy (Marroni et al. 2003; Sisodiya 2003). Furthermore, brain tissue from patients with intractable epilepsy exhibits an increased expression of P-gp in endothelial cells, which is thought to reduce the ability of antiepileptic drugs to reach their neuronal targets (Löscher and Potschka 2002; Marroni et al. 2003; Sisodiya 2003). In addition, P-gp has recently been detected in neurons of epileptogenic human brain tissue (Aronica et al. 2003; Marchi et al. 2004).
Differences among laboratories have also been reported for P-gp immunolocalization in the rat brain. In primary cultures of rat brain astrocytes, P-gp expression has been described, but at lower levels when compared with primary endothelial cultures (Decleves et al. 2000). The latter authors also showed that both mdr1a and mdr1b are expressed in endothelial cells, whereas astrocytes preferentially expressed mdr1b (Decleves et al. 2000). In the normal rat brain, most researchers did not detect P-gp in astrocytes, but Zhang et al. (1999) reported P-gp labeling of cell bodies and foot processes of astrocytes in the hippocampus following kainate-induced seizures. Confocal microscopy of double-immunofluorescence-stained sections of rat hippocampus showed that P-gp was expressed in the same cells as GFAP, a marker for astrocytes (Zhang et al. 1999). However, using the same antibodies to P-gp, C219, and mdr-(Ab-1), and confocal microscopy of double-immunofluorescence-stained sections of the hippocampus of rats after kainate-induced seizures, we could not reproduce the findings of Zhang et al. (1999), but unequivocal staining was confined to capillary endothelial cells (Seegers et al. 2002a, b; Volk et al. 2004a, b). Furthermore, following kainate- and pilocarpine-induced seizures, we recently demonstrated expression of P-gp in hippocampal neurons, but again not in astrocytes (Volk et al. 2004b). The only obvious difference between the experiments of Zhang et al. (1999) and our group was that Zhang et al. (1999) used paraformaldehyde-fixed vibratome sections whereas we used acetone-fixed cryostat sections of snap-frozen tissue for P-gp immunohistochemistry. Our protocol was based on studies of Thiebaut et al. (1989) with the monoclonal P-gp antibody C219, showing that formaldehyde fixation dramatically reduced localization of P-gp at the brain capillaries when compared with acetone-fixed sections, thus indicating that the epitope reactive with C219 is very aldehyde sensitive. C219 is by far the most widely used antibody for P-gp immunodetection in both research and diagnosis but cross-reacts with other proteins in addition to P-gp, thus rendering ambiguous results (van Den Elsen et al. 1999).
Many of the physiological functions of P-gp are still unknown (Scheffer and Scheper 2002). To better understand the functional roles of P-gp in the brain, it is essential to know its cellular localization under both physiological and pathological conditions. In the present study we directly compared the different protocols used for P-gp immunohistochemistry by Zhang et al. (1999) and our group in normal rat brains and after kainate-induced seizures, using three different P-gp antibodies. Because freezing, pH, and dehydration are additional variables in immunohistochemical localization of proteins in rat brain (Werner et al. 2000; Montero 2003), we examined such variables also with respect to P-gp immunostaining.
Materials and Methods
Animals
As in our previous studies on P-gp expression in rats (Seegers et al. 2002a, b; Volk et al. 2004a, b), female Wistar outbred rats (Harlan-Winkelmann; Borchen, Germany) weighing 200-230 g were used. Animals were purchased from the breeder at ~10 weeks of age. Following arrival in the animal colony, rats were kept under controlled environmental conditions (ambient temperature 24-25C, humidity 50-60%, 12/12 hr light/dark cycle, light on at 6:00
Induction of Status Epilepticus by Kainate
Rats were injected IP with 10 mg/kg kainate (Sigma-Aldrich; Munich, Germany) dissolved in 3 ml/kg isotonic saline (pH 7.2). About 60% of rats developed a convulsive status epilepticus (SE) at this dose of kainate, consisting of ongoing generalized convulsive seizure activity. The time that a rat spent in SE with generalized motor seizures was limited to 90 min by IP administration of diazepam (10 mg/kg IP). Kainate-treated rats (n = 15) were killed 24 hr after the SE. Together with kainate-treated rats, a group of control rats (n = 13) was prepared. These controls were treated with saline instead of kainate and received diazepam, as did the kainate-treated rats. Controls were killed together with kainate-treated rats and were concurrently used for P-gp expression analysis.
Tissue Preparation
To determine the effect of fixation variables on P-gp expression, various protocols were used for tissue preparation. In one protocol (protocol #1: acetone-fixed cryostat sections of snap-frozen tissue), rats were decapitated and brains were immediately embedded in Tissue Freezing Medium (Jung; Nussloch, Germany), rapidly removed and snap frozen in liquid nitrogen. Frozen brains were stored at −80C for at least overnight. Frozen transverse sections of forebrain and midbrain were cut at 14 μm using a cryostat (HM 560 M, Microm; Walldorf, Germany) and fixed in 100% acetone for 10 min at −20C.
In a second protocol (protocol #2: paraformaldehydefixed cryosections), rats were deeply anesthetized with chloral hydrate (500 mg/kg IP) and transcardially perfused with phosphate-buffered saline (PBS; 0.01 M phosphate-buffered 0.9% NaCl solution, pH 7.6) for ~10 sec, followed by perfusion with ~300 ml of a fixative (4C) containing 4% paraformaldehyde, 0.05% glutardialdehyde, and 1.2% picric acid in 0.1 M phosphate buffer (pH 7.6). After fixation, brains were removed and cryoprotected in 30% sucrose in 0.1 M phosphate buffer (pH 7.6) at 4C for 3 days, frozen, and cryosectioned at 40-μm thickness on a freezing microtome (Frigomobil 1205; Jung, Heidelberg, Germany).
In a third protocol (protocol #3: paraformaldehyde-fixed vibratome sections), which was almost identical to the protocol used by Zhang et al. (1999), rats were deeply anesthetized and perfused with paraformaldehyde/glutaraldehyde as described in protocol #2, but brains were immediately sectioned at 100-μm thickness using a vibratome (Series 1000; Technical Products International Inc., St Louis, MO).
Sections obtained following the three protocols were mounted on slides and used for immunohistochemical labeling of P-gp. For potential unmasking of P-gp epitopes (Shi et al. 1997), part of the sections obtained following protocols #2 and #3 were pretreated with a mixture of 33% acetic acid and 66% ethanol for 10 min at −20C. We have previously used this mixture to optimize immunostaining of terminal deoxynucleotidyl biotinylated dUTP nick-ending labeling (TUNEL) in formaldehyde-fixed sections of rat brain after kainate-induced seizures (Brandt et al. 2003). Thus, we investigated whether acetate/ethanol also optimizes P-gp staining in formaldehyde-fixed sections.
For comparative evaluation of P-gp staining obtained with different protocols, we chose sections of the hippocampus, because this brain region is particularly sensitive to kainate and thought to be critically involved in its epileptogenic activity (Sperk, 1993). Therefore, sections from −2.3 to −3.8 mm relative to bregma were chosen (Paxinos and Watson 1998).
For each protocol, three P-gp antibodies were used (see below). The different antibodies were compared in the same animal in adjacent brain sections for a given fixation protocol, and this experiment was repeated in at least four rats per protocol.
Immunohistochemistry
In a first step, sections were extensively washed in 0.05 M Tris-buffered saline (TBS; pH 7.6) and incubated in 0.5% H2O2 to block endogenous peroxidase activity. Because this step might in itself damage antigens, we also evaluated a milder method to block endogenous peroxidase by using a 0.05% solution of phenylhydrazine. This, however, led to increased nonspecific background staining without changing the cellular localization of P-gp in a given fixation protocol, so that we preferred to use the H2O2 incubation described above for all experiments mentioned in the present study. In this respect, it is important to note that, although brain tissue on average contains only low levels of peroxidase as compared with other tissues, brain capillary endothelial cells contain endogenous peroxidase at levels that might crisscross the accurate interpretation of P-gp expression, particularly pertaining to the aim of this study.
To inhibit unspecific antibody binding, sections were first incubated with a blocking solution containing 2% bovine serum albumin, 0.3% Triton X-100, and 5% normal pig or rabbit serum in TBS for 60 min directly followed by incubation with the primary antiserum for 12 hr at 4C. Next, sections were rinsed with TBS and incubated with biotin-labeled secondary antiserum (pig anti-rabbit or rabbit anti-mouse IgG, 1.2 (μg/ml; DAKO, Hamburg, Germany) for 60 min. Sections were subsequently rinsed with TBS and incubated with horseradish peroxidase-labeled streptavidin (1.65 μg/ml; DAKO) for 90 min, followed by a nickel-intensified diaminobenzidine (DAB) reaction (0.05% 3,3-diaminobenzidine and 0.6% ammonium nickel sulfate; both from Sigma) in the presence of 0.01% H2O2 for 15 min. Antibodies were dissolved in TBS containing 1% pig or rabbit serum, 1% bovine serum albumin, and 0.3% Triton X-100. Sections were washed, air dried, treated with toluol for 1 min, and coverslipped with Entellan (Merck; Darmstadt, Germany).
Antibodies
To detect P-gp expression, we used three different antibodies. The polyclonal rabbit antibody mdr-(Ab-1) (Oncogene Research Products; San Diego, CA), which was raised against the peptide SALDTESEKVVQEALDKAREG of the P-gp C-terminal cytoplasmic domain, recognizes both human and rodent P-gp, due to shared peptide sequence in this portion of P-gp (Tatsuta et al. 1992; Biegel et al. 1995). The monoclonal mouse antibody C219 (Calbiochem; Bad Soden, Germany) binds to a conserved cytoplasmic region, which is present in all classes of P-gp cytosolic nucleotide-binding domains from rodents and humans (Georges et al. 1990; Jette et al. 1993; van Den Elsen et al. 1999). It recognizes the amino acid sequence VQEALD, i.e., part of the sequence of the P-gp C-terminal cytoplasmic domain that is also recognized by mdr-(Ab-1). The polyclonal rabbit antibody H241 (Santa Cruz Biotechnology, Inc.; Santa Cruz, CA), which was raised against a recombinant protein corresponding to amino acids 1040-1280 mapping at the P-gp C-terminal cytoplasmic domain, recognizes P-gp of human, mouse, and rat origin (Califano et al. 2000; Weisberg and Griffin 2000). All three antibodies recognize both P-gp isoforms, mdr1a and mdr1b, in rodents. Their specificity for P-gp has been demonstrated by the manufacturers using Western blotting, immunoprecipitation, and immunohistochemstry.
Antibodies were used at final concentrations of 2 μg/ml (mdr-(Ab-1)), 1 μg/ml (C219), or 1 μg/ml (H241), except for immunohistochemistry on vibratome sections, for which higher concentrations were used (up to 3 μg/ml) to replicate the protocol of Zhang et al. (1999). As negative control, the same amount of pig or rabbit serum replaced the primary antibody. Preabsorption experiments were performed using each antibody and a blocking mdr-peptide (product of the human mdr-1 gene; 1.5 μg or 0.5 μg in 100 μl; Oncogene Research Products), which led to complete loss of P-gp immunostaining. This, however, does not rule out that there could be other antigens in the tissue that react with the antibodies. We therefore performed an NCBI-BLAST search using the peptide sequence SALDTESEKVVQEALDKAREG that did not show any other putative proteins that might cross-react with the antibodies used in this study. Furthermore, monoclonal antibodies such as C219 do not contain any other antibodies per se, and both monoclonal as well as polyclonal antibodies are supplied as affinity-purified products.
Scoring of P-gp-staining Intensity in Astrocytes
Because of the high background reaction caused by paraformaldehyde fixation (see Results), P-gp staining intensity could not be accurately quantified by computer-assisted image analysis based on optical density measurements that we recently used to examine immunohistochemical expression of P-gp in acetone-fixed sections (Volk et al. 2004a). Therefore, we used a semiquantitative grading system, in which intensity of P-gp expression of astrocytes within hippocampus tissue sections was scored in coded sections by an experienced investigator. The investigator took into account the background staining but was unaware whether sections were from kainate-treated rats or controls. Intensity of the P-gp reaction was graded on a scale from 0-2, with 0 = no reaction, 1 = weak reaction, 2 = strong reaction. The mean values per region determined for each rat were used for calculating group means. Significance of differences between groups (n = 6-7 per group) was calculated by the Mann-Whitney U test.
Because a similar scoring system was recently used to demonstrate significantly increased P-gp staining of endothelial cells following kainate-induced seizures (Seegers et al. 2002a, b), analysis of endothelial staining was not performed again in the present experiments. Furthermore, we recently used computer-assisted image analysis of endothelial P-gp staining based on optical density measurements, which demonstrated a significant increase of endothelial P-gp expression after kainate treatment (Volk et al. 2004a). Detection, however, was possible in acetone-fixed sections only and was not repeated in the present experiments.
Double Labeling of Immunofluorescence-stained Sections
For double labeling of cells, transverse sections were prepared according to protocol #1 (acetone-fixed cryostat sections of snap-frozen tissue) and incubated with an antibody mixture containing an antibody against P-gp (C219, 1 μg/ml) and either GLUT-1 (2 μg/ml, rabbit polyclonal antibody; Chemicon, Hofheim, Germany), or glial fibrillary acidic protein (GFAP, 0.55 μg/ml, rabbit polyclonal antibody; DAKO), respectively. For double labeling of neurons, the polyclonal P-gp antibody mdr-(Ab-1) (2 μg/ml) and a mouse monoclonal antibody against the neuronal nuclear antigen (NeuN, 2 μg/ml; Chemicon) were used. The secondary antibodies [1.2 μg/ml biotinylated goat anti-mouse IgG; DAKO, and 1.5 μg/ml carbocyanin 2-labeled goat anti-rabbit (Jackson ImmunoResearch Laboratories; Soham, Cambridgeshire, UK)] were applied for 1 hr. Sections were subsequently washed, incubated with carbocyanin 3-labeled streptavidin (1.8 μg/ml; Sigma) for 60 min, and coverslipped as described. Fluorescent signals from double-labeled sections were detected with a confocal microscope (Leica TCS SP2; Benstein, Germany).
Assessment of Neurodegeneration after Kainate Treatment
To determine neuronal damage following kainate-induced SE, an additional group of rats was treated with kainate as described, deeply anesthetized with chloral hydrate (24 hr after kainate), and transcardially perfused with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). Several series of 40-μm-thick transverse sections of the rats' forebrain were cut on a freezing microtome. Damage of brain regions was visualized by neuronal Nissl staining with thionine. Saline-treated rats served as control. Details of assessment of neuronal damage were published recently (Brandt et al. 2003).
Results
Neurodegeneration after Kainate
Twenty-four hr after a kainate-induced SE, neurodegeneration was seen in the pyramidal cell layers of the hippocampus, particularly in the CA1 (Figure 1D) and CA3c/CA4 layers (Figure 1F). Furthermore, there was a marked reduction of hilar neurons, whereas no obvious neuronal loss was observed in the granular cell layer of the dentate gyrus (Figures 1B and 1F). The severe, subregion-specific neuronal damage in the hippocampus has to be considered when evaluating neuronal P-gp expression after kainate-induced SE as described in the following.
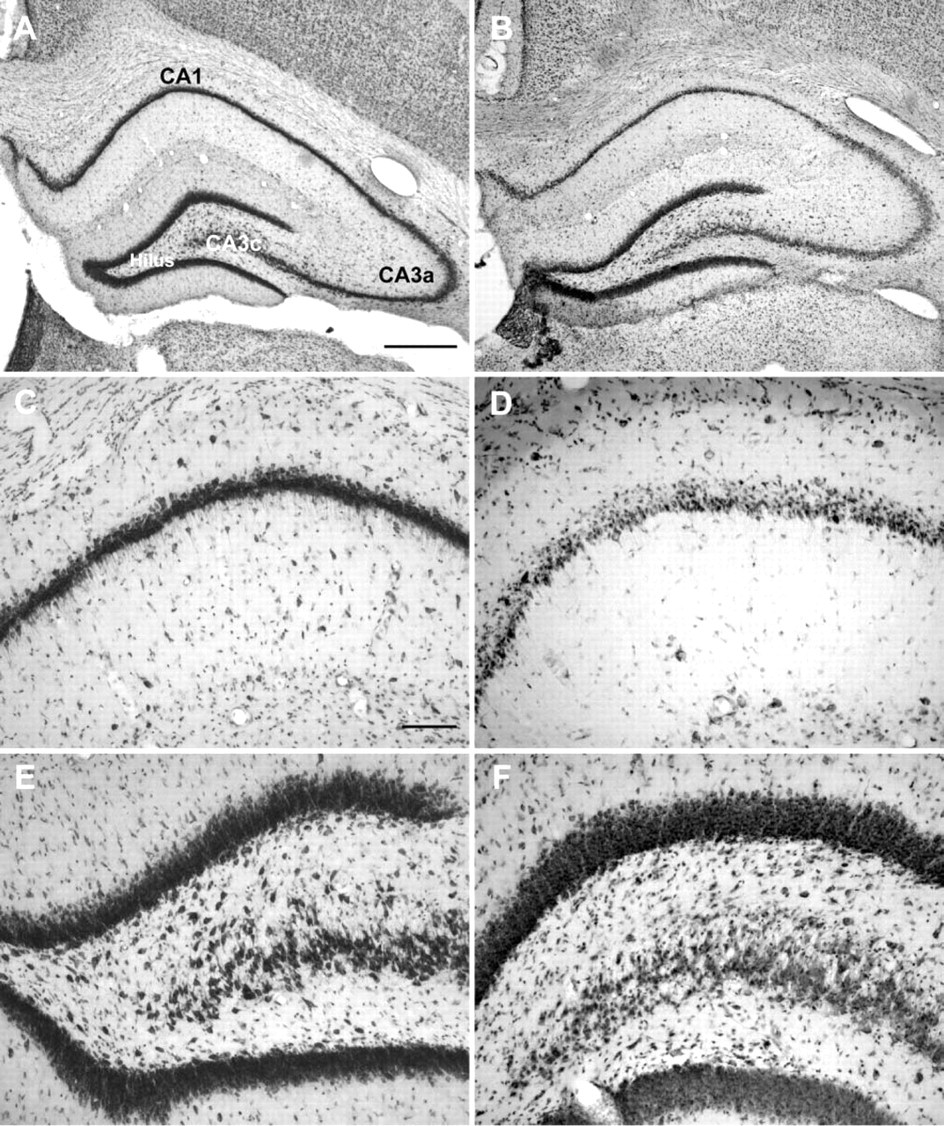
Representative thionin-stained coronal sections of the hippocampal formation of a control rat (
P-gp expression in normal rat brain determined with protocol #1 (acetone-fixed cryostat sections of snap-frozen tissue). Sections are from the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody mdr-(Ab-1) was used in
Protocol #1: Acetone-fixed Cryostat Sections of Snap-frozen Tissue
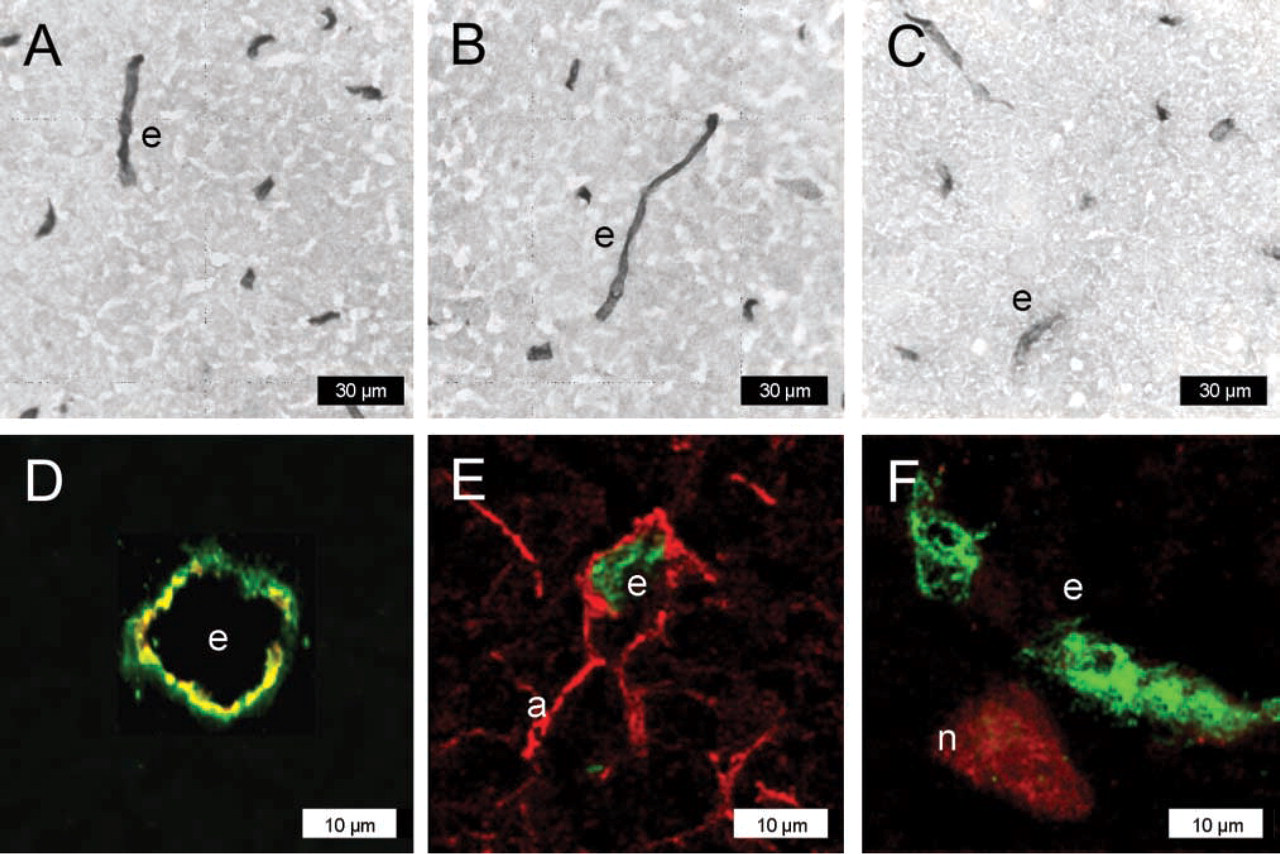
In normal rat brains, all three P-gp-antibodies detected P-gp exclusively in capillary endothelial cells (Figure 2, Figure 3A). Confocal microscopy of double-immunofluorescently stained sections using C219 or mdr-(Ab-1) for P-gp and an antibody against the brain capillary endothelial cell marker GLUT-1 confirmed the presence of P-gp in endothelial cells (Figure 2D), whereas no double labeling of P-gp was obtained with the astroglia marker GFAP (Figure 2E) or the neuronal marker NeuN (Figure 2F) in any brain region. As shown in Figure 2E, neither perivascular nor parenchymal astroglial cells expressed P-gp under the conditions of this protocol.
Following kainate-induced seizures, P-gp labeling was seen in capillary endothelial cells but not in astrocytes (Figures 3B-3D). Furthermore, neurons in the hippocampal CA3c/CA4 layer and the hilus of the dentate gyrus were labeled for P-gp by all three antibodies (Figures 3B-3D; Table 1). As reported recently (Volk et al. 2004b), double-immunofluorescencently stained sections using C219 or mdr-(Ab-1) for P-gp and an antibody against the neuronal marker NeuN confirmed the presence of P-gp in neurons, whereas no double labeling of P-gp was obtained with the astroglia marker GFAP (not illustrated).
P-gp expression with protocol #1 (acetone-fixed cryostat sections of snap-frozen tissue) in normal rat brain (
Immunohistochemical localization of P-gp in rat brain: comparison of different fixation and staining protocols
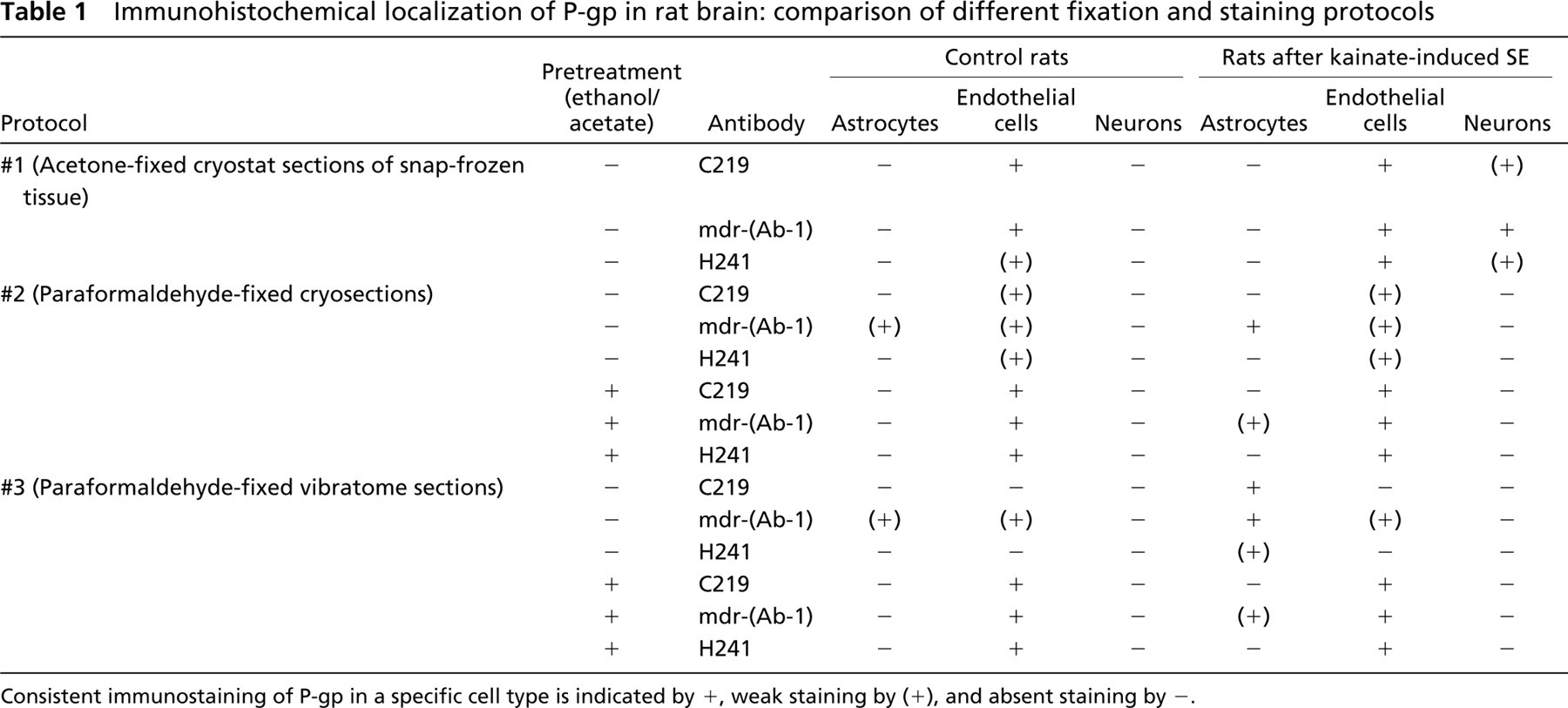
Consistent immunostaming of P-gp in a specific cell type is indicated by +, weak staining by (+), and absent staining by -.
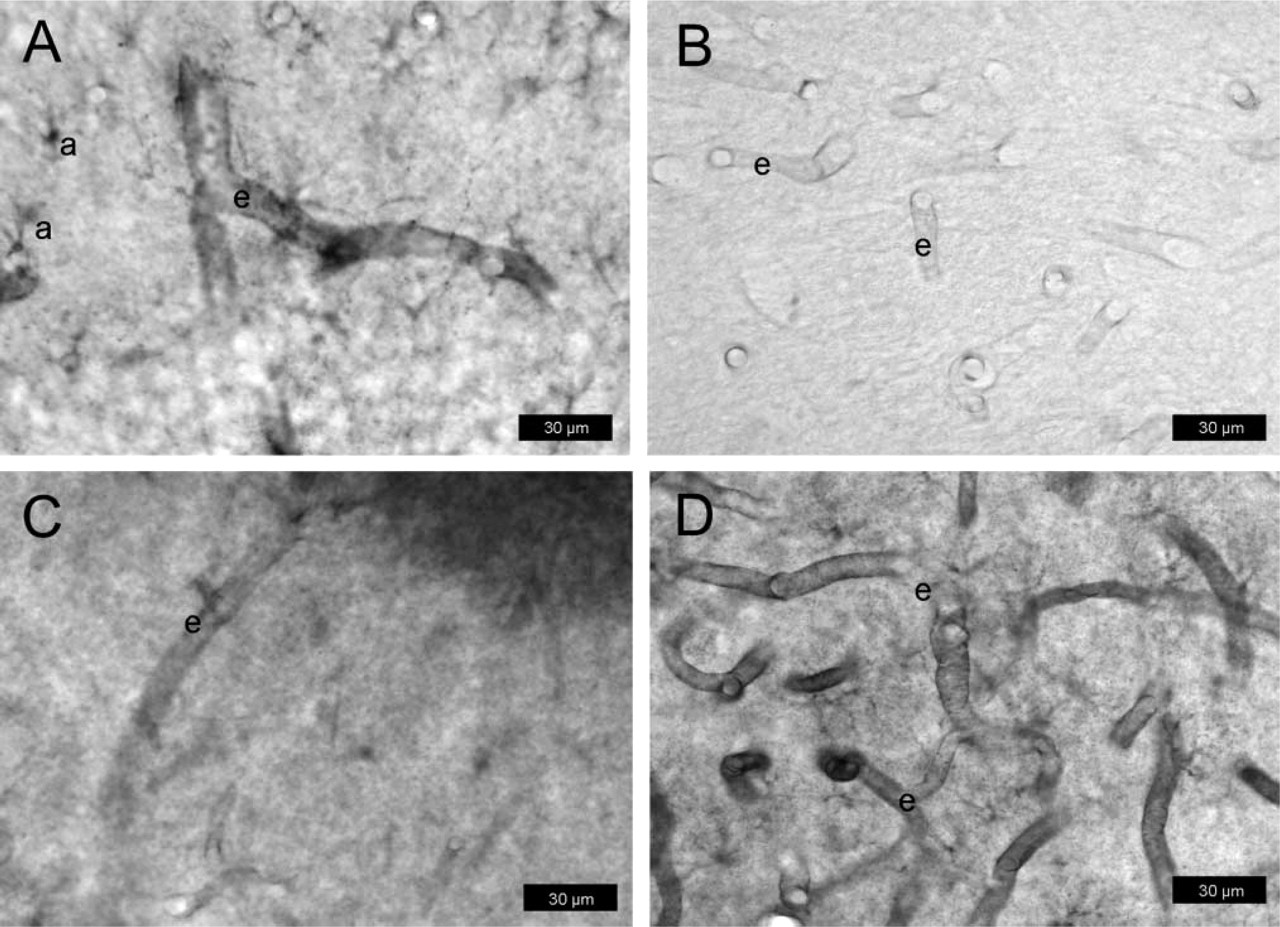
P-gp expression in normal rat brain determined with protocol #2 (paraformaldehyde-fixed cryosections). Sections are from the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody mdr-(Ab-1) was used in
Protocol #2: Paraformaldehyde-fixed Cryosections
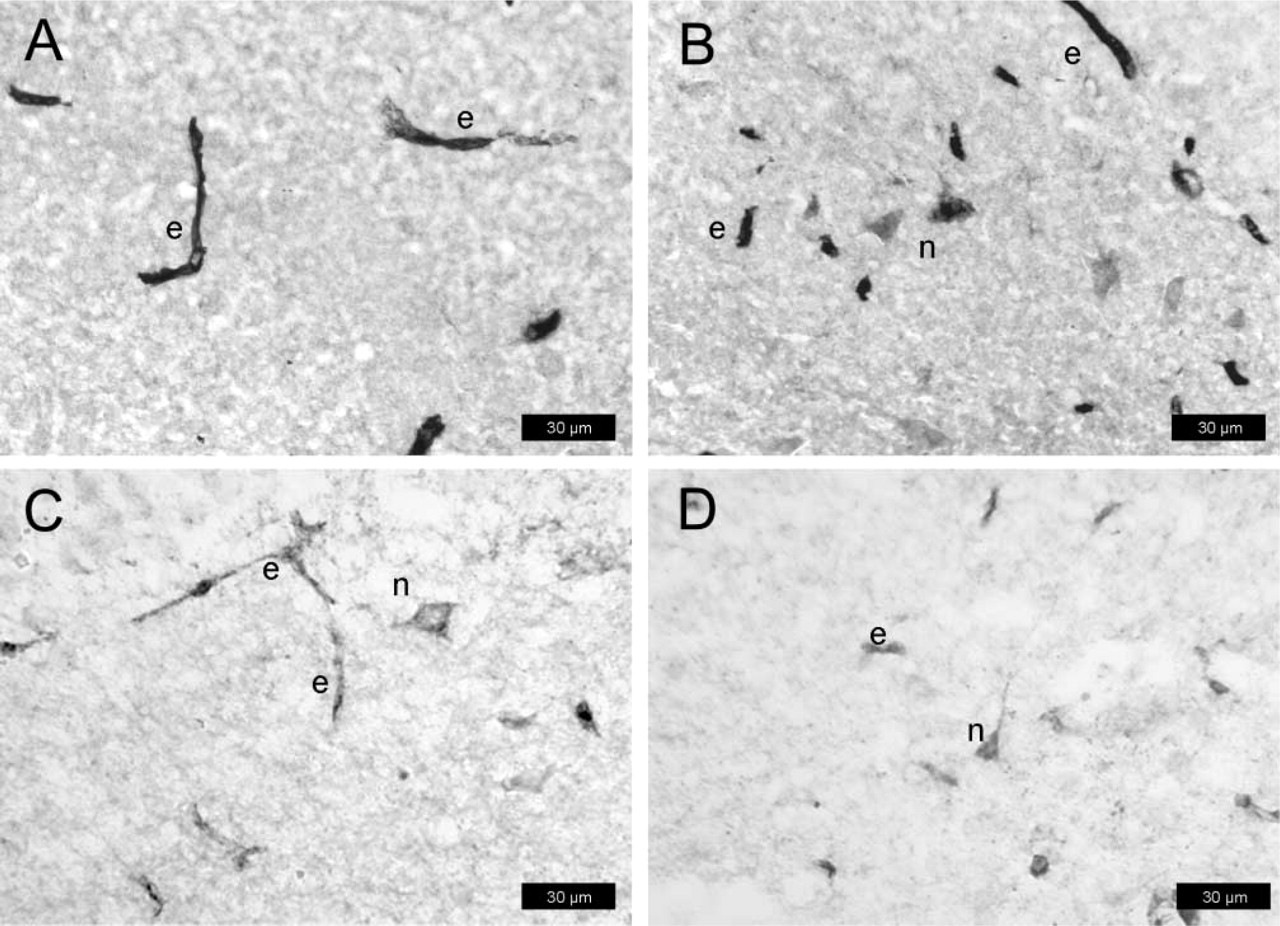
As with the sections from protocol #1, all three P-gp-antibodies detected P-gp in capillary endothelial cells of normal rat brain (Figures 4A-4C), but staining intensity was far below that obtained according to protocol #1. With all antibodies, staining intensity of P-gp in endothelial cells was markedly enhanced by treatment of sections with ethanol/acetate (Figure 4D; Table 1). With mdr-(Ab-1), weak P-gp staining was seen in astrocytes (Figure 4A) which, however, was partially abolished by treatment of sections with ethanol/acetate (Figure 4D).
Following kainate-induced SE, intense P-gp-labeling of endothelial cells was obtained with all three antibodies (Figures 5A-5C). Furthermore, enhanced astroglial labeling was seen with mdr-(Ab-1) (Figure 5A) as compared with controls (Figure 4A). The astrocytic P-gp expression was observed at both parenchymal cell bodies and processes and at perivascular astrocytes with end-feet terminating on the abluminal membrane of microvessels. Pretreatment with ethanol/acetate increased the intensity of P-gp labeling in endothelial cells (Figure 5A; Table 1). Furthermore, although no clear astroglia labeling was seen under these conditions in controls (Figure 4D), astrocytic P-gp labeling was clearly visible after seizures (Figure 5D). None of the antibodies detected P-gp in neurons under the conditions of this protocol.
Protocol #3: Paraformaldehyde-fixed Vibratome Sections
In contrast to the other protocols, only mdr-(Ab-1) weakly stained endothelial cells in normal rat brain (Figure 6A), whereas no such labeling was seen with C219 (Figure 6B) or H241 (Figure 6C). Following pretreatment of sections with ethanol/acetate, intense P-gp labeling of capillary endothelial cells was obtained with all three antibodies (Figure 6D; Table 1). With mdr-(Ab-1), weak P-gp labeling was also seen in astrocytes (Figure 6A), which disappeared by pretreatment with ethanol/acetate (Figure 6D).
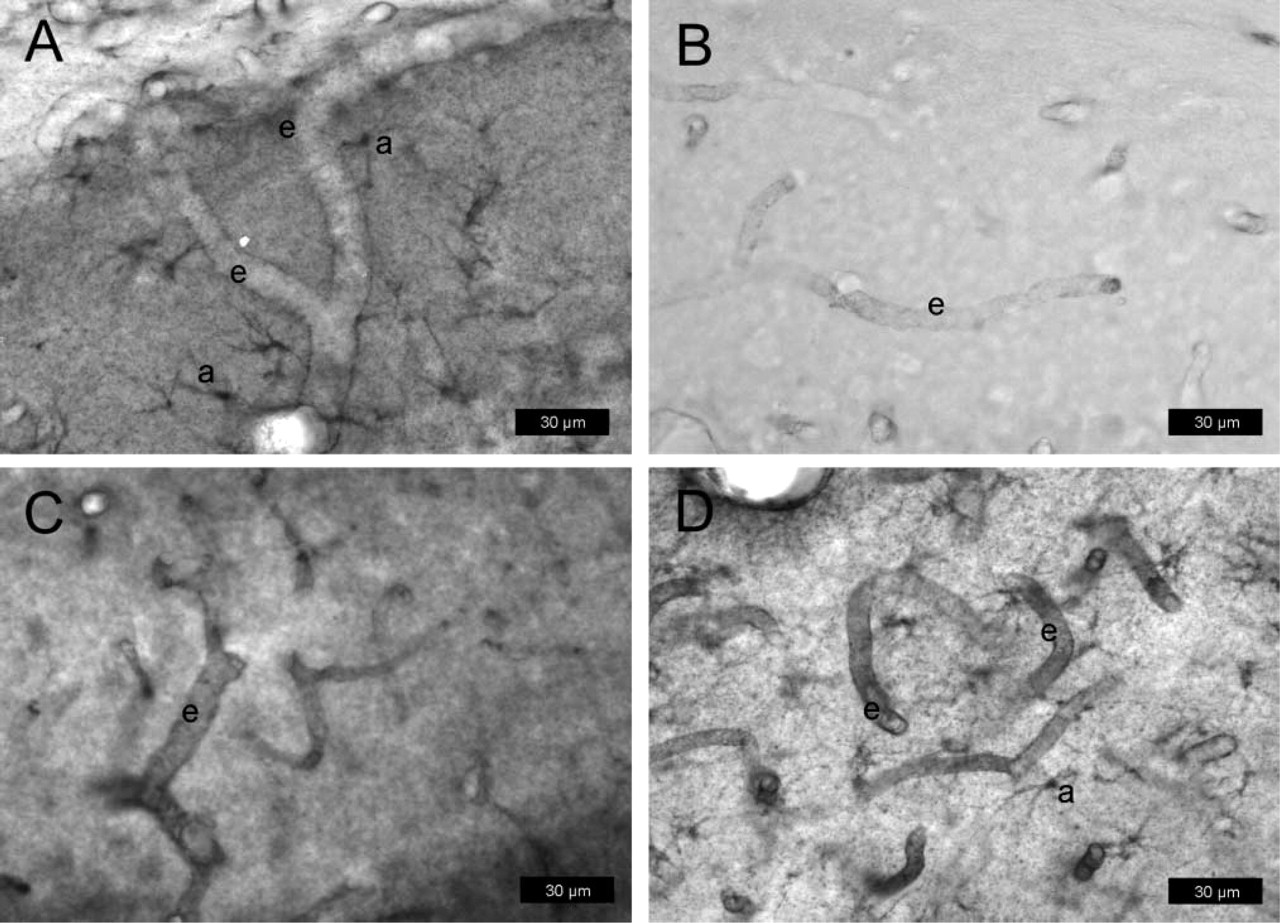
P-gp expression in rat brain determined with protocol #2 (paraformaldehyde-fixed cryosections) after kainate-induced seizures. Sections are from the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody mdr-(Ab-1) was used in
After kainate-induced seizures, astrocytic expression of P-gp was seen with all three antibodies (Figures 7A-7C), whereas only mdr-(Ab-1) detected P-gp also in endothelial cells (Figure 7A). The astrocytic P-gp expression was observed at cell bodies and astrocyte processes within brain parenchyma as well as at astrocytic end-feet terminating on the abluminal membrane of microvascular endothelium. This visual rendering was qualitatively different after pretreatment with ethanol/acetate in that intense P-gp labeling was seen with all three antibodies in endothelial cells, whereas, with the exception of mdr-(Ab-1), the astroglial labeling disappeared (Figure 7D; Table 1). None of the antibodies detected P-gp in neurons under the conditions of this protocol.
A semiquantitative scoring system was used to evaluate whether the qualitative impression that astrocytic P-gp staining was enhanced by kainate-induced seizures could be confirmed by scoring sections from controls and kainate-treated rats in a blinded fashion. As shown in Figure 8, astrocytic P-gp expression (labeled by C219) was significantly enhanced in the CA1 sector of the hippocampus. No significant increase in P-gp staining was seen in the hilus of the dentate gyrus (Figure 8). In a small number of astrocytes, weak P-gp labeling with C219 was also seen in some control sections (Figure 8).
Comparison of Cellular P-gp Localization According to the Three Protocols
As shown in Table 1, in spite of the several differences between protocols and the different antibodies used, none of the protocols allowed simultaneous localization of P-gp in endothelial cells, astrocytes, and neurons. Thus, at least two protocols had to be used to determine the localization of P-gp in these three cell types in the brain. Furthermore, intensity of endothelial staining was much higher in acetone-fixed tissues when compared with paraformaldehyde-fixed tissues, which could be improved by pretreating paraformaldehyde-fixed sections with ethanol/acetate. The finding that endothelial P-gp staining obtained according to protocol #2 was superior to that according to protocol #3, particularly with C219 and H241, indicates that freezing in protocol #2 enhanced endothelial P-gp staining. A general problem of the two protocols with paraformaldehyde was the higher and variable background staining when compared with acetone-fixed cryosections, which excluded the use of computer-assisted image analysis for quantification of staining intensity as previously used for acetone-stained sections (Volk et al. 2004a).
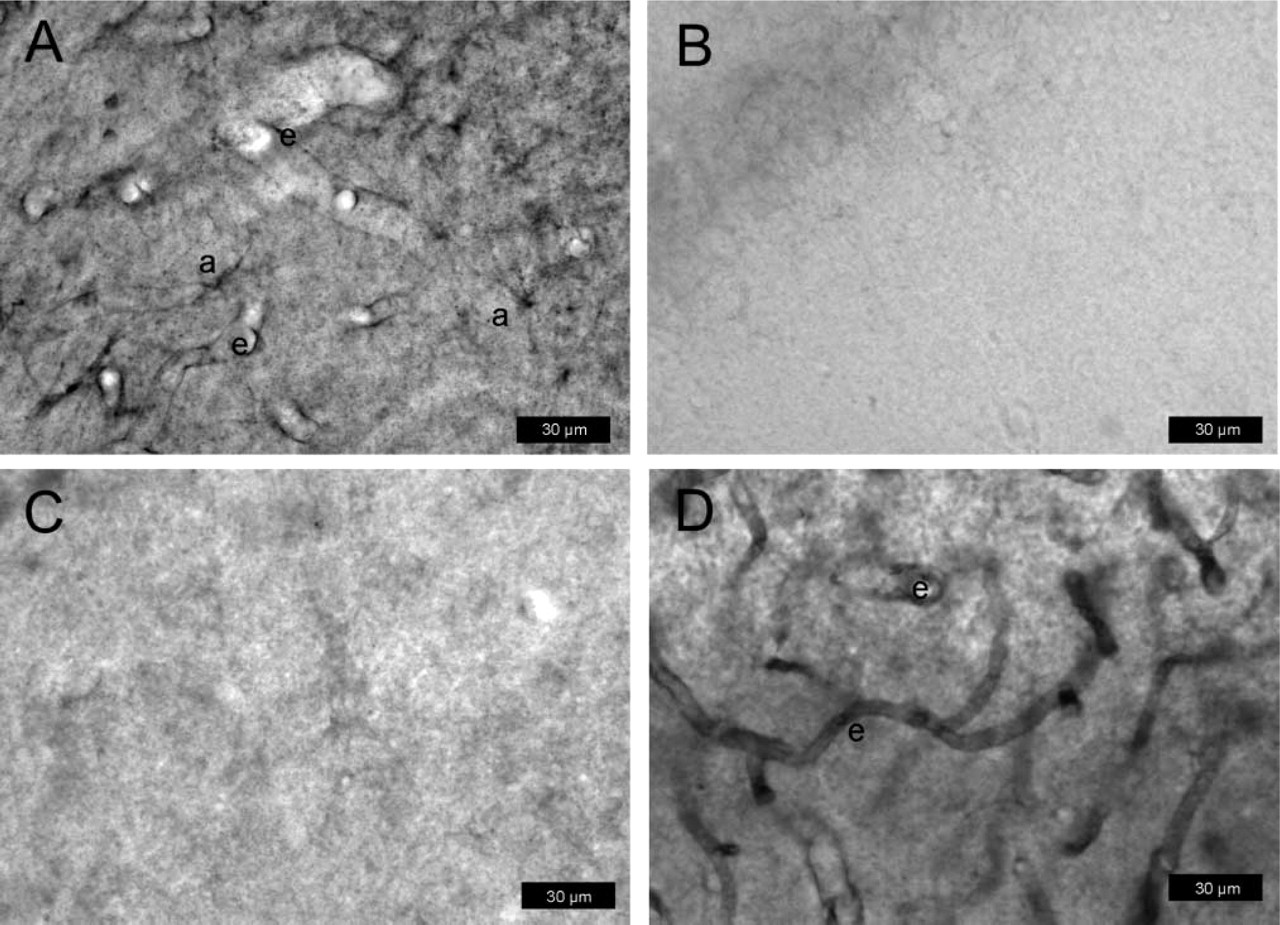
P-gp expression in normal rat brain determined with protocol #3 (paraformaldehyde-fixed vibratome sections). Sections are from the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody mdr-(Ab-1) was used in
Discussion
As reviewed by Larsson (1988), certain criteria are generally recognized as hallmarks of immunocytochemical specificity. These include the use of high-titer antisera at great dilution, total loss of staining when antisera are preincubated with purified antigen, consistent immunolocalization of the protein by at least two, and preferably three, antibodies, and consistent levels of immunoreactivity under variable conditions of fixation, sectioning, and staining. However, it is well known that certain types of fixation, particularly those using formaldehyde, can cause loss of immunoreactivity of many antigens, e.g., due to cross-linking of unrelated proteins to the target antigen, resulting in “masking” of the antigen (Shi et al. 1997; Werner et al. 2000; MacIntyre 2001; Montero 2003). Such loss of immunoreactivity by aldehyde fixation can often be recovered by target unmasking (“antigen retrieval“), using heat or different enzymatic and non-enzymatic reagents such as concentrated acids to restore the immunochemical integrity of the antigen (Shi et al. 1997; Werner et al. 2000; MacIntyre 2001; Montero 2003). As previously described by Thiebaut et al. (1989), the P-gp epitope reactive with C219 is very aldehyde sensitive, so that formaldehyde fixation may dramatically reduce its endothelial localization in brain tissue as compared with acetone-fixed sections. This was also observed in the present study in that endothelial P-gp expression was almost completely lost in paraformaldehyde-fixed sections when using C219 or other P-gp antibodies that recognize the same cytoplasmic epitope as C219. This, however, could be prevented by pretreatment of sections with ethanol/acetate. A likely scenario to explain this finding is that acidic treatment by acetate leads to reversal of antigenic masking (Shi et al. 1997), and that the retrieved P-gp epitope is fixed by the ethanol treatment. In addition, ethanol may enhance penetration of the antibodies to the cytoplasmic P-gp epitope.
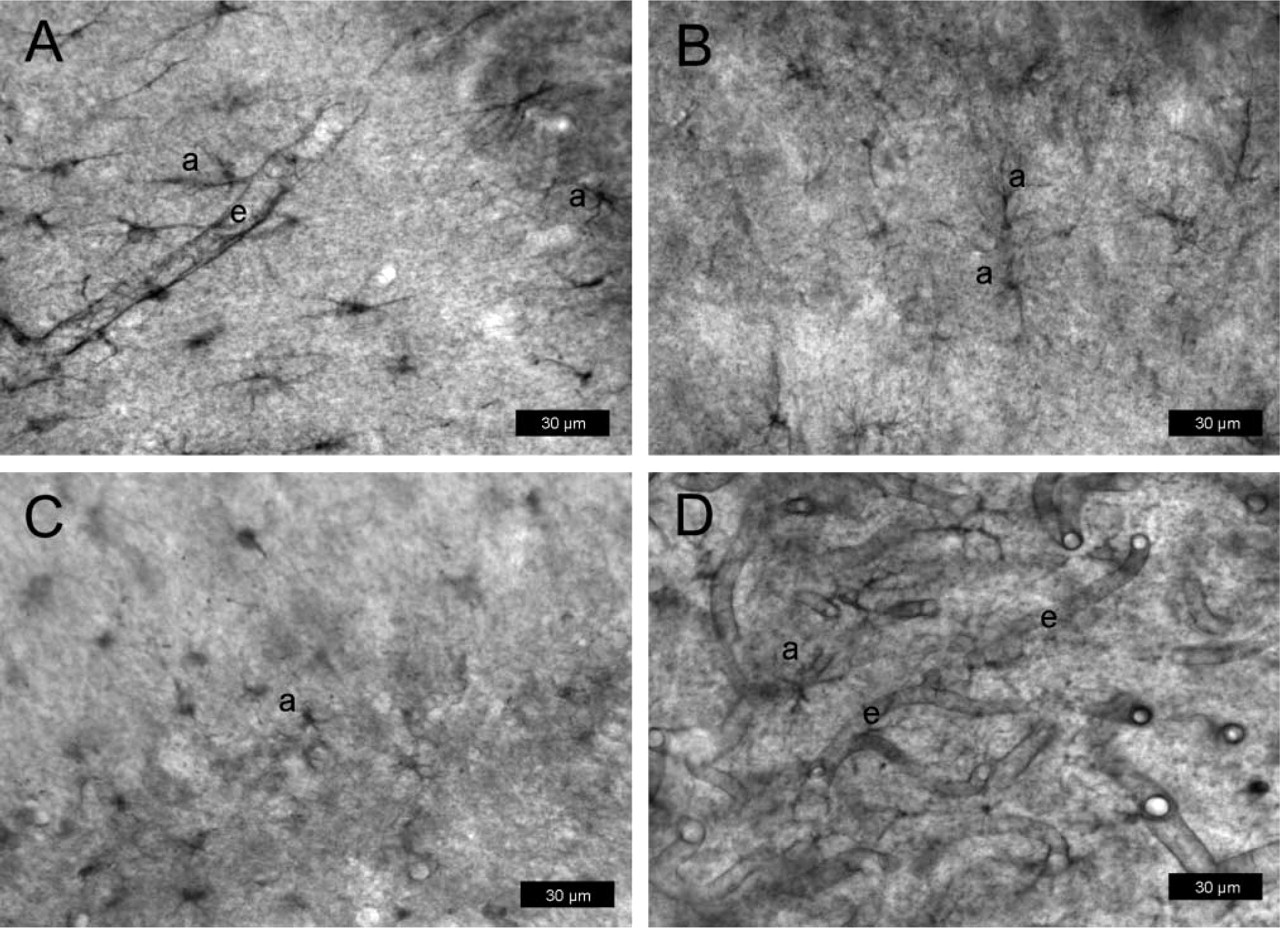
P-gp expression in rat brain determined with protocol #3 (paraformaldehyde-fixed vibratome sections) after kainate-induced seizures. Sections are from the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody mdr-(Ab-1) was used in
Similar to the reduction or loss of endothelial P-gp expression by aldehyde fixation, the neuronal P-gp expression that could be detected in acetone-fixed cryosections from rats after kainate-induced seizures was lost by aldehyde fixation, even when sections were pretreated with ethanol/acetone. In contrast to the endothelial and neuronal localization of P-gp immunoreactivity in acetone-fixed cryosections substantiated with confocal microscopy of double-immunofluorescently stained sections and by determining P-gp-encoding genes (mdr1a, mdr1b) by real-time quantitative RT-PCR (Volk et al. 2004b), an astrocytic expression of P-gp was seen only in aldehyde-fixed sections. This resolves the discrepancy between our previous studies on brain P-gp expression and the study of Zhang et al. (1999), in which paraformaldehyde-fixed vibratome sections were used for immunodetection of P-gp by C219 or mdr-(Ab-1). The loss of endothelial P-gp labeling by aldehyde fixation also explains why Zhang et al. (1999) and other groups (e.g., Lazarowski et al. 2004; van Vliet et al. 2004) observed only weak endothelial P-gp staining with C219 or mdr-(Ab1) in rat brain. Astrocytic P-gp expression after seizures has also been shown in a rat model with electrical SE induction (van Vliet et al. 2004), in rats treated with the convulsant 3-mercaptopropionate (Lazarowski et al. 2004), and in various clinical studies on patients with epilepsy undergoing brain surgery (e.g., Sisodiya et al. 1999; Sisodiya et al. 2002; Aronica et al. 2003). All these laboratory and clinical studies used aldehyde-fixed sections for P-gp immunohistochemistry. Why acetone fixation of cryosections leads to a loss of astrocytic P-gp expression but optimal endothelial and neuronal P-gp expression is unknown at present, but these findings suggest that the optimal fixation conditions vary for different P-gp localizations in the brain. One likely explanation for this observation is that the antigenic epitopes of P-gp might sterically be encumbered in different cell types and thus differ in their accessibility by the antibodies used in this study. Another explanation is that P-gp polymerizes into various complexes (di-, tetra-, hexamers) in different cell types, which variably react after acetone fixation with the antibodies used in this study (Jachez et al. 1994).
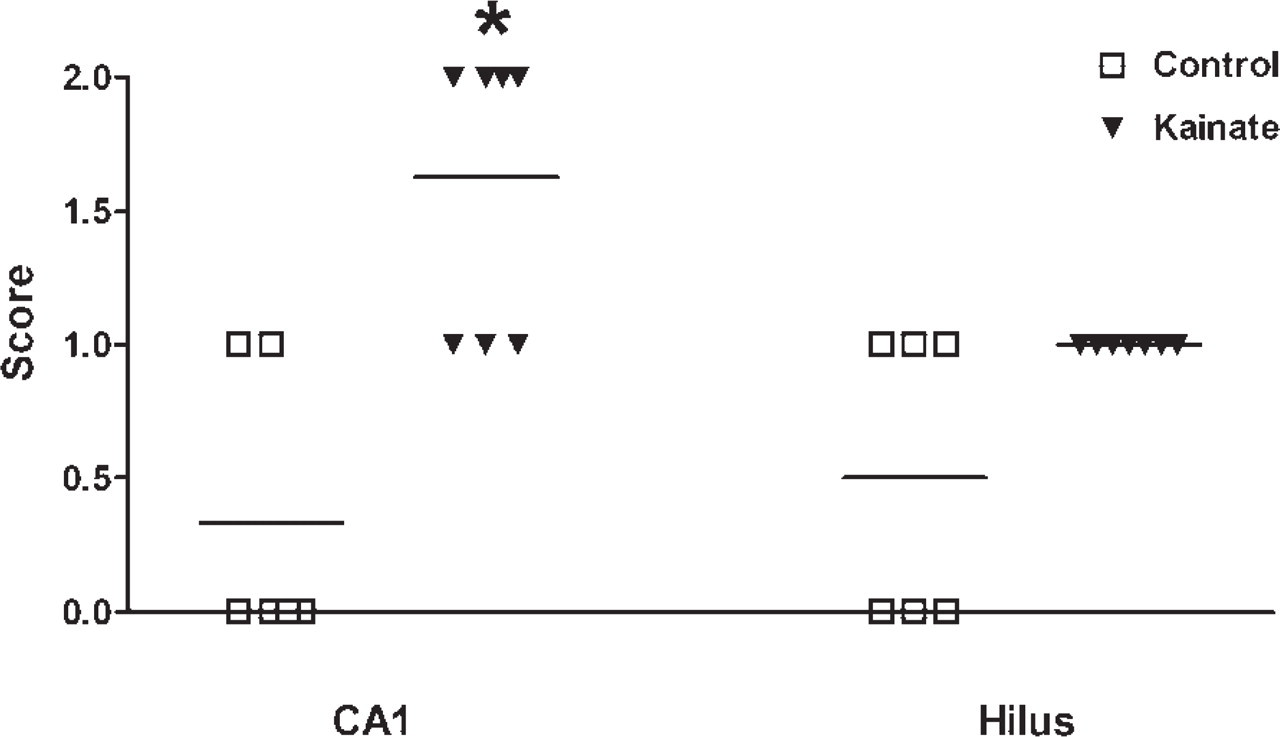
P-gp expression determined with protocol #3 (paraformaldehyde-fixed vibratome sections) in normal rat brain (controls) and rat brain 24 hr after a kainate-induced status epilepticus. The extent of expression was scored in a blinded fashion for sections of the CA1 sector of the hippocampus and the hilus of the dentate gyrus (3.8 mm posterior to bregma). For P-gp labeling, the antibody C219 was used. Each symbol is from one rat. Horizontal lines indicate the mean of values. Astrocytic P-gp expression was significantly enhanced by kainate seizures in the CA1 sector (p=0.0119; indicated by asterisk).
P-gp is not only present in different cell types but also in different subcellular localizations. Although P-gp is predominantly localized in the cell membrane and acts as drug efflux transporter in diverse cell types, more recent studies have shown that it is also expressed in intracellular compartments (Shapiro et al. 1998; Bendayan et al. 2002; Rajagopal and Simon 2003). P-gp is localized in cytoplasmic vesicles, in which P-gp is oriented so that drugs are transported and concentrated in the interior of the vesicles, leading to sequestering drugs away from their subcellular targets (Shapiro et al. 1998; Rajagopal and Simon 2003). In rat cerebral endothelial cells, immunocytochemical analysis employing various P-gp monoclonal antibodies demonstrated the presence of the protein along the plasma membrane, in plasmalemmal vesicles and nuclear envelope (Bendayan et al. 2002). Experimental modulation of endocytic traffic between intracellular and surface P-gp has been shown to change the amount of P-gp on the cell surface (Kim et al. 1997). Furthermore, cytoplasmic factors regulate the membrane orientation of the C-terminal domain of P-gp, so that P-gp expressed in different cell types may have different topological structures modulated by cell-specific cytoplasmic factors (Zhang and Ling 1995; Han and Zhang 1998). This may affect the epitope accessibility by antibodies such as C219 or mdr-(Ab-1) that react with an epitope within the C-terminal domain of P-gp. Within the cell membrane, P-gp may be present in two populations. One of these populations is located in caveolae, i.e., flask-shaped plasma membrane invaginations involved in many cellular events, including transport of macromolecules across cells by transcytosis (Demeule et al. 2000; Jodoin et al. 2003; Schlachetzki and Pardridge 2003). In brain capillary endothelial cells, ~70% of P-gp is localized in caveolae (Jodoin et al. 2003). This caveolar fraction of P-gp cannot be solubilized by the non-ionic detergent Triton X-100, which is a characteristic of proteins associated with the caveolae (Jodoin et al. 2003). Thus, cell-specific differences in cytoplasmatic factors, intracellular vs membrane localization of P-gp, P-gp association with caveolae, and differences in membrane composition are to be expected to lead to a variable presentation and accessibility of antigenic P-gp epitopes, which is a likely explanation for the striking effects of fixation on P-gp localization in different cell types observed in the present study in the rat brain.
One may argue that part of the differences in cellular P-gp localization between different protocols of tissue fixation and processing is due to increased nonspecific binding of the primary antibody. However, the use of three different P-gp antibodies for each protocol makes this an unlikely possibility. Furthermore, for C219 our findings on aldehyde- and acetone-fixed rat brain sections are consistent with those of Thiebaut et al. (1989). Also, Grogan et al. (1990) reported that immunohistological assessment of P-gp by C219 or JSB1 could be optimized by fixation in acetone, and epitope integrity of P-gp has been shown to be better maintained in frozen sections than in formalin-fixed, paraffin-embedded tissues (Toth et al. 1994).
Among the three P-gp-antibodies used in the present study, C219 and mdr-(Ab-1) were about equieffective to stain P-gp in endothelial cells in fresh-frozen tissue with acetone fixation, whereas the labeling with H241 was weak and variable as also observed with the other protocols. Interestingly, the only antibody which consistently labeled P-gp in astrocytes of normal brain was mdr-(Ab-1). By using the same antibody and confocal microscopy, Schlachetzki and Pardridge (2003) recently described the presence of P-gp in both parenchymal and perivascular astrocytes of the normal primate brain, whereas most other groups could not localize P-gp in normal astrocytes of primates or rats by using other antibodies or fixation protocols (Schinkel, 1999; Abbott et al. 2002). In contrast to primates, only few P-gp-antibodies are available for rats, of which C219 is most widely used. This, however, may lead to differences in P-gp localization as shown by the present data.
Irrespective of the effects of fixation and staining variables of P-gp localization and expression, the present data substantiate that P-gp is predominantly localized in capillary endothelial cells in the normal rat brain, but that seizure activity leads to a marked expression of P-gp also in parenchymal and perivascular astrocytes and in neurons. By using computer-assisted image analysis of acetone-fixed sections, we have recently shown that kainate-induced seizures significantly increase P-gp expression in endothelial cells when determined 24 hr after SE (Seegers et al. 2002a, b; Volk et al. 2004a). Furthermore, as shown by the present data, a significant increase of P-gp expression is observed in astroglia, thus confirming the data of Zhang et al. (1999). As previously suggested (Abbott et al. 2002), the increased P-gp expression in astrocytic end-feet surrounding brain capillaries (perivascular astrocytes) in response to seizures may act as a “second line of defense” to support the primary endothelial barrier of the BBB.
The seizure-induced expression of P-gp in parenchymal astroglia and neurons is in line with recent findings in the epileptic human brain from Janigro's group (Marchi et al. 2004). These authors reported that in epileptogenic brain tissue taken during surgical resections in patients with pharmacoresistant epilepsy, a large portion of neurons and astrocytes were P-gp positive. Marchi et al. (2004) suggested that P-gp overexpression in brain capillary endothelial cells may cause reduced AED penetration into the brain, whereas P-gp in parenchymal astroglia and neurons may deserve a role related to protection from apoptotic cell death. This is in line with our recent suggestion for the neuronal P-gp expression seen after kainate (Volk et al. 2004b). P-gp has recently been shown to play a drug efflux-independent role in prevention or inhibition of apoptosis (Johnstone et al. 2000a, b; Pallis et al. 2002). P-gp-mediated efflux of cytokines and signaling lipids and modulation of intracellular pH have been suggested as ways by which P-gp may affect cellular resistance to apoptosis (Pallis et al. 2002). Indeed, these recent findings have challenged the notion that P-gp has evolved merely to efflux xenobiotics out of cells and have raised the possibility that P-gp and related transporter molecules might play a fundamental role in regulating cell survival (Johnstone et al. 2000a, b; Pallis et al. 2002).
In conclusion, the requirements for immunolocalization of P-gp in the rat brain are specific and unusual. As previously reported for human tissues (O'Brien and Cordon-Cardo 1996; Schinkel 1999), tissue fixation and processing may lead to lack of P-gp recognition by obliteration of epitopes in specific cell types. Both freezing and ethanol/acetate treatment of sections seem to expose critical epitopes and thereby improve P-gp immunolocalization. Attention to these details may help to resolve conflicting data regarding the cellular distribution of P-gp and may contribute to future investigations of its functions in the rat brain. Taken together, the present data from rat brain substantiate that under physiological conditions P-gp is predominantly expressed by capillary endothelial cells but, although much weaker, also by parenchymal and perivascular astrocytes, which is in line with the recent study of Schlachetzki and Pardridge (2003) in primate brain. Seizures induce an overexpression of P-gp in these different cell types and, in addition, lead to P-gp expression in neurons. Expression of P-gp in these four different cell types suggests that the functions of P-gp in the brain might be more complex than previously thought.
Footnotes
Acknowledgments
The study was supported by a grant (Lo 274/9-2) from the Deutsche Forschungsgemeinschaft (Bonn, Germany).
We thank Dr. Claudia Brandt for performing the analysis of cell damage after kainate, and Christiane Bartling for technical assistance. We thank Prof. Alexandru C. Stan (Department of Neuropathology, Hannover Medical School, Germany), Prof. Martin Werner (Institute of Pathology, University of Freiburg, Germany), Prof. Ulrich Ebert (Abbott GmbH & Co. KG, GGRP-Neuroanatomy, Ludwigshafen, Germany), and Dr. F. Schlachetzki (Department of Medicine, UCLA) for helpful discussions during preparation of this manuscript.