
Abstract
Several proteins implicated in hormonogenesis of the adrenal cortex have alternatively spliced isoforms, which respond differently to adrenocorticotropic hormone (ACTH). Heterogeneous nuclear ribonucleoproteins A2 and B1 are among the abundant pre-mRNA-binding proteins involved in alternative splicing. We examined the expression of A2 and B1 in normal adrenal cortex and tumors. B1 was variably expressed in the zona fasciculata-reticularis, although A2 was diffusely expressed in the three zones. B1 was more abundant in compact cells than clear cells, and B1 expression was frequent in the zona reticularis, which consists mainly of compact cells. In three kinds of cortical adenomas autonomously producing hormones, B1 was generally overexpressed and there were no significant differences among them. In cortisol-producing tumors, non-tumor parts of the cortex, which were generally atrophic due to low ACTH, had less B1 protein than normal adrenals. These results suggested a correlation between B1 expression and the hormonal activity responding to ACTH. In vitro ACTH stimulation induced a biphasic expression of B1 in an H295R cortical carcinoma cell line, and it paralleled hormonogenesis. Conclusively, B1 expression varied in relation to the hormonal activity responding to the ACTH, and it may provide a key to elucidating the splicing mechanisms involved in hormonogenesis.
T
The role of alternative splicing in responsiveness to ACTH is important (Leo et al. 2002). In cortical cells, ACTH stimulation is transduced mainly through a cyclic AMP-dependent signaling pathway, which regulates the transcription of target genes that have a consensus cAMP response element (CRE) in their promoters. CRE-binding transcription factors, such as CREM (CRE modulator) and CREB (CRE-binding protein), play important roles in the final step of this pathway (Lalli and Sassone-Corsi 1994). CREM and CREB function either as repressors or as activators, which are controlled by alternative splicing or alternative transcriptional start sites of these genes (Yin et al. 1995). Furthermore, ACTH, angiotensin II receptor, and some enzymes related to hormone production have several splicing variants that act differently (Yang et al. 1995; Martin et al. 2001; Rotzer et al. 2001). These variants suggest that alternative splicing is an important regulating mechanism in hormonogenesis, although this mechanism is poorly understood.
The heterogeneous nuclear ribonucleoproteins (hnRNPs) are a family of nucleic acid-binding proteins and comprise 24 different polypeptides termed hnRNPA1 through hnRNPU (Pinol-Roma et al. 1988). Although the functions of hnRNPs are diverse, they are implicated in most stages of mRNA metabolism, including transcription, splicing, nucleocytoplasmic transport, stability, and localization (Krecic and Swanson 1999). The basic A/B group of hnRNPs, such as A1 and A2/B1 proteins, antagonizes the in vitro alternative splicing activity of splicing factors SF2/ASF or SC35 and influences 5′ splice-site selection and exon skipping (Mayeda et al. 1994). Previously, we reported that A2 and B1 proteins show an interesting expression pattern in murine adrenals (Kamma et al. 1995). The A/B group of hnRNPs is being considered as an important candidate for regulation of hormonal function in alternative splicing. However, the adrenal expression of these has not yet been studied. On the other hand, there have been many reports that the B1 protein is overexpressed specifically in tumors of several organs (Hamasaki et al. 2001; Zhou et al. 2001).
The final goal of our study is to elucidate the alternative splicing mechanism, which is related to the adjustment of cytodifferentiation and steroidogenesis regulation in the adrenal cortex. In this study, the histological expression of the A2 and B1 proteins was examined in the adrenal cortex and functional adrenocortical tumors. Interesting expression patterns were found related to the functional differentiation of the adrenal cortex.
Materials and Methods
Patients and Tissue Samples
The research protocol was approved by the Ethics Committee of the University of Tsukuba, and all samples were obtained after the patient's approval. Seven normal adrenal samples were obtained from total nephrectomy materials from patients who had renal cell carcinoma without adrenal involvement. Seventeen samples of three kinds of functional adrenocortical adenomas were surgically removed from 10 patients with Conn's syndrome (aldosterone-producing tumors, aldosteronomas), 6 patients with Cushing's syndrome (cortisol-producing tumors), and a case with adrenogenital syndrome (testosterone-producing tumor). For the immunohistochemical study, tissue samples were fixed with 4% phosphate-buffered formaldehyde and paraffin embedded. For Western blotting, tissue samples were frozen and stored in liquid nitrogen. In the adrenocortical adenoma cases, except the case with adrenogenital syndrome, samples were obtained from both tumor and non-tumor parts of the adrenal from the same patient. In the case with the adrenogenital syndrome, the non-tumor part was unavailable as an SDS sample.
Cell Cultures
A human adrenocortical carcinoma cell line, H295R, was purchased from ATCC (Rockville, MD). It was grown in DMEM/Ham's F-12 (1:1) supplemented with 2.5% fetal calf serum, 2% Ultroser G, 2 mmol/L glutamine, 5 μg/ml insulin, 5 μg/ml transferrin, 5 ng/ml selenium, 5 ng/ml linoleic acid, 1.25 mg/ml bovine serum albumin, 50 U/ml penicillin, and 50 mg/ml streptomycin (Sigma; St Louis, MO). HeLa cell line (JW 36) was utilized as a control. Both cells were cultured at 37C in a 5% CO2 incubator.
Preparation of Antibodies and Immunohistochemistry
Anti-hnRNP A2 and B1 monoclonal antibodies, 4G8 and 2B2, were generated previously in our laboratory (Kamma et al. 2001). All antibodies were in the form of ascites fluid or protein-A-purified antibody, and their titers were adjusted by normalization to immunostaining of HeLa cells.
Paraffin tissue sections were cut into 4-μm-thick slices, deparaffinized, and heated in 0.01 M citrate buffer (pH 6.0) by microwave irradiation for antigenic retrieval. The sections were preincubated with 3% BSA in phosphate-buffered saline (PBS) to diminish the background and then incubated with antibodies for 1 hr at room temperature. Bound antibodies were detected by a peroxidase-labeled polymer-conjugated anti-mouse antibody (EnVision+; DAKO, Fort Collins, CO), and reacted with 0.025% 3,3′-diaminobenzidine tetrahydrochloride. Hematoxylin was utilized for counterstaining. To objectively estimate the expression pattern of the A2 and B1 proteins, the positive cells for 4G8 and 2B2 were morphometrically analyzed as described previously (Wu et al. 2002) in the seven normal adrenal tissues. The results were presented as the mean ± SD. Statistical analysis was evaluated by Student's t-test. Differences were considered significant at p<0.05.
Gel Electrophoresis and Western Blotting
The frozen samples of normal adrenal tissues and adrenocortical adenomas were crushed and homogenized in sodium dodecyl sulfate (SDS) sample buffer. The protein concentration of each sample was measured by a protein assay kit (Bio-Rad; Hercules, CA). Protein from each sample (50 μg) was separated by electrophoresis on 12.5% SDS-polyacrylamide gels. Proteins were transferred to nitrocellulose (PROTRAN; Schleicher and Schuell, Keene, NH) and reacted with monoclonal antibodies 4G8 and 2B2. Bound antibodies were detected with peroxidase-conjugated anti-mouse antibodies from sheep (Amersham; Buckinghamshire, UK) and the ECL system (Amersham). To quantitatively estimate the expression of A2 and B1, SDS tissue samples were adjusted to contain 50 μg of protein/lane, and the blotting result images were analyzed by a densitometer (GS-700 Imaging Densitometer; Bio-Rad).
ACTH Stimulation of H295R Cells
H295R cells were washed twice with PBS and then cultivated in a medium containing ACTH (Sigma), ranging from 0 M to 10−7 M, or forskolin (Sigma), ranging from 0 μM to 20 μM. The cells were harvested from 0 to 72 hr at 12-hr timepoints, counted, and prepared at 3 × 106 cells with 300-μl SDS sample buffer for each timepoint. Concurrently, the culture supernatants were collected for estimating hormone production by radioimmunoassay, including cortisol and dehydroepiandrosterone sulfate (DHEA-S), which is stably produced from the H295R cells and is recognized as an adrenal sexual steroid precursor (Gell et al. 1998). The experiments were performed three times.
Results
Variable Expression of A2 and B1 Proteins in the Adrenal
Immunohistochemically, we studied the expression patterns of A2 and B1 proteins in the adrenal. Monoclonal antibodies 4G8 and 2B2 detect A2 and B1 proteins, respectively, work well in human adrenal tissue, and demonstrate distinctive nuclear staining. The staining intensities of A2 and B1 proteins were variable in the cortex, although they were weaker than in the medulla. A2 and B1 proteins tended to be more strongly detected in the zona reticularis than in the zona fasciculata and glomerulosa. This tendency was quite obvious, especially for B1 protein in comparison to A2 protein (Figure 1). Furthermore, B1 expression was different between cortex cell types, and B1 protein was more abundantly expressed in compact cells than in clear cells (Figure 1). The nuclear-positive ratios for A2 and B1 proteins were calculated and statistically compared among cortical zones and the medulla, and between the two cell types, compact and clear cells, that are distributed in the zona fasciculatareticularis. In the cortex, B1 expression varied more obviously among cortical zones than did A2 protein expression. B1 protein was highly expressed in the zona reticularis (70%), but was low in the zona glomerulosa and fasciculata (29% and 27%, respectively). On the other hand, the positive ratios for A2 protein were 64%, 68%, and 70% in the three cortical zones, respectively, and they were not significantly different (Figure 2). In the medulla, the positive ratios for both A2 and B1 proteins were high, ~85%. Furthermore, in the zona fasciculata-reticularis, compact cells expressed the B1 protein more frequently (78%) than did the clear cells (32%). This expression pattern was significantly different, suggesting that B1 expression is related to cortical cell activity.
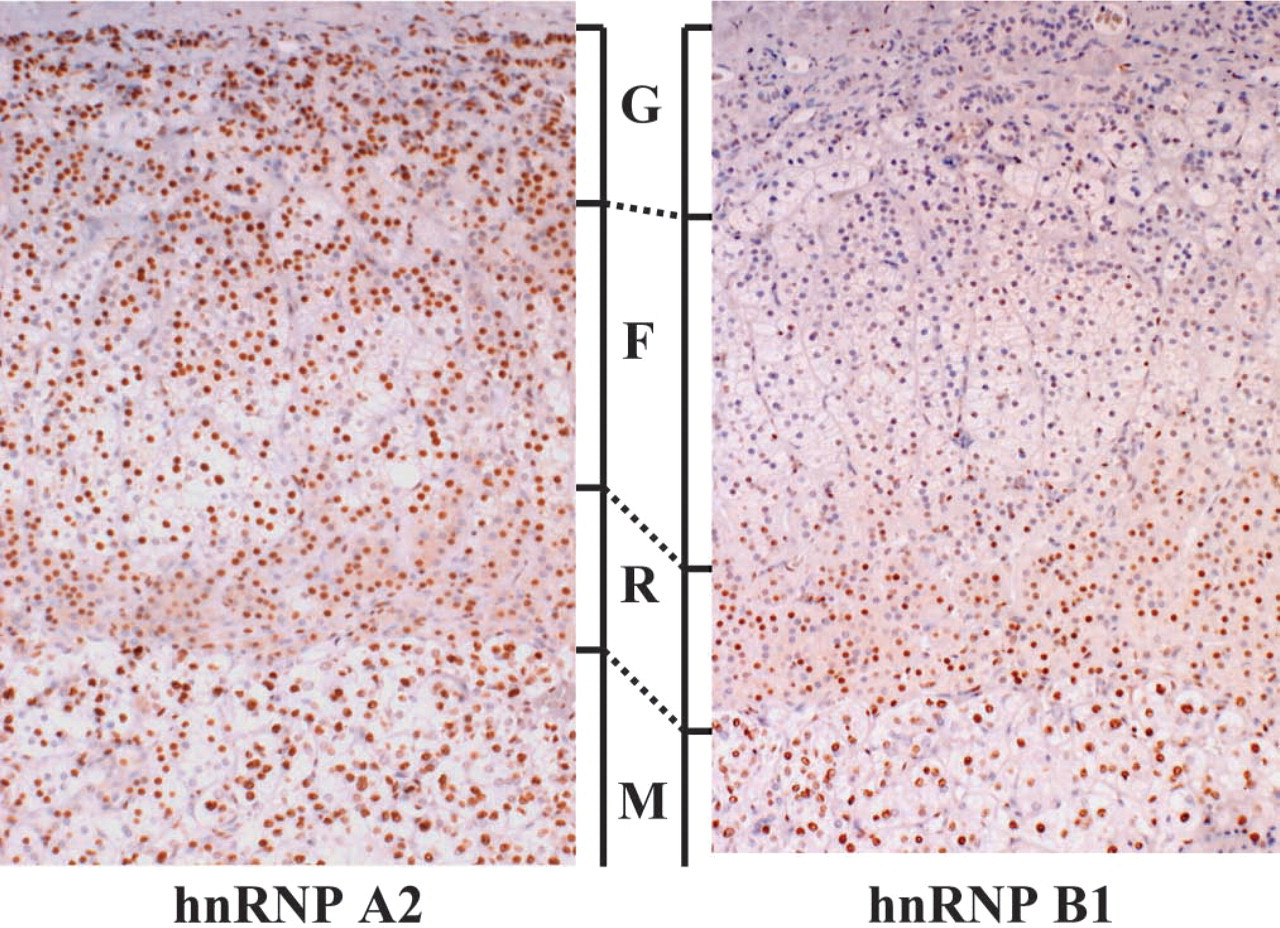
Expression of A2 protein (left panel) and B1 protein (right panel) in the normal adrenal. Proteins A2 and B1 are more strongly detected in the zona reticularis, which is predominantly composed of compact cells, than in the zona fasciculate, which is predominantly composed of clear cells and glomerulosa. G, zona glomerulosa; F, zona fasciculate; R, zona reticularis; M, medulla. Original magnification, X40.
Expression of B1 Protein in Functional Cortical Adenomas
B1 expression in the three kinds of adrenocortical adenomas that produce aldosterone, cortisol, and testosterone was immunohistochemically studied and compared with B1 expression in the normal cortex (Figure 3). B1 protein was overexpressed in all tumors, and it was generally higher than in the corresponding zones in the normal cortex, although the expression patterns were somewhat similar among the three adenomas. The androgen-producing adenoma, which was composed only of compact cells, showed homogeneous expression throughout the tumor, and the nuclear-positive ratio was ~100%. Aldosteronomas were relatively lower in the positive ratio, and the average was ~70%. The average of the cortisol-producing tumors was ~85%. The positive ratios for B1 were analyzed between the compact and clear components in aldosterone- and cortisol-producing adenomas. In contrast to normal cortical cells, B1 expression was higher in the clear-type tumor cells than in the compact-type ones. Furthermore, we investigated non-tumor parts of adrenal cortex in functional adenoma cases (Figure 4). Interestingly, expression patterns of B1 protein were different between aldosteromas and cortisol-producing adenomas. In aldosteromas, the non-tumor part was not atrophic and showed the same level of B1 expression as the tumor part, whereas in cortisol-producing tumors, the non-tumor parts were atrophic and showed a lower expression level of B1 protein than did the tumor parts.
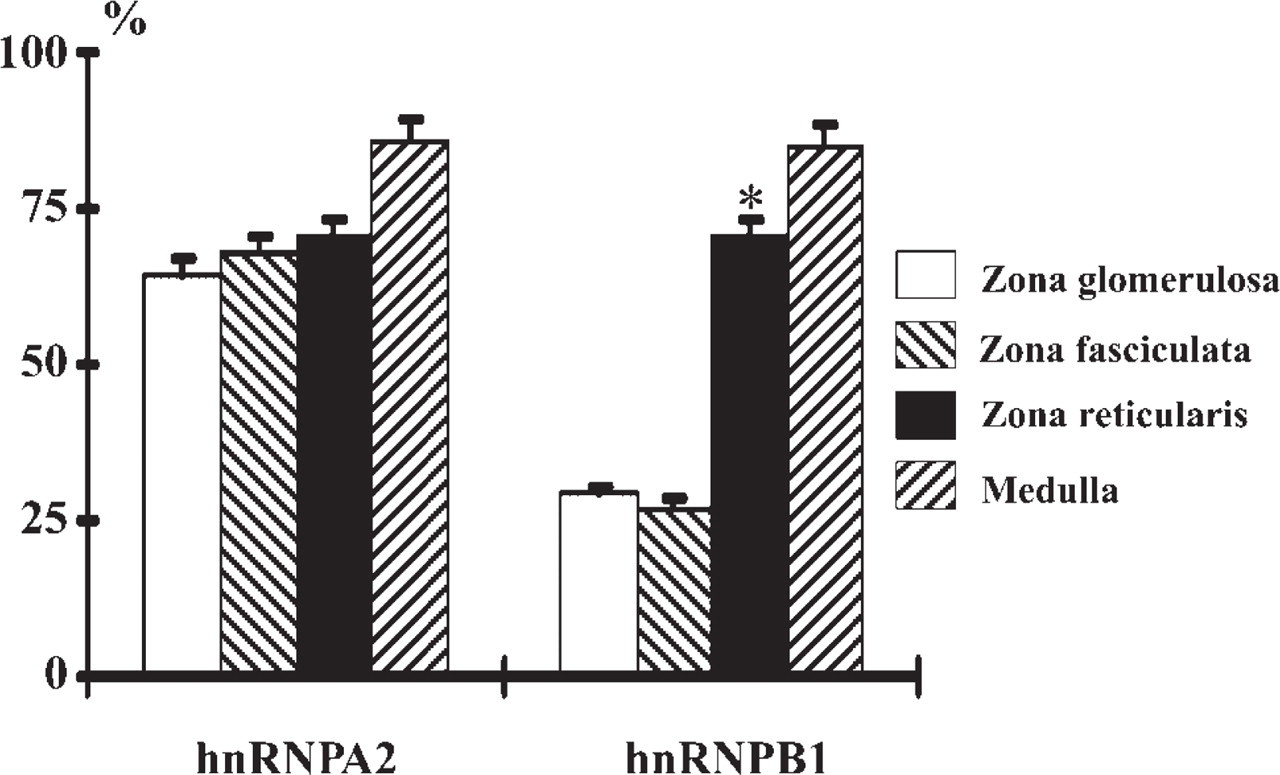
Expression ratios of A2 and B1 proteins in three cortical zones. The positive ratio of B1 protein is significantly higher in the zona reticularis than in the zonas glomerulosa and fasciculata; however, the difference is not significant in that of A2 protein. Error bars indicate SD. ∗Significantly different at p<0.05.
Quantitative Analysis of hnRNPA2 and B1 Expression in Adrenal
Western blotting was performed to confirm A2 and B1 proteins detected by 4G8 and 2B2 antibodies and to estimate their expression amounts in H295R cells derived from a human adrenocortical carcinoma, compared with HeLa cells as control. Antibodies 4G8 and 2B2 detected 36 kD and 38 kD bands corresponding to A2 and B1 in H295R cells as well as in HeLa cells. The relative expression amount of B1 to A2 protein was higher in H295R cells than in HeLa cells (Figure 5). Next, we comparatively estimated the amount of B1 expression between the normal adrenal tissue and tumor tissue (Figure 6). The adrenocortical adenomas clearly displayed strong signals compared with normal adrenal tissues and the non-tumor part of the same patient, although the intensities were variable among the three kinds of adenomas. Interestingly, B1 expression in the non-tumor parts was different between aldosterone- and cortisol-producing adenomas. That of aldostromas was more abundant than in normal adrenals, whereas that of cortisol-producing tumors was less abundant than in normal adrenals.
A2 and B1 Expression in H295R Cells Stimulated by ACTH
A2 and B1 protein expression was quantitatively analyzed 72 hr after the addition of various concentrations of ACTH to the H295R cell culture media. B1 expression decreased in a dose-dependent manner, and it was significantly lowered by the addition of 10−7 M ACTH, which is 104 times higher than the physiological concentration. In contrast, A2 expression did not show a significant difference (Figure 7).
The hormonal activity of H295R cells stimulated by 10−7 M ACTH was estimated by a time course that monitored intracellular secreted cortisol and DHEA-S, which is an androgen precursor. DHEA-S gradually increased after ACTH addition and reached a plateau, because DHEA-S seldom decomposed in the culture supernatant and, thus, accumulated there (Figure 8). The cortisol failed to show such a noticeable change as DHEA-S, although it gradually increased in the culture supernatant. Thus, H295R cells produced intracellular cholesterol and DHEA-S in a biphasic pattern after ACTH stimulation. A2 and B1 protein expression was also examined in a time course after ACTH stimulation by quantitative Western blotting (Figure 9). B1 expression rapidly increased right after ACTH addition, approximately doubling the original concentration in 24 hr, which was earlier than the cholesterol accumulation. It gradually decreased and then decreased to a level lower than the initial concentration after 72 hr of incubation. In contrast, A2 protein expression gradually decreased. The expression, especially of B1, interestingly paralleled DHEA-S production.
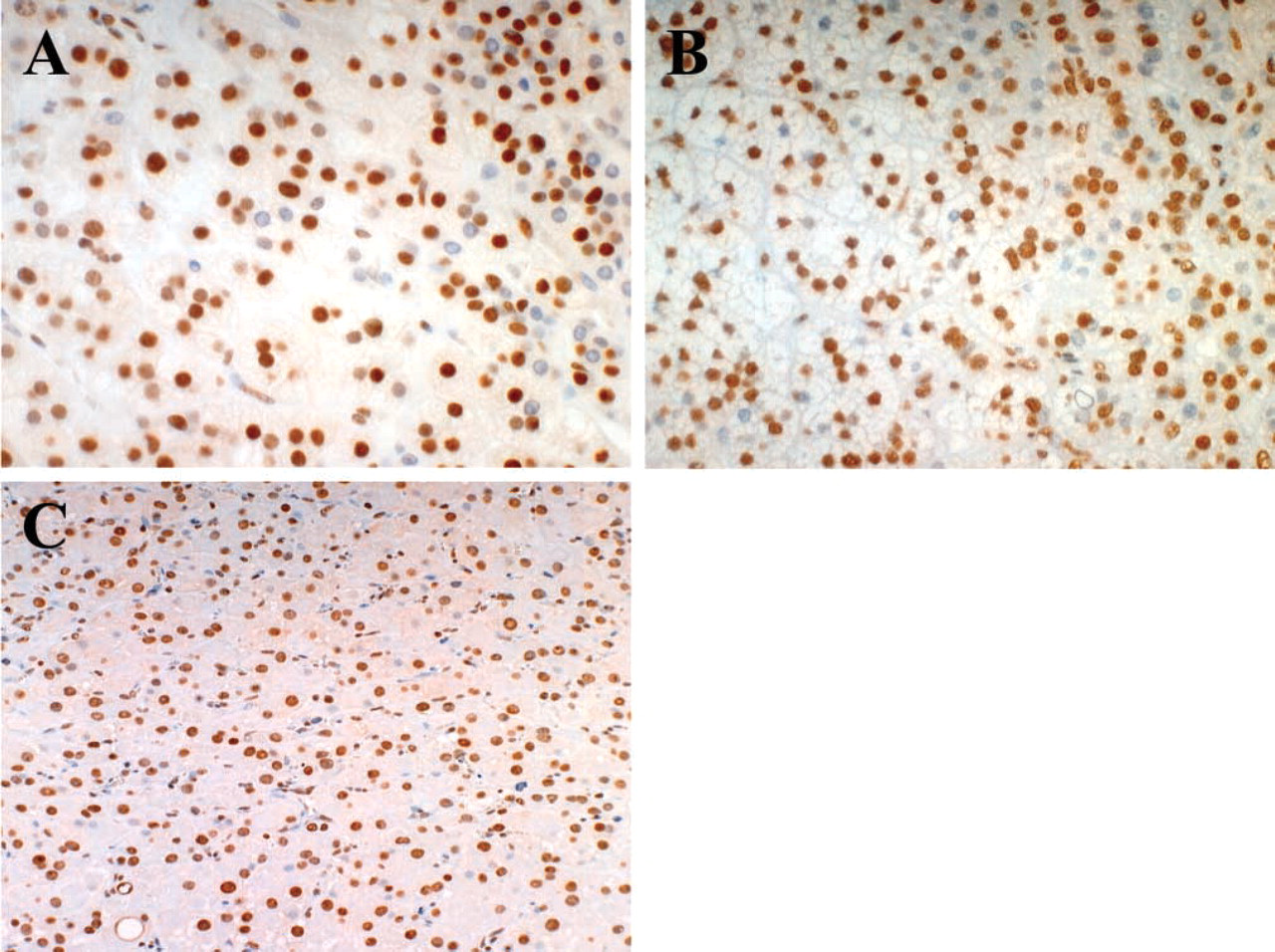
Expression of B1 protein in functional adenocortical adenomas.
Discussion
A previous immunoelectron microscopic study suggested that the composition of the hnRNPs changes according to hormone-producing activity in the adrenal cortex (Malatesta et al. 1995). Our results demonstrated that the B1 protein was variably expressed and higher in the zona fasciculata-reticularis than in the zona glomerulosa, whereas A2 protein was abundantly expressed in three zones, and there was no significant difference in expression among the three zones. Regarding expression in particular cell types, B1 protein is expressed more frequently in compact cells than in clear cells in the zona fasciculata-reticularis. It is reasonable that B1 protein is expressed most frequently in the zona reticularis because it consists mainly of compact cells. It is thought that the clear and compact cells have a differential response to ACTH, in which the compact cells have high hormone-producing activities because they are rich in smooth ER and mitochondria. These findings suggested the possibility that the alteration of B1 expression is related to the hormonal activity of cortical cells in normal adrenals. B1 expression should increase in response to ACTH stimulation. However, B1 expression does not simply reflect the particular hormone that is present. B1 expression was compared among three adrenocortical tumors, which autonomously produce aldosterone, cortisol, and androgen adenomas and express large amounts of B1 protein in comparison with normal adrenals. There was no significant difference in B1 expression among the three cortical hormones, indicating that there is no relationship between B1 expression and cortical hormone. B1 expression in cortical tumor cells seems to also reflect the high hormone-producing activity that is independent of ACTH stimulation. B1 protein is overexpressed in tumors of other organs, such as lung (Hamasaki et al. 2001; Zhou et al. 2001), suggesting that B1 expression is related to tumorigenesis and may be involved with adrenocortical adenomas.
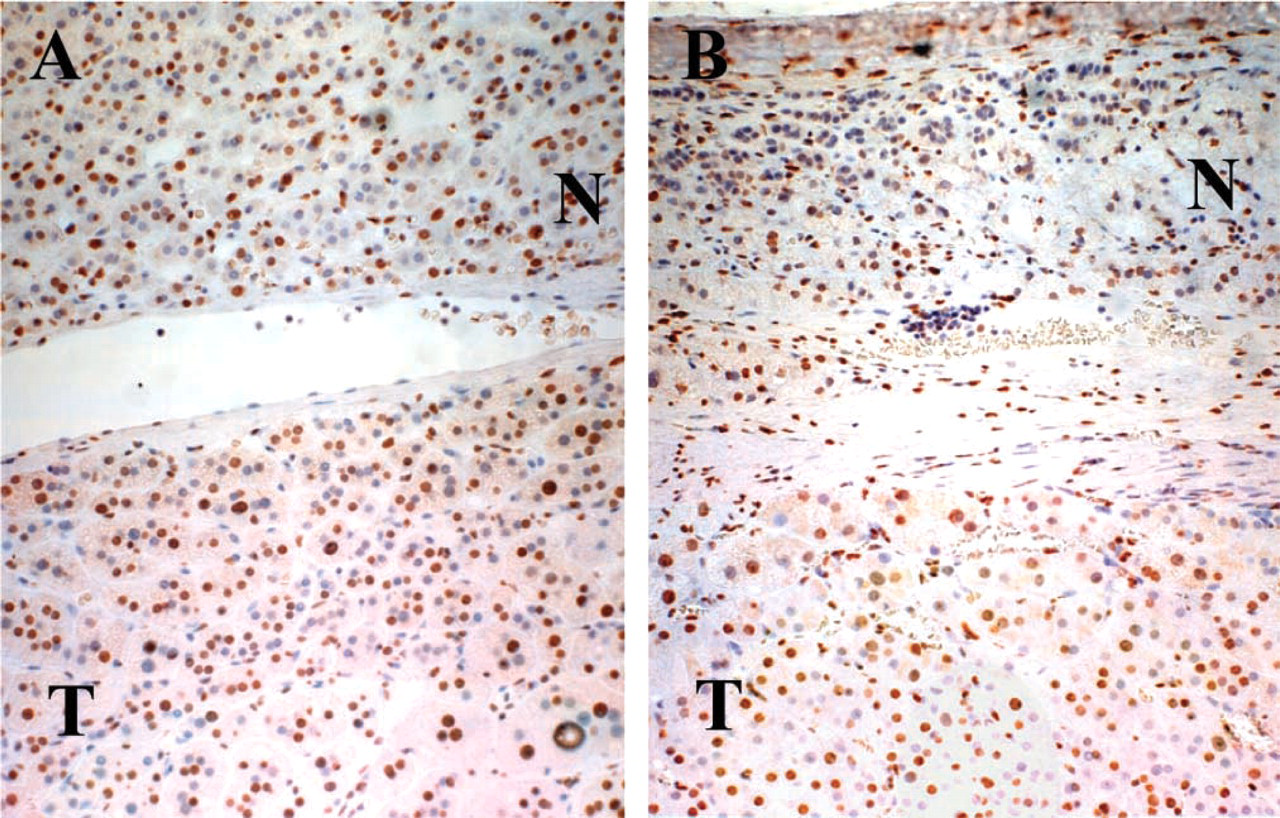
Expression of B1 protein in non-tumor part in functional adenoma cases.
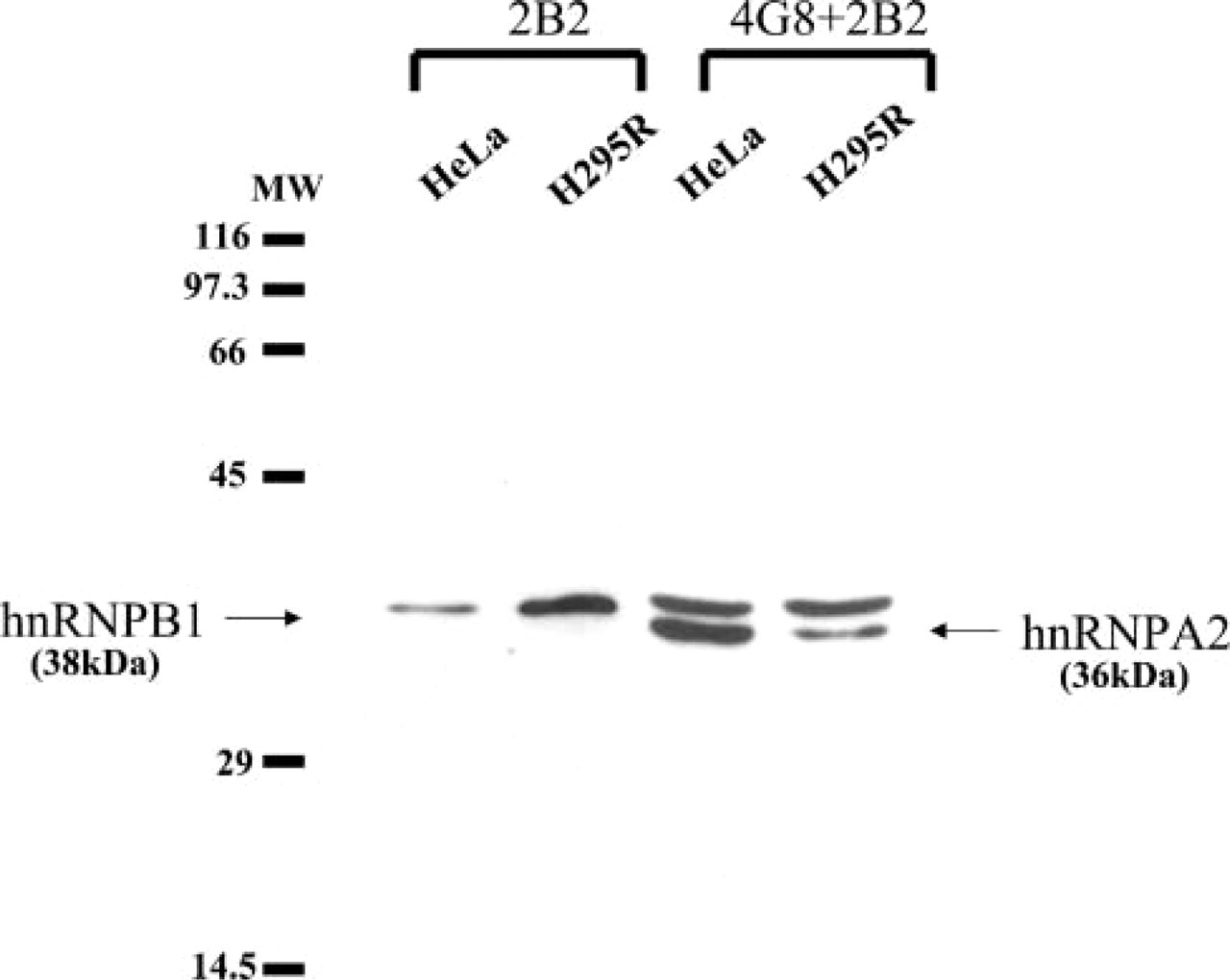
Western blotting of A2 and B1 proteins in H-295R adrenocortical carcinoma cells with 4G8 and 2B2 antibodies. B1 proteins are detected by 2B2 as a 38-kD single band in both H-295R and HeLa cells and is more abundant in H-295R cells. The relative ratio of B1 to A2, which is simultaneously detected by a mixture of 4G8 and 2B2 antibodies, is obviously higher in H295R cells than in HeLa cells.
Interestingly, B1 expression was higher in clear-type tumor cells than in compact-type tumor cells, which was reverse to B1 expression in the normal adrenal cortex. Electron microscopic studies of cortical tumors have revealed that clear-type tumor cells have abundant smooth endoplasmic reticulum and mitochondria (Eto et al. 1979; Gazdar et al. 1990). This suggests that clear-type tumor cells have high hormone-producing activities, unlike normal cortical cells.
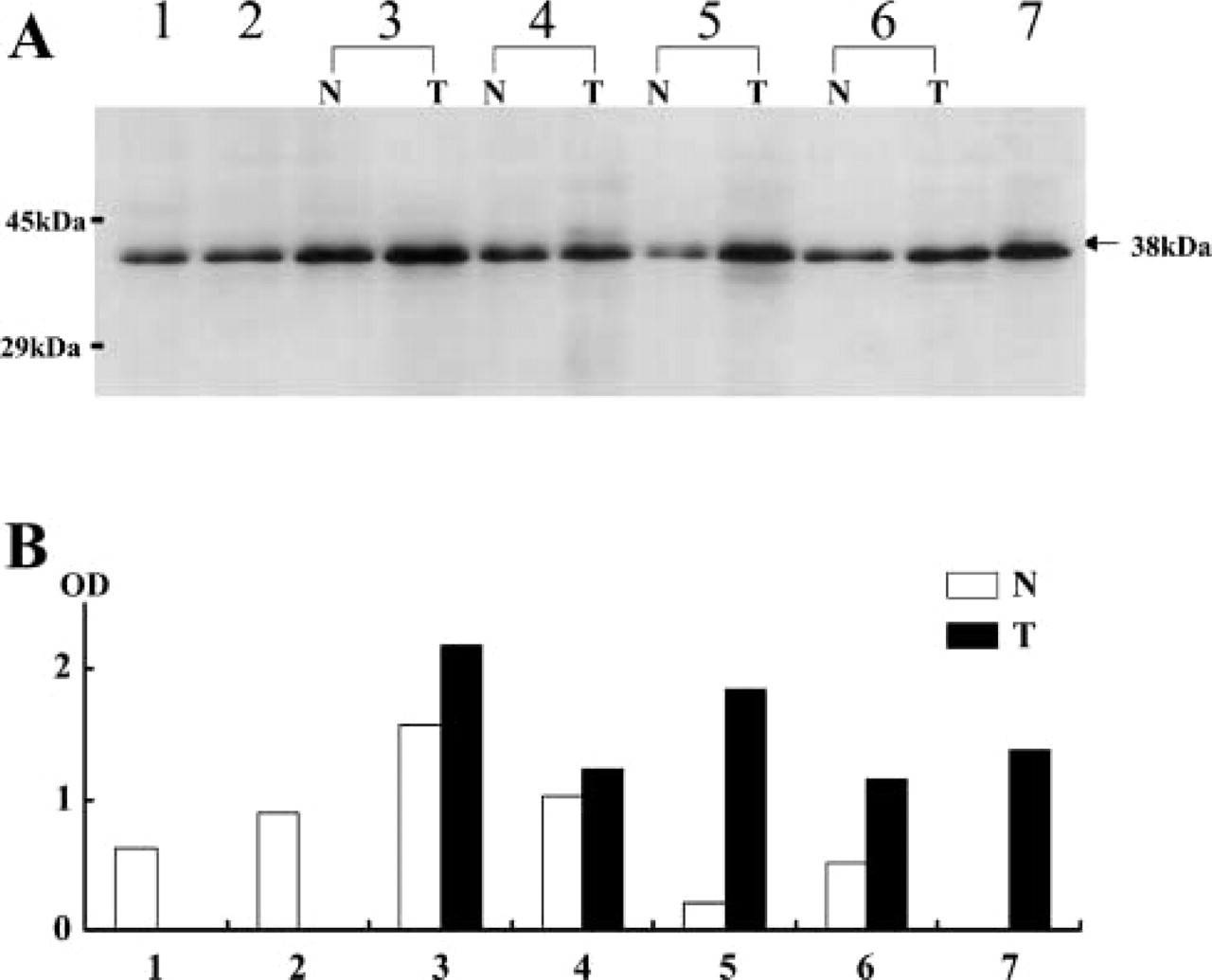
Quantitative analysis of B1 protein expressed in tumor (T) and non-tumor tissues (N).
In addition, non-tumor parts of the adrenal cortex in cortisol-producing tumors had less B1 protein than in normal adrenals, whereas those in the aldosterone-producing tumors had more B1 protein. The cortical cells in vivo function under a complicated feedback mechanism, which is controlled mainly by ACTH secreted from the pituitary gland. In patients with cortisol-producing tumors, the ACTH serum level is usually low and the non-tumor part is histologically atrophic, indicating low hormonal activity. On the other hand, in patients with aldosterone-producing tumors, ACTH secretion is not suppressed and the non-tumor parts are normal or hyperplastic (Murakoshi et al. 1995). This also suggests the correlation of B1 expression in the cortical cells to the hormone-producing activity responding to ACTH.
A simple experimental model of cortical hormono-genesis is necessary to show the relationship between B1 expression and ACTH stimulation. The H295R cell line has receptors for ACTH and retains the ability to produce mainly androgen in response to ACTH (Gazdar et al. 1990; Mountjoy et al. 1994), although the number of receptors is less than in normal cortical cells. ACTH increases intracellular cAMP production and hence activates the PKA pathway and CRE-binding transcription factors. In this experiment, H295R cells were utilized to reproduce the adrenocortical hormonogenesis controlled by ACTH. After ACTH stimulation, H295R cells showed biphasic patterns of DHEA-S secretion. This biphasic reaction to ACTH has been reported previously (Jefcoate et al. 1987; Le Roy et al. 2000) and was explained by the biphasic up-and downregulation of ACTH receptors in H295R cells (Fassnacht et al. 1998, 2000). This was reproduced also in a forskolin stimulation experiment (data not shown). In this experiment, B1 also showed a dramatic biphasic expression pattern that almost paralleled that of DHEA-S secretion, whereas the change of A2 was less dramatic, although it showed the same biphasic pattern. The maximum point of the biphasic change of B1 expression apparently precedes hormonal production. After prolonged stimulation (72 hr) with ACTH (Figure 7), H29R cells showed a notable decrease of B1 expression in a dose-dependent manner. However, A2 did not exhibit obvious change. The decrease of B1 may reflect the low responsiveness to ACTH, which has been observed under a distinct experimental condition of prolonged ACTH stimulation, representing an in vivo shock state (Slawik et al. 2004). B1 protein may be involved in an intercellular feedback mechanism via the cAMP pathway during chronic continuous stimulation with ACTH. A2 and B1 are transcribed from an identical gene (Kozu et al. 1995). The change of the ratio between the A2 and B1 proteins may be caused by alternative splicing or posttranslational mechanisms. Preliminary results of quantitative RT-PCR in H295 cells stimulated by ACTH showed no apparent change in the A2/B1 mRNA ratio (data not shown). It is assumed that the dissociation between the A2 and B1 proteins is caused by differences in posttranslational metabolism.
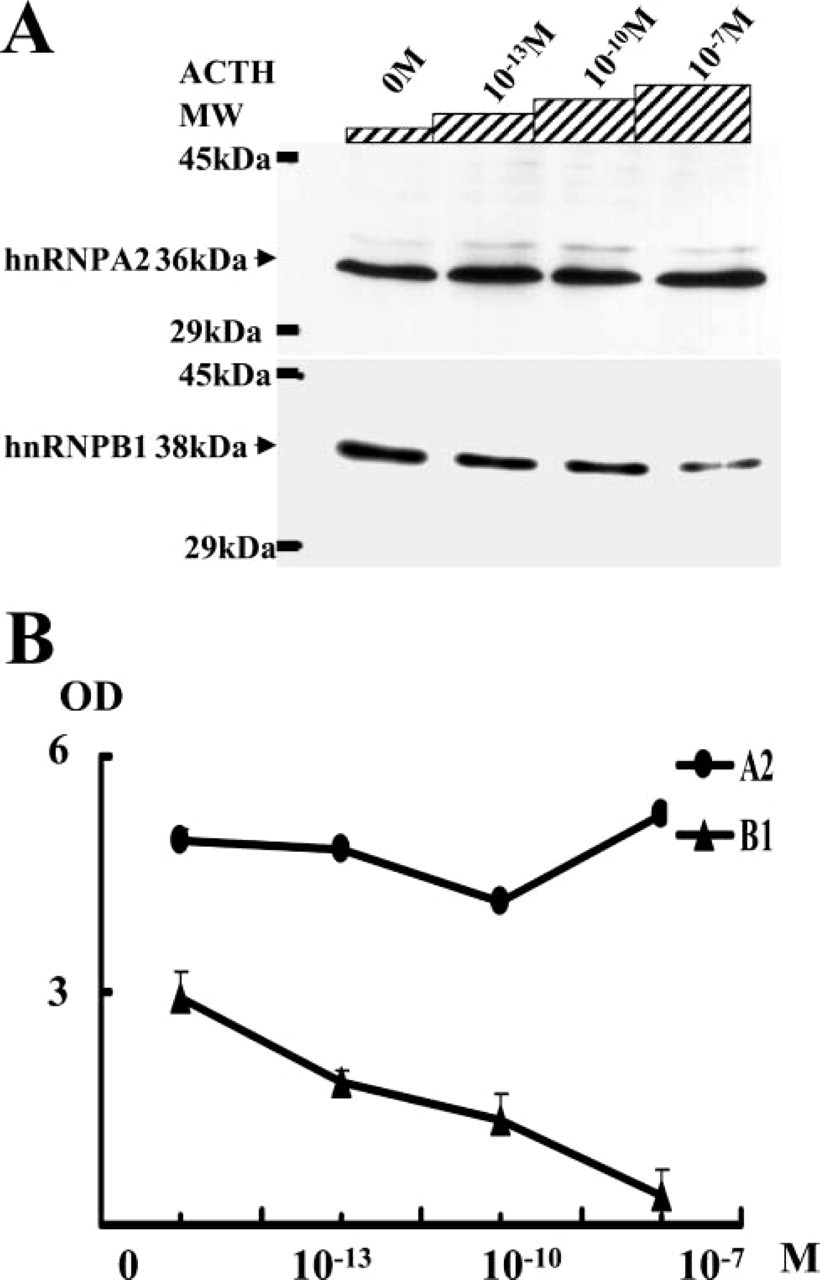
Dose-dependent expression of A2 and B1 proteins in H295R cells stimulated by ACTH.
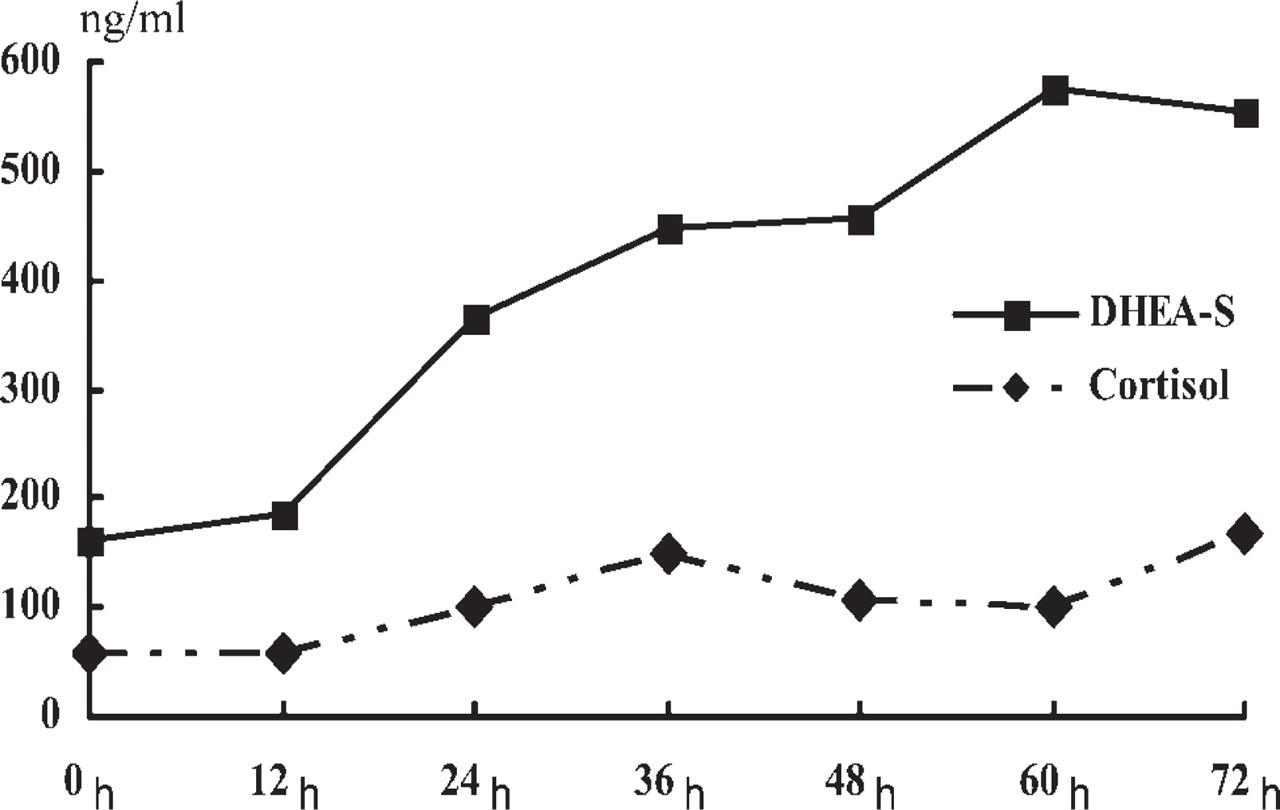
Hormone production in H295R cells stimulated by ACTH. Dehydroepiandrosterone sulfate (DHEA-S) (solid line) and cortisol (dotted line) were measured in a time course after ACTH addition.
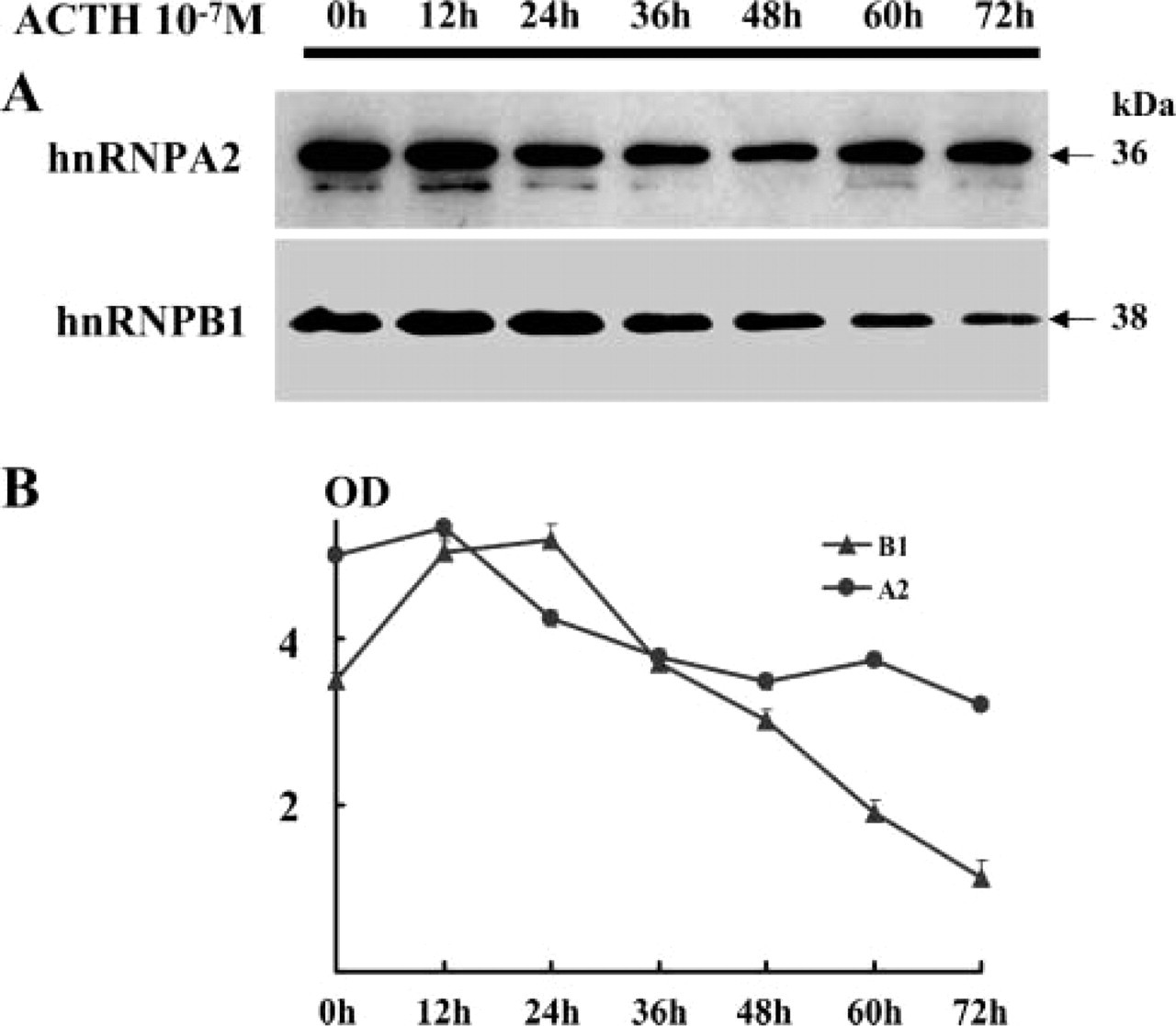
Expression of B1 and A2 proteins in H295R cells stimulated after ACTH in a time course.
In conclusion, we have shown that B1 protein expression is variable in the adrenal cortex, especially in the zona fasciculata-reticularis, and correlates with the producing activity of cortical hormones responding to ACTH stimulation. An additional goal will be to elucidate the alternative splicing mechanism in the adrenal cortex. Further in vivo studies are necessary to clarify the relationship between B1 protein expression and the alternative splicing pattern of regulatory genes such as ACTH receptors, CRE-binding transcription factors, and enzymes involved in steroidogenesis.