Free accessResearch articleFirst published online 2004-9
Occurrence of l - and d -Crustacean Hyperglycemic Hormone Isoforms in the Eyestalk X-Organ/Sinus Gland Complex During the Ontogeny of the Crayfish Astacus leptodactylus
We studied the ontogeny of the eyestalk structure and of the l-CHH and d-Phe3-CHH synthesis in the X-organ/sinus gland (XO/SG) complex by light microscopy and immunocytochemistry in the freshwater crustacean Astacus leptodactylus. The optic ganglia start to differentiate in embryos at EI 190 μm (EI: eye index; close to 410 μm at hatching). At EI 270 μm, the three medullae (externa, interna, and terminalis) and the lamina ganglionaris are present and are organized as in the adult eyestalk. The l-CHH was localized in perikarya of neuroendocrine cells, in their tracts, and in SG from the metanauplius stage to the adult. The d-Phe3-CHH was visualized in XO perikarya, in their tracts and in SG of embryos from EI 350 μm and in all later studied stages. Co-localization of both CHH stereoisomers always occurred in the d-Phe3-CHH-producing cells. These results show that the synthesis of CHH enantiomers starts during the embryonic life in A. leptodactylus, and that the d-isomer is synthesized later than its l-counterpart. We discuss the post-translational isomerization as a way to generate hormonal diversity and the putative relation between d-Phe3-CHH synthesis and the ability to osmoregulate, occurring late during the embryonic life of Astacus leptodactylus.
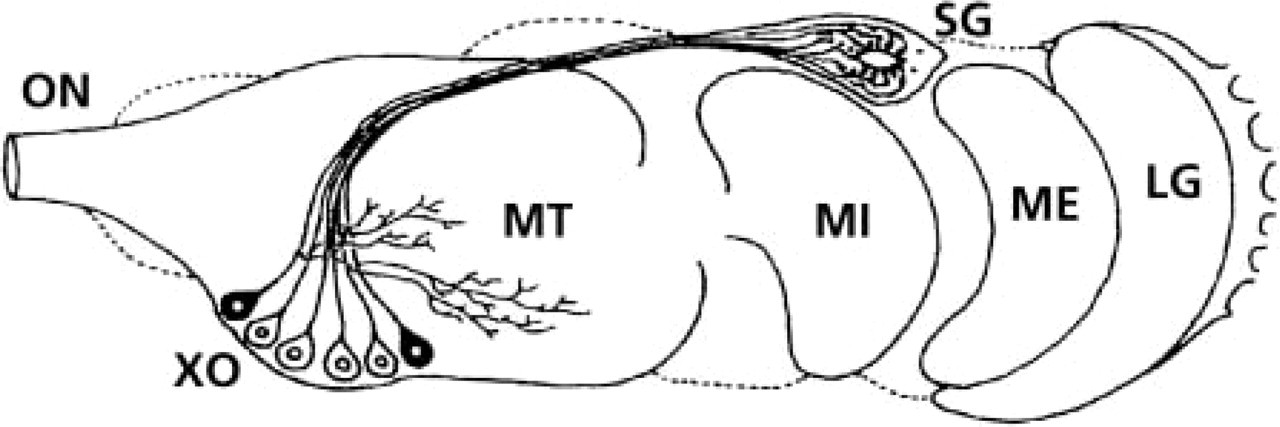
The X-organ/sinusgland (XO/SG) complex located in each eyestalk represents a major neuroendocrine structure in decapod crustaceans (for review see Fingerman 1992). Several physiological processes are controlled by neurohormones from the eyestalks. The neurohormones, synthesized by a proximal cluster of neuroendocrine cells called the X-organ (XO) (Hanström 1928), are conducted distally via axons regrouped in tractus to the storage and release neurohemal organ, the sinus gland (SG) (Figure 1). A wide variety of 8–9.5-kD peptides, isolated from the XO/SG complex and exhibiting a sequence homology and overlapping biological effects (Soyez 1997; Van Herp 1998; Lacombe et al. 1999), have been grouped in the so-called crustacean hyperglycemic hormone family: the molt-inhibiting hormone (MIH), the vitellogenesis-inhibiting hormone (VIH), the mandibular organ-inhibiting hormone (MOIH), and the crustacean hyperglycemic hormone (CHH) (for review see Keller 1992).
The biological and physiological properties of CHH, the most abundant hormone from the SG, have been intensively studied. Initially known as a hyperglycemic factor (Abramowitz et al. 1944), CHH is also involved in lipid metabolism, release of enzymes from the digestive gland, inhibition of methyl farnesoate synthesis, inhibition of ecdysteroid synthesis, stimulation of oocyte development (for review see Böcking et al. 2002), and osmoregulation (Charmantier-Daures et al. 1994; Chung et al. 1999; Spanings-Pierrot et al. 2000; Serrano et al. 2003). In astacid crustaceans, different molecular forms of CHH, including stereoisomers, have been characterized from the XO/SG complex (Soyez et al. 1990,1998). CHH polymorphism resulting from the post-translational isomerization of the phenylalanine residue in position 3 of the N-terminal fragment from the l- to the d-configuration has been reported in the adult lobster Homarus americanus (Soyez et al. 1994) and in adult crayfish species (Yasuda et al. 1994; Aguilar et al. 1995; Soyez et al. 1998; Bulau et al. 2003; Serrano et al. 2003). This kind of modification often generates peptides that have different specialized functions or with differential effects (Soyez et al. 2000).
Diagrammatic representation of the ganglia and the X-organ/sinus gland complex in eyestalks of decapod crustaceans (dorsal view). LG, lamina ganglionaris; ME, medulla externa; MI, medulla interna; MT, medulla terminalis; ON, optic nerve; SG, sinus gland; XO, X-organ perikarya (after Böcking et al. 2002).
The histogenesis of the eyestalk neuroendocrine centers, with special emphasis on the XO-SG axons, has been most often studied during the period of post-embryonic development. Although the XO cell bodies were usually identified at the first larval stages, the SG was detected from the early post-hatch crustaceans (mainly in freshwater species) to the fifth larval stage (Rotllant et al. 1994,1995). Through immunocytochemical (IHC) and in situ hybridization (ISH) techniques, well-defined neurohormone-producing cells have been detected during the postembryonic period: the CHH-producing system in juvenile Astacus leptodactylus (Gorgels-Kallen and Meij 1985), cells producing proctolin-like substances in the larvae of H. americanus (Beltz and Krawitz 1987; Beltz et al. 1990), the MIH-producing system in the first zoeal stage of Carcinus maenas (Webster and Dircksen 1991), CHH and GIH neuroendocrine cells in H. gammarus larvae and postlarvae (Rotllant et al. 1993; Castany 1996), and CHH-containing cells in two postembryonic stages of Procambarus clarkii (Escamilla-Chimal et al. 2001).
However, studies dealing with the organogenesis of the eyestalk and the development of neurohormone-producing cells in the XO/SG complex during the embryonic phase of crustaceans are very scarce. In the crayfish, which undergoes direct development (post-embryonic larval stages are suppressed), the genesis of one of the main neuroendocrine centers (XO/SG) appears important to investigate from the embryonic phase, while essential physiological processes develop. We therefore used light microscopy and immunocytochemical techniques to follow the histogenesis of the eyestalk and the localization and the appearance of two CHH enantiomers, l-CHH and d-Phe3-CHH, during the embryonic and postembryonic development of the crayfish A. leptodactylus.
Materials and Methods
Animals
Berried females of A. leptodactylus, as identified by Eschscholtz in 1823, imported from Russia and Turkey were obtained from a commercial retailer (“Petit Verdus;” Saint Guilhem-Le-Désert, HÉrault, France). In the laboratory, they were first kept in 3 m3 freshwater (FW: dechlorinated tap-water) tanks, then transferred to 40 l FW plastic tanks when the eggs hatched. FW was aereated and filtered (Eheim system). Each tank was covered with an opaque black plastic sheet to reduce visual disturbance of the crayfish. Temperature was kept at 19 ± 1C and photoperiod was held constant at 12-hr light/12-hr dark. Adult crayfish were fed three times per week with fragments of mussels, and juveniles daily with frozen Artemia sp. Samples of 5–10 eggs per female were removed from the egg mass attached on the pleopods of the female two to three times per week to monitor the rate of development of the embryos. During the crayfish embryonic development, the nauplius molts into a metanauplius and the eye pigmented area appears and grows throughout the metanauplius period (Zehnder 1934). After the eyes’ appearance, the eye index (EI) of embryos was calculated according to the method developed by Perkins (1972) in the American lobster. The maximal width and length of the pigmented part of each eye were measured under a stereomicroscope, their mean yielding the EI in μm. The embryonic development duration and the EI at hatch may slightly vary between two seasons of reproduction. In this study, hatching occurred at EI 410 μm. After hatching, the postembryonic development comprises eight juvenile stages separated by molts, including morphological, anatomic, eco-ethological, and physiological changes (Payen 1973; Susanto 2000).
Fixation Procedure
Embryos, juveniles, and eyestalks from adult crayfish were fixed for 48 hr in Bouin's fixative (picric acid-saturated solution 75%; formaldehyde 20%; acetic acid 5%). Eggs membranes were opened with fine forceps to facilitate embryo fixation and the cephalothoraxes of juveniles were isolated under a dissecting microscope 8 hr after the beginning of the fixation. Samples were rinsed in 70% ethanol and fully dehydrated in a graded series of ethanol before embedding in Paraplast X-tra (Sigma-Aldrich; Saint-Quentin-Fallavier, France).
Light Microscopy
Serial transverse sections of embryos and juveniles, and longitudinal sections of eyestalks from adult, were cut at 7 μm on a Leitz Wetzlar microtome and collected on albuminated glycerine-coated slides. Sections were then deparaffinized in two baths of Histochoice Clearing Agent (Amresco; Solon, OH), rehydrated, and stained with Masson trichrome (Martoja and Martoja-Pierson 1967).
Anti-CHH Antisera
The primary antisera were two hapten-specific antisera discriminating between the N-terminal of CHH stereoisomers (anti-octapeptide antisera). These antisera were raised against two synthetic octapeptides with a sequence identical to the N-terminal part of the CHH isoforms of the lobster H. americanus: pGlu-Val-Phe-Asp-Gln-Ala-Cys-Lys for anti-octapeptide-l antiserum (anti-l-CHH) and pGlu-Val-d-Phe-Asp-Gln-Ala-Cys-Lys for anti-d-octapeptide antiserum (anti-d-Phe3-CHH). The production and characterization of the two antisera have been described by Soyez et al. (1998,2000).
Immunocytochemistry
Serial frontal sections of embryos and juveniles, and longitudinal sections of eyestalks from adults, were cut at 4 μm on a Leitz Wetzlar microtome and collected on poly-l-lysine-coated slides. Serial sections were then deparaffinized in two baths of Histochoice Clearing Agent (Amresco), rehydrated, and preincubated for 10 min in 0.01% Tween 20, 150 mM NaCl in 10 mM PBS, pH 7.3. They were then treated for 5 min with 50 mM NH4Cl in PBS and incubated for 10 min in a blocking solution [BS; 1% bovine serum albumin (BSA), 0.1% gelatin in PBS]. To identify the different CHH isoforms, a double immunofluorescence staining was applied to each slide. The sections were first incubated for 2 hr in a moist chamber at room temperature with 100 μl of a mix of both antibodies: guinea-pig anti-octapeptide-l serum diluted 1:200 and rabbit anti-octapeptide-d serum diluted 1:200 in PBS. After rinsing in BS (three times, 5 min), the slides were incubated for 1 hr in a dark moist chamber at RT with secondary antibodies diluted in 1% BSA in PBS: rhodamine TRITC-conjugated goat anti-guinea pig diluted 1:150 (Jackson Immunoresearch Laboratories; West Chester, PA) and Alexa Fluor-conjugated goat anti-rabbit diluted 1:500 (Molecular Probes; Eugene, OR). After extensive washes in BS (four times, 5 min), the sections were mounted in an anti-bleaching mounting medium (80% glycerin, 20% PBS, 2% N-propyl-gallate; Sigma-Aldrich). Control sections were prepared to determine the positively labeled cells and the putative autofluorescence of the tissues: they were treated as described above, except that the primary antisera were omitted.
Sections were examined with a fluorescent microscope (Leitz Diaplan coupled to a Ploemopak 1-Lambda lamp) equipped with two filter sets (570 nm and 488 nm to visualize rhodamine TRITC and Alexa Fluor, respectively) and a phase-contrast device.
Counting and Morphometric Analysis of Labeled Neurons
For counting and morphometric analysis, photographs of every eyestalk section displaying CHH-immunoreactive cell bodies in the XO were taken with a Leica DC 300F numeric camera mounted on the fluorescence microscope at ×400 magnification. The number of labeled cells was counted on photographs of serial sections. On the same photographs, the maximal diameter of each labeled cell was measured using the Leica FW4000I software. The mean ± SD of the diameter and of the number of labeled cells was calculated.
Results
Histogenesis of the Eyestalk
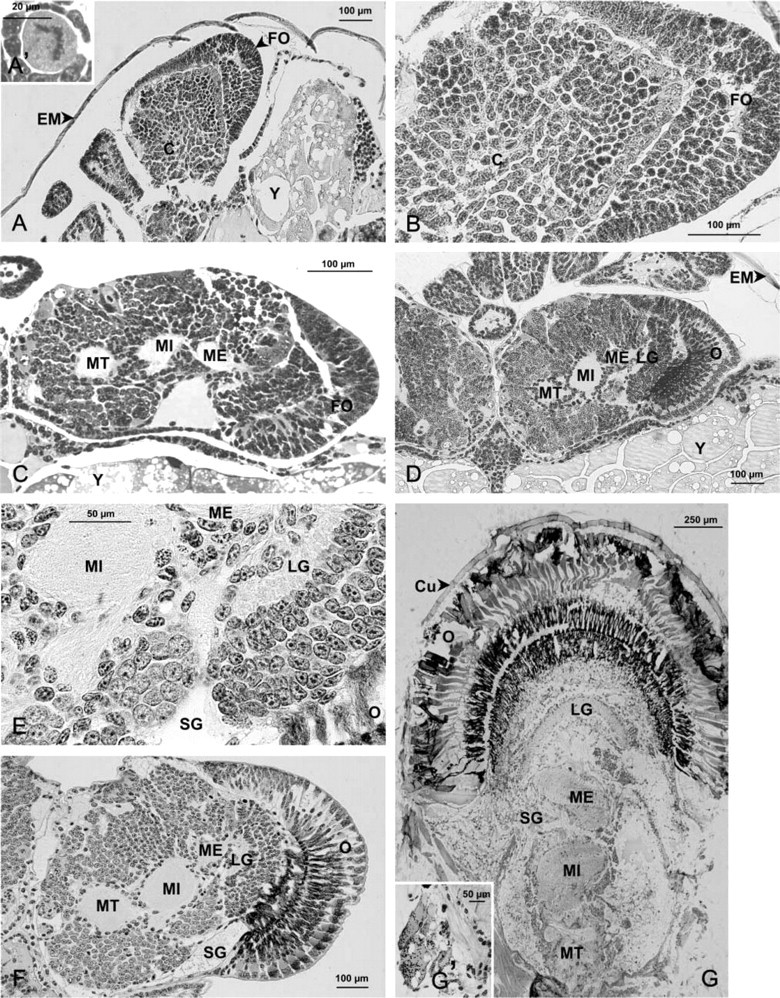
The morphology of the eyestalk in the crayfish A. leptodactylus was examined from nauplius stage to adult by light microscopy (Figure 2). During embryonic development, the prospective eyestalks are sessile and are oriented perpendicular to the longitudinal and rostral axis of the body (Figures 2A, 2C, 2D, and 2F). At the nauplius stage, the eyestalk shape is not distinguishable (not shown). In early metanauplius (before the appearance of eye pigment), the prospective eyes become distinct (Figure 2A) and are composed of two well-separated perikaryon masses (Figures 2A and 2B): a cap of cells at the distal periphery and a central and proximal mass of cells (Figure 2B). Many cells undergoing mitosis were observed (Figure 2A'). At the embryonic stage EI 190 μm, the central neuron mass starts to differentiate into several optic ganglia (Figure 2C). The future ommatidian cells appear still undifferentiated. At EI 270 μm, the medulla terminalis, medulla interna, medulla externa, and lamina ganglionaris are present, superimposed between the optic nerve at the base of the eyestalk and the ommatidia (Figure 2D). The ommatidia are differentiated, longer and dark-colored. All these structures are retained until hatching (Figures 2E and 2F), during the juvenile stages (results not shown), and throughout adult life (Figure 2G).
The SG was first observed in embryos at EI 250 μm (not shown). It is then clearly distinguishable by light microscopy from embryos at EI 320 μm (Figure 2E) to later stages (Figure 2F) and adults (Figure 2G'). A large hemolymph sinus, into which the neurohormones are released, is generally observed on the lateral side close to the yolk.
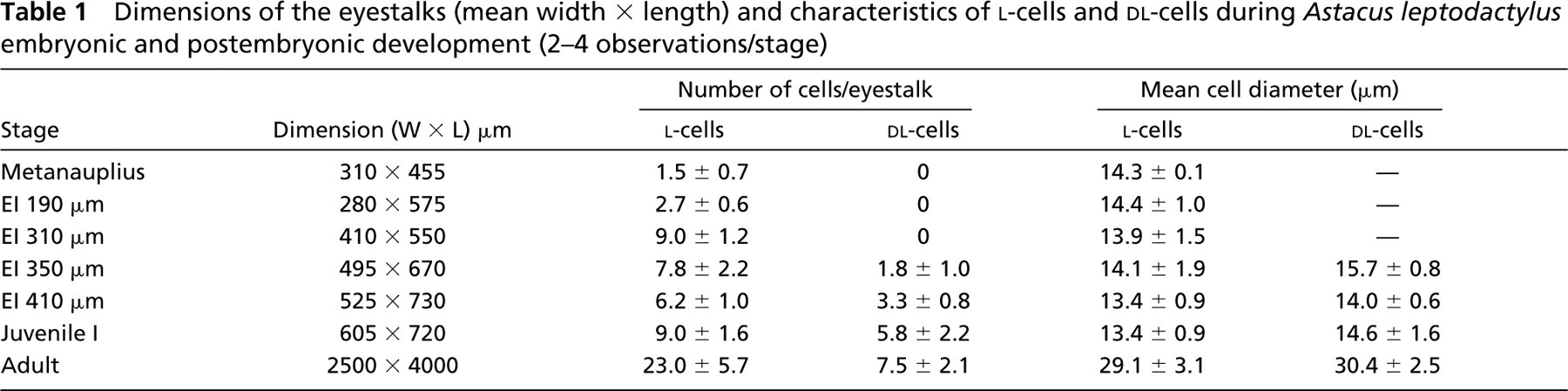
The size of the crayfish eyestalk increases by ≃900% from the youngest metanauplius to the adult stage (Table 1).
Histogenesis of the neuroendocrine system in the eyestalk of Astacus leptodactylus from metanauplius embryonic stage to adult stage. (A–A') Metanauplius (A); detail of a mitotic cell in the eyestalk (A'). (B) metanauplius, eyestalk. (C) Embryo at EI 190 μm, eyestalk. (D) Embryo at EI 270 μm. (E) Embryo at EI 320 μm, detail of the sinus gland region. (F) Embryo at EI 410 μm, eyestalk. (G,G') Adult, eyestalk (G); detail of the sinus gland (G'). C, undifferentiated cells; Cu, cuticle; EM, egg membrane; FO, future ommatidia; LG, lamina ganglionaris; ME, medulla externa; MI, medulla interna; MT, medulla terminalis; O, ommatidia; SG, sinus gland; Y, yolk.
Dimensions of the eyestalks (mean width × length) and characteristics of l-cells and dl-cells during Astacus leptodactylus embryonic and postembryonic development (2–4 observations/stage)
Number of cells/eyestalk
Mean cell diameter (μm)
Stage
Dimension (W×L) μm
l-cells
dl-cells
l-cells
dl-cells
Metanauplius
310 × 455
1.5 ± 0.7
0
14.3 ± 0.1
—
EI 190 μm
280 × 575
2.7 ± 0.6
0
14.4 ± 1.0
—
EI 310 μm
410 × 550
9.0 ± 1.2
0
13.9 ± 1.5
—
EI 350 μm
495 × 670
7.8 ± 2.2
1.8 ± 1.0
14.1 ± 1.9
15.7 ± 0.8
EI 410 μm
525 × 730
6.2 ± 1.0
3.3 ± 0.8
13.4 ± 0.9
14.0 ± 0.6
Juvenile I
605 × 720
9.0 ± 1.6
5.8 ± 2.2
13.4 ± 0.9
14.6 ± 1.6
Adult
2500 × 4000
23.0 ± 5.7
7.5 ± 2.1
29.1 ± 3.1
30.4 ± 2.5
Immunolocalization of l-CHH and d-Phe3-CHH in the X-Organ/Sinus Gland Complex
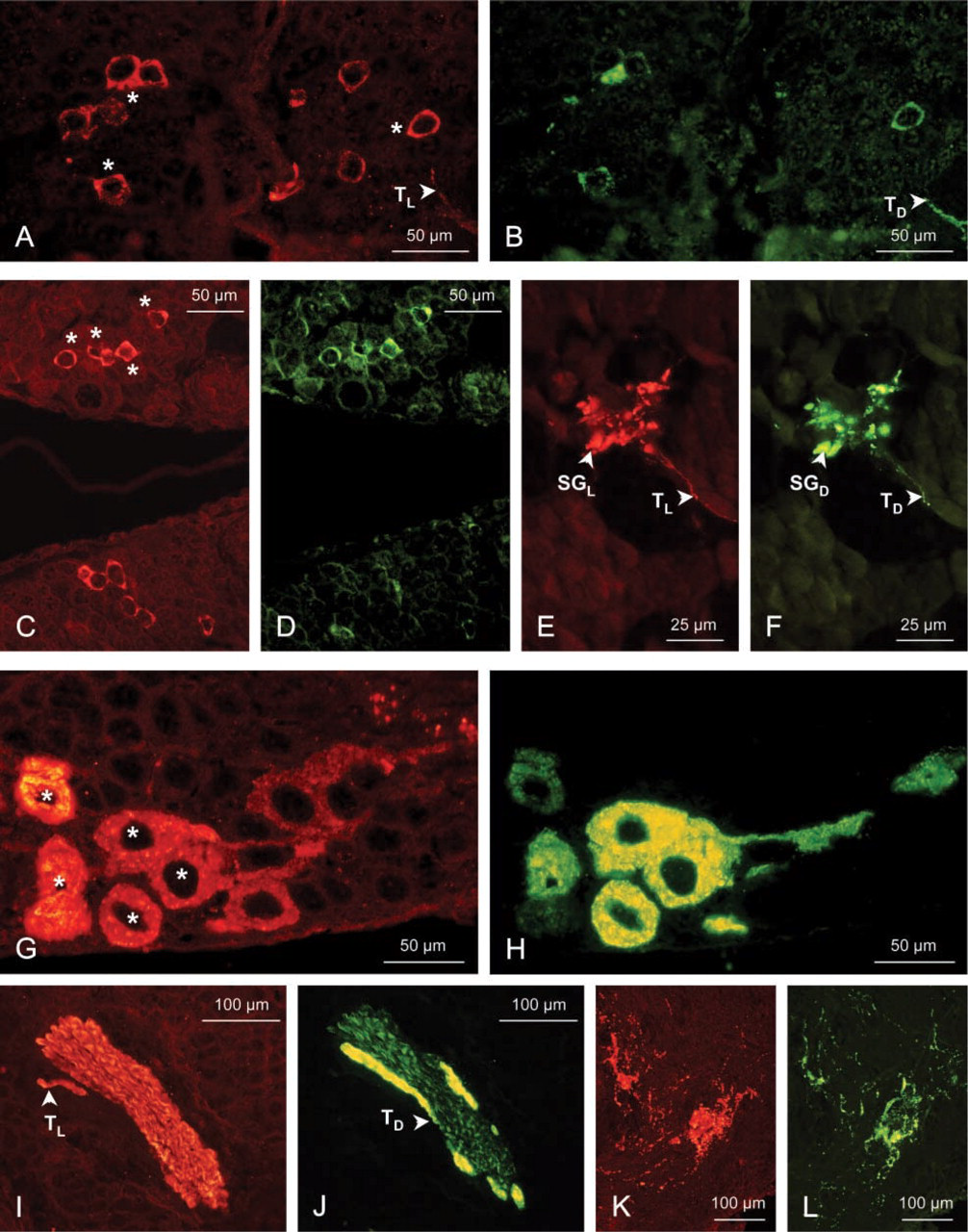
The presence and location of the two CHH stereoisomers, l-CHH and d-Phe3-CHH, were studied in the XO/SG complex of the crayfish A. leptodactylus. Double immunofluorescence labeling using anti-l-CHH and anti-d-Phe3-CHH antisera was performed in embryos starting from the metanauplius stage, in juveniles, and in adults. The results are shown in Figures 3 and 4.
Bouin fixation, paraffin-embedding procedures, and immunocytochemistry yielded good histocytological preservation. Immunoreactivity was not detected in other regions of the eyestalk. The immunoreaction remained negative for all the controls (results not shown).
Localization of Immunoreactivity
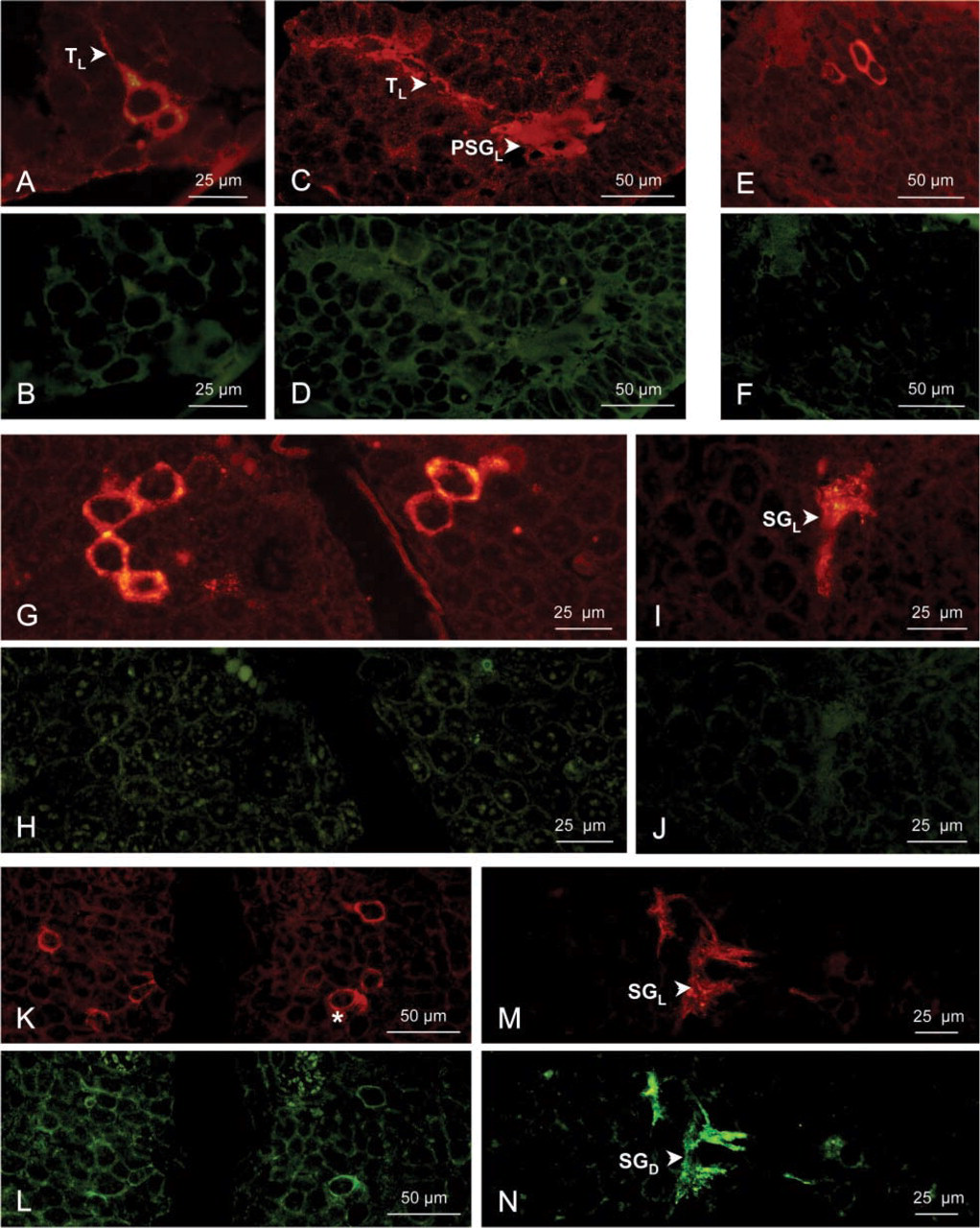
During embryonic development from the metanauplius stage to embryos at EI 300 μm (Figures 3A–3J), immunoreactivity to the anti-d-Phe3-CHH (in green fluorescence) remains negative (Figures 3B, 3D, 3F, 3H, and 3J), whereas the immunoreaction to the anti-l-CHH (in red fluorescence) is positive in a few XO cell bodies (Figures 3A, 3E, and 3G). The anti-l-CHH immunostaining of the tracts and of the presumptive SG is dim in the metanauplius (Figure 3C) but is brighter at EI 190–300 μm (Figure 3I).
In embryos at EI 350 μm and 410 μm, in juveniles, and in adults, the immunoreactivity is positive for the two anti-CHH antisera in several perikarya of the XO neuroendocrine cells. These cells are located at the proximal side of each eyestalk in embryos (Figures 3K, 3L, 4A, and 4B) and in juveniles (Figures 4C and 4D), or they are typically clustered near the medulla terminalis in adults (Figures 4G and 4H). Immunostaining for the d-Phe3-CHH in the XO cells is notably weaker at EI 350 μm (Figure 3L) than at later stages. Two different stainings in the XO cell bodies have been observed from embryos at EI 350 μm to adults: the l-cells, which display a red fluorescence, i.e., staining by the anti-l-CHH alone, and the dl-cells, red and green fluorescence, attesting to labeling by both anti-CHH antisera.
The axon tracts between the neuroendocrine cells and the SG are strongly double-stained (Figures 4A, 4B, 4E, 4F, 4I, and 4J). In the SG, the immunostaining is positive and strong with both antisera, showing red and green aggregations (Figures 3M, 3N, 4E, and 4F). A double fluorescence is also observed close to the XO cell bodies in juveniles (not shown) and in adults (Figures 4K and 4L), whereas it has not been observed during embryonic development. This immunoreaction corresponds to the dendritic branches from the cluster of XO perikarya. When the immunoreactivity was positive in the SG, it was generally located on the external side (close to the yolk) between the medulla externa and medulla interna.
Counting and Morphometry of l- and dl-cells
The results of the morphometric study are summarized in Table 1. From the metanauplius stage to the embryos at EI 310 μm, the number of l-cells per eyestalk increases and then slightly decreases at EI 350 μm while the dl-cells appear. After hatching, the number of l- and dl-cells continues to increase, reaching 23.0 and 7.5, respectively, in adults. In embryos and juvenile I eyestalks, the l- and dl-cell perikarya are ovoid with large nuclei enclosed by a thin layer of cytoplasm, and their mean diameters remain constant (≃14 μm). In adults, the l- and dl-cells have a proportionally larger cytoplasm and a larger diameter (≃30 μm) (Figures 4G and 4H). The two cell types are morphologically indistinguishable at all stages.
Immunolocalization of l-CHH (using a 570-nm filter; red fluorescence, RF) and of d-Phe3-CHH (using a 488-nm filter; green fluorescence, GF) in the X-organ/sinus gland complex of Astacus leptodactylus embryos: metanauplius, EI 190 μm, 300 μm, and 350 μm. (A–D) Metanauplius, RF (A) and GF (B) in the X-organ; RF (C) and GF (D) in the presumptive sinus gland. (E,F) Embryo at EI 190 μm, RF (E) and GF (F) in the X-organ. (G–J) Embryo at EI 300 μm, RF (G) and GF (H) in the X-organs at the proximal side of the two eyestalks; RF (I) and GF (J) in the sinus gland. (K–N) Embryo at EI 350 μm, RF (K) and GF (L) in the X-organs at the proximal side of the two eyestalks; RF (M) and GF (N) in the sinus gland. PSGL, anti-l-CHH-positive presumptive sinus gland; SGD, anti-d-Phe3-CHH-positive sinus gland; SGL, anti-l-CHH-positive sinus gland; TL, anti-l-CHH-positive tractus. Asterisks indicate co-localization of l-CHH and d-Phe3-CHH in cell bodies.
Discussion
Histogenesis and CHH Isoforms Ontogeny in the Eyestalk of A. leptodactylus
The ontogenesis of the eyestalk structure was studied during the embryonic development of the crayfish A. leptodactylus. In the young metanauplius stage, some embryonic cells get closer to shape the future eyestalk but they rapidly divide in distal and proximal groups. In embryos at EI 190 μm, the proximal neuron mass differentiates into three neuroendocrine medullae: medulla externa, interna, and terminalis. In embryos at EI 270 μm, the lamina ganglionaris and the ommatidia cells are elaborated. The general organization of the eyestalk tissues and the spatial arrangement of the four ganglia typically described in adult A. leptodactylus (Van Herp and Van Buggenum 1979; Gorgels-Kallen and Van Herp 1981; Van Deijnen et al. 1985) are therefore already present in the middle phase of embryonic development.
The existence of CHH enantiomers resulting from the isomerization of one amino acid residue in position 3 of the N-terminal fragment from the l- to the d-configuration has been demonstrated in adults of the lobster H. americanus (Soyez et al. 1994) and of several crayfish species such as Orconectes limosus (Soyez et al. 1998), Procambarus bouvieri (Aguilar et al. 1995), Procambarus clarkii (Yasuda et al. 1994), A. leptodactylus (Serrano et al. 2003), and Cherax destructor (Bulau et al. 2003). In the present work, we have studied the ontogenetic pattern of occurrence of the two CHH stereoisomers in the eyestalk of A. leptodactylus. The l-CHH isoform was observed in the neuroendocrine cells of the XO/SG complex from the metanauplius stage to the adult. Immunostaining, intense in the perikarya but weak in the presumptive SG area at the metanauplius stage, showed that the synthesis and transport of this enantiomer start very early during embryonic development, even when the neurohemal organ is not yet well organized. The synthesis, axon transport, and storage of the d-Phe3-CHH in the neuroendocrine complex were observed from embryos at EI 350 μm only and in later stages, including the adult.
In A. leptodactylus, the ratio of XO/SG complex l-cells to dl-cells increases from 2:0 to 6:3 during embryonic development and increases to ≃9:6 and 23:8 in juvenile I and in the adult, respectively. It is noteworthy that the number of dl-cells is proportionally important just before hatching and in early postembryonic stage. The physiological significance of this increase in d-Phe3-CHH production close to the hatching period is discussed below. The average diameter is 14 μm throughout embryonic and early postembryonic development and it reaches 30 μm in the adult. The CHH-cell diameter is remarkably smaller in the lobster embryo eyestalk (7 μm) compared with the crayfish (Rotllant et al. 1995). The morphometric analysis is in agreement with previous studies on juveniles and adults of A. leptodactylus (Gorgels-Kallen and Meij 1985; Gallois et al. 2003) and is very close to the results reported on adults of the related crayfish species O. limosus (Gorgels-Kallen et al. 1982; Soyez et al. 1998,2000). In H. gammarus, the number of CHH-cells increases progressively from 2 to 28 during embryonic development (Rotllant et al. 1995), slightly declines in larval stage I (Rotllant et al. 1993), and doubles in the adult (40 CHH-cells; Gorgels-Kallen et al. 1982). Several authors have suggested that the increase in number of neurohormone-producing cells (CHH- or MIH-cells) during juvenile, larval, or post-larval development is caused by the differentiation of neuroblasts into neuroendocrine cells or by variations in neuropeptide gene expression in differentiated cells, rather than because of cell divisions (Gorgels-Kallen and Meij 1985; Webster and Dircksen 1991; Rotllant et al. 1993).
Immunolocalization of l-CHH (using a 570-nm filter; red fluorescence, RF) and of d-Phe3-CHH (using a 488-nm filter; green fluorescence, GF) in the X-organ/sinus gland complex of Astacus leptodactylus embryos at EI 410 μm, juveniles and adults. (A,B) Embryo at EI 410 μm, RF (A) and GF (B) in the X-organs at the proximal side of the two eyestalks. (C–F) Juvenile I, RF (C) and GF (D) in the X-organs at the proximal side of the two eyestalks; RF (E) and GF (F) in the sinus gland. (G–L) Adult, RF (G) and GF (H) in the X-organ; RF (I) and GF (J) in the axon tractus; RF (bK) and GF (L) in the dendritic branches of the neuroendocrine cells. SGD, anti-d-Phe3-CHH-positive sinus gland; SGL, anti-l-CHH-positive sinus gland; TD, anti-d-Phe3-CHH-positive tractus; TL, anti-l-CHH positive tractus. Asterisks indicate co-localization of l-CHH and d-Phe3-CHH in cell bodies.
Only a few studies have dealt with the occurrence of CHH or CHH enantiomers in the XO/SG complex of crustacean embryos. Using a polyclonal anti-CHH H. americanus antiserum in the European lobster H. gammarus, synthesis of CHH was detected from embryos at EI 114 μm (the mean EI at hatch for this species is 650 μm) (Rotllant et al. 1995). In the same species, Castany (1996) showed a negative immunoreaction for both anti-l-CHH and anti-d-Phe3-CHH antisera in embryos at EI 100 μm and a positive immunoreaction for both antisera in embryos at EI 427 μm, but no data are available between these two stages. We report here that, in the eyestalk of A. leptodactylus, some neuroendocrine cell bodies have been labeled exclusively by anti-l-CHH antiserum (l-cells) or by both anti-l- and anti-d-Phe3-CHH antisera (dl-cells) starting at EI 350 μm. Such a co-localization has also been observed in adults of the same species by immunogold electron microscopy (Gallois et al. 2003), of the crayfish O. limosus (Soyez et al. 2000), and of the lobsters H. americanus (Soyez et al. 1998) and H. gammarus (Van Herp 1998). In dl-cells of A. leptodactylus, Gallois et al. (2003) have demonstrated the presence of l-, d-, and dl-granules from the cell body to the SG, the percentage of l-granules decreasing and that of d-granules increasing throughout the transport. This observation indicates that the l to d isomerization occurs in the dl-cells’ perikarya but also through the axons and in the SG. The dl-cells most probably derive from l-CHH-producing cells that acquire the ability to isomerize l-CHH during development. Another hypothesis is that non-producing CHH neurons start at a given time to produce CHH very rapidly isomerized in the d-isoform.
The presence of immunoreactivity in dendritic collateral branches close to the XO cell bodies observed in juveniles and adults of A. leptodactylus (this study) has also been reported for CHH in different crustacean groups (Dircksen et al. 1988; Subramoniam et al. 1998; Soyez et al. 2000) and for MIH in different crab species (Dircksen et al. 1988; Watson et al. 2001; Lee and Watson 2002). These dendritic branches are classically considered as important regulatory input areas. In particular, Ollivaux et al. (2002) reported in O. limosus close appositions between terminals of enkephalinergic neurons and dendritic branches of l-and dl-cells, suggesting a control via direct synaptic contacts.
Which Benefits in d-stereoisomerization? Why Does d-Phe3-CHH Occur Late During the Embryonic Development of A. leptodactylus?
CHH from the eyestalks of adult decapod crustaceans is known to be mainly involved in the regulation of carbohydrate metabolism. In this study we observed that the l-CHH is synthesized and transported starting at the embryonic metanauplius stage and that it is stored in a functional SG at EI 190 μm. These results indicate that l-CHH matures and is released rather precociously, probably to regulate a physiological function(s) that might be vital at the onset of embryonic development, such as control of glycemia.
More recently, studies have emphasized that CHH plays a direct or indirect role in crustacean osmoregulation. After the first demonstration of this function in H. americanus (Charmantier-Daures et al. 1994), the implication of a gut CHH-like peptide in osmoregulatory mechanisms at ecdysis has been reported in the crab Carcinus maenas (Chung et al. 1999). Then a purified CHH from the sinus gland of the crab Pachygrapsus marmoratus was shown to increase Na+ influx in perfused gills (Spanings-Pierrot et al. 2000). In addition, the neurohormone ITP isolated from different orthopteran insects, and showing a high structural homology with hormones of the CHH family (Soyez 1997), stimulates ileal reabsorption of ions and fluid (reviewed in Phillips et al. 1998; Macins et al. 1999). Interestingly, two studies investigating differential effects of CHH isoforms on osmoregulatory parameters have revealed that the d-isomer was the most efficient one in the adult of two astacid crustaceans, the lobster H. americanus (Charmantier-Daures et al. 1994) and the crayfish A. leptodactylus (Serrano et al. 2003).
The ontogeny of osmoregulation has been studied in embryos of A. leptodactylus. The ability to hyperosmoregulate and the presence of gill Na+-K+-ATPase, one of the main enzymes involved in ionoregulation, appear late in the course of embryonic development, corresponding to 93–97% of the eye pigment development (Susanto 2000; Susanto and Charmantier 2001). Interestingly, the synthesis, transport, and storage of the d-Phe3-CHH in the XO/SG complex start slightly earlier, i.e., at 85% of eye pigment development (this study). Because CHH is involved in osmoregulating processes in different crustaceans, and particularly because a stimulating effect of the d-Phe3-CHH on hemolymph osmolality and Na+ content has been demonstrated in adult A. leptodactylus, we can hypothesize that the occurrence of the d-Phe3-CHH is positively involved in the control of the ability to osmoregulate in crayfish late embryos. The occurrence of a functional neuroendocrine control (XO/SG and d-Phe3-CHH) would anticipate by a few days the onset of hyper-osmoregulation, occurring itself a few hours before hatching of the young juvenile crayfish in freshwater. During the very late embryogenesis of the shore crab C. maenas, new neurons in the nervous system also begin to express two CHH forms (X-organ variant or pericardial organ variant), and this peak of synthesis appears to be involved in the massive surge in CHH released immediately before larval eclosion (Webster and Chung 2003). It is noteworthy that zoeae I of C. maenas are slightly able to osmoregulate at hatch (Cieluch et al. 2004). Because their ability to osmoregulate increases during their postembryonic ontogeny, it would be worthwhile to study the expression of CHH over this period.
The advantage(s) of elaborating d-peptides is still an open debate. A first hypothesis is that the presence of a d-amino acid residue in the N-terminal part of a peptide may increase its resistance to proteolytic enzymes, thus extending its half-life. For example, the d-Ala2-dermorphin isolated from the skin of a frog is not hydrolyzed by aminopeptidases, whereas the l-peptide is rapidly degraded (Kreil 1997). The venom of the spider Agenelopsis aperta contains several calcium channel toxins, including w-aga-IVB (= d-Ser46-agatoxin) and w-aga-IVC (= l-agatoxin), the d-agatoxin being four times more efficient than the l-form (Heck et al. 1994). The d-Phe3-CHH from the eye-stalks of the crayfish O. limosus and A. leptodactylus shows a longer hyperglycemic effect compared with l-CHH (Keller et al. 1999; Serrano et al. 2003). A second postulate is that stereoisomerization may modify the affinity between the peptide and its receptors, owing to the change in the secondary structure of the peptide. This process would generate peptides with different specialized functions from one gene (Soyez et al. 2000). For example, among several stereoisomers of achatin isolated from the African giant snail Achatina fulica (Kamatani et al. 1989), d-Phe2-achatin (achatin-I) has a neuroexcitatory effect on the giant neurons of the snail, whereas its l-stereoisomer (achatin-II) does not possess biological activity and has not yet a defined role (Fujimoto et al. 1991; Kamatani et al. 1991; Kim et al. 1991). The d-Phe3-CHH inhibits ecdysteroid synthesis like the MIH in Procambarus clarkii, whereas the l-CHH isomer has no effect on this process (Yasuda et al. 1994). In eyestalkless A. leptodactylus, hemolymph osmolality and Na+ content increased after injection of d-Phe3-CHH, but no effect on osmolality and a much lower effect on ionic concentration were noted with l-CHH (Serrano et al. 2003).
This study reports the entire pattern of occurrence of the l-CHH and the d-Phe3-CHH throughout the embryonic development of a freshwater crustacean. In summary, the l-isomer is synthesized early in the young Astacus leptodactylus metanauplius, whereas the d-enantiomer occurs late in the course of embryonic life. Crayfish embryos may start to isomerize CHH at the end of their development to generate a hormonal factor involved in osmoregulation and essential at hatching. Unlike marine or brackish water species larvae, in which the ability to osmoregulate generally occurs during the metamorphic transition (for review see Charmantier 1998), the freshly hatched crayfish juveniles are efficient hyperosmoregulators (Susanto and Charmantier 2001). Because they hatch in freshwater and therefore must respond to the corresponding osmotic stress (massive water influx and ion loss), the osmoregulatory mechanisms and the (neuro) endocrine systems that regulate them are functional when the egg membranes open.
Footnotes
Acknowledgements
We thank Dr D. Soyez for the gift of the primary antisera and for fruitful discussions regarding the manuscript. We also wish to thank Mr F. Aujoulat and Ms L. Kernegger for technical help. L.S. was financially supported by a doctoral fellowship (no. 1491–2001) from the French Ministère de l'Education Nationale, de la Recherche et de la Technologie.
References
1.
AbramowitzAAHisawFLPapandreaDN (1944) The occurrence of a diabetogenic factor in the eyestalks of crustaceans.Biol Bull86:1–5
2.
AguilarMBSoyezDFalchettoRArnottDShabanowitzJHuntDFHubermanA (1995) Amino acid of the minor isoform of the crustacean hyperglycemic hormone (CHH-II) of the Mexican crayfish Procambarus bouvieri (Ortmann): presence of a d-amino acid.Peptides16:1375–1383
3.
BeltzBSKrawitzEA (1987) Physiological identification, morphological analysis and development of identified serotonin-proctolin containing neurones in the lobster ventral cord.Neurosci7:533–546
4.
BeltzBSPontesMHelluySMKrawitzEA (1990) Pattern of appearance of serotonin and proctolin immunoreactivities in the developing nervous system of the American lobster.J Neurobiol21:521–542
5.
BöckingDDircksenHKellerR (2002) The crustacean neuropeptides of the CHH/MIH/GIH family: structures and biological activities. In WieseKThe Crustacean Nervous System.Berlin, Heidelberg, New York, Springer, 84–97
6.
BulauPMeisenIReichwein-RoderburgBPeter-KatalinicJKellerR (2003) Two genetic variants of the crustacean hyperglycemic hormone (CHH) from the Australian crayfish, Cherax destructor: detection of chiral isoforms due to posttranslational modification.Peptides24:1871–1879
7.
CastanyJ (1996) Ontogenèse du complexe neuroendocrine organe X-glande du sinus et production de CHH (crustacean hyperglycemic hormone) chez Homarus gammarus (L., 1758). PhD thesis, Université Montpellier II, Montpellier
8.
CharmantierG (1998) Ontogeny of osmoregulation in crustaceans: a review.Invert Reprod Dev33:177–190
9.
Charmantier-DauresMCharmantierGJanssenKPCAikenDEVan HerpF (1994) Involvement of eyestalk factors in the neuroendocrine control of osmoregulation in adult lobster Homarus americanus.Gen Comp Endocrinol94:281–293
10.
ChungJSDircksenHWebsterSG (1999) A remarkable, precisely timed release of hyperglycemic hormone from endocrine cells in the gut is associated with ecdysis in the crab Carcinus maenas.Proc Natl Acad Sci USA96:13103–13107
11.
CieluchUAngerKAujoulatFBuchholzFCharmantier-DauresMCharmantierG (2004) Ontogeny of osmoregulatory structures and functions in the green crab Carcinus maenas (Crustacea, Decapoda).J Exp Biol207:325–336
12.
CookeIMSullivanRE (1982) Hormones and neurosecretion. Neurobiology: structure and function. In AtwoodHLSandemanDCThe Biology of Crustacea.New York, Academic Press, 250–290
13.
DircksenHWebsterSGKellerR (1988) Immunocytochemical demonstration of the neurosecretory systems containing putative moult-inhibiting hormone and hyperglycemic hormone in the eyestalk of brachyuran crustaceans.Cell Tissue Res251:3–12
14.
Escamilla-ChimalEGVan HerpFFanjul-MolesM-L (2001) Daily variations in crustacean hyperglycaemic hormone and serotonin immunoreactivity during development of crayfish.J Exp Biol204:1073–1081
15.
FingermanM (1992) Glands and secretions. In HarrisonFWHumesAGMicroscopic Anatomy of Invertebrates.New York, Willey-Liss, 345–394
16.
FujimotoKKubotaIYasuda-KamataniYMinakataHNomotoKYoshidaMHaradaA. (1991) Purification of achatin-I form from the atria of African giant snail, Achatina fulica, and its possible function.Biochem Biophys Res Commun177:847–853
17.
GalloisDBrisorgueilM-JConrathMMaillyPSoyezD (2003) Posttranslational isomerization of a neuropeptide in crustacean neurosecretory cells studied by ultrastructural immunocytochemistry.Eur J Cell Biol82:431–440
18.
Gorgels-KallenJMeijJTA (1985) Immunocytochemical study of the eyestalk of the crayfish Astacus leptodactylus during larval and postlarval development.J Morphol185:155–163
19.
Gorgels-KallenJVan HerpF (1981) Localization of crustacean hyperglycemic hormone (CHH) in the X-organ sinus gland complex in the eyestalk of the crayfish, Astacus leptodactylus (Nordmann, 1842).J Morphol170:347–355
20.
Gorgels-KallenJVan HerpFLeuvenRSEW (1982) A comparative immunocytochemical investigation of the crustacean hyperglycemic hormone (CHH) in the eyestalks of some decapod crustacea.J Morphol174:161–168
21.
HanströmB (1928) Vergleichende Anatomie des Nervensystems der wirbellosen Tiere.Berlin, Springer
22.
HeckSDSiokCJKrapchoKJKelbaughPRThadeioPFWelshMJWilliamsRD. (1994) Functional consequences of post-translational isomerization of Ser46 in a calcium channel toxin.Science266:1065–1068
23.
JarosPPKellerR (1979) Immunocytochemical identification of hyperglycemic hormone-producing cells in the eyestalk of Carcinus maenas.Cell Tissue Res204:379–385
24.
KamataniYMinakataHKennyPIwashitaTWatanabeKFunaseKSunXP. (1989) Achatin-I, an endogenous neuroexcitatory tetrapeptide from Achatina fulica Férussac containing a D-amino acid residue.Biochem Biophys Res Commun160:1015–1020
25.
KamataniYMinakataHNomotoKKimKYongsiriATakeuchiH (1991) Isolation of achatin-I, a neuroactive tetrapeptide having a D-phenyalanine residue, from Achatina ganglia, and its effects on Achatina giant neurones.Comp Biochem Physiol98C:97–103
26.
KellerR (1992) Crustacean neuropeptides: structures, functions and comparative aspects.Experientia48:439–448
27.
KellerRKegelGReichweinBSedlmeierDSoyezD (1999) Biological effects of neurohormones of CHH/MIH/GIH peptide family in crustaceans. In RoubosEWWendelaar BongaSEVaudryHDe LoofARecent Developments in Comparative Endocrinology and Neurobiology.Maastricht, Shaker, 209–212
28.
KimKTakeuchiHKamataniYMinakataHNomotoK (1991) Slow inward current induced by achatin-I, an endogenous peptide with D-Phe residue.Eur J Pharmacol194:99–106
29.
KreilG (1997) D-amino acids in animal peptides.Annu Rev Biochem66:337–345
30.
LacombeCGrèvePMartinG (1999) Overview on the sub-grouping of the crustacean hyperglycemic hormone family.Neuropeptides33:71–80
31.
LaverdureAMCarette-DesmoucellesCBreuzetMDescampsM (1994) Neuropeptides and related nucleic acid sequences detected in peneid shrimps by immunohistochemistry and molecular hybridations.Neuroscience60:569–579
MacinsAMeredithJZhaoYBrockHWPhillipsJE (1999) Occurrence of ion transport peptide (ITP) and ion transport-like peptide (ITP-L) in Orthopteroids.Arch Insect Biochem Physiol40:107–118
34.
MarcoHGGädeG (1999) A comparative immunocytochemical study of the hyperglycaemic, moult-inhibiting and vitellogenesis-inhibiting neurohormone family in three species of decapod Crustacea.Cell Tissue Res295:171–182
35.
MartojaRMartoja-PiersonM (1967) Initiation aux techniques de l'histologie animale.Paris, Masson et Cie
36.
MontecucchiPCDe CastiglioneRErspamerVR (1981) Identification of dermorphin and Hyp dermorphin in skin extracts of the Brazilian frog Phyllomedusa sauvagei.Int J Peptide Prot Res17:316–321
37.
OllivauxCDircksenHToullecJ-YSoyezD (2002) Enkephalinergic control of the secretory activity of neurons producing stereoisomers of crustacean hyperglycemic hormone in the eyestalk of the crayfish Orconectes limosus.J Comp Neurol444:1–9
38.
OllivauxCSoyezD (2000) Dynamics of biosyntheses and release of crustacean hyperglycemic hormone isoforms in the X-organsinus gland complex of the crayfish Orconectes limosus.Eur J Biochem267:5106–5114
39.
PayenG (1973) Etude descriptive des principales étapes de la morphogenèse sexuelle chez un crustacé décapode à développement condensé, l'écrevisse Pontastacus leptodactylus leptodactylus (Eschscholtz, 1823).Abstr Ann Embryol Morphog6:179–206
40.
PerkinsHC (1972) Developmental rates at various temperatures of embryos of the northern lobster (Homarus americanus Milne-Edwards).Fish Bull70:95–99
41.
PhillipsJEMeredithJAudsleyNRichardsonNMacinsARingM (1998) Locust ion transport peptide (ITP): a putative hormone controlling water and ion balance in terrestrial insects.Am Zool38:461–470
42.
RotllantGCharmantier-DauresMDe KleinDCharmantierGVan HerpF (1995) Ontogeny of neuroendocrine centers in the eyestalk of Homarus gammarus embryos: an anatomical and hormonal approach.Invert Reprod Dev27:233–245
43.
RotllantGCharmantier-DauresMTrillesJ-PCharmantierG (1994) Ontogeny of the sinus gland and the organ of Bellonci in larvae and postlarvae of the European lobster Homarus gammarus.Invert Reprod Dev26:13–22
44.
RotllantGDe KleinDCharmantier-DauresMCharmantierGVan HerpF (1993) Localization of crustacean hyperglycemic hormone (CHH) and gonad-inhibiting hormone (GIH) in the eyestalk of Homarus gammarus larvae by immunocytochemistry and in situ hybridization.Cell Tissue Res271:507–512
45.
SerranoLBlanvillainGSoyezDCharmantierGGroussetEAujoulatFSpanings-PierrotC (2003) Putative involvement of crustacean hyperglycemic hormone isoforms in the neuroendocrine mediation of osmoregulation in the crayfish Astacus leptodactylus.Exp Biol206:979–988
46.
ShihT-WSuzukiYNagasawaHAidaK (2003) Immunohistochemical and morphological studies of hyperglycemic hormone- and molt-inhibiting hormone-producing cells in the eyestalk of Kuruma prawn, Penaeus japonicus, in relation to molting.Invert Reprod Dev43:1–8
47.
SithigorngulPPanchanNVilaivanTSithigorngulWPetsomA (1999) Immunochemical analysis and immunocytochemical localization of crustacean hyperglycemic hormone from the eye-stalk of Macrobrachium rosenbergii.Comp Biochem Physiol124B:73–80
48.
SoyezD (1997) Occurrence and diversity of neuropeptides from the crustacean hyperglycemic hormone family in arthropods.Ann NY Acad Sci814:319–323
49.
SoyezDLaverdureAMKallenJVan HerpF (1998) Demonstration of a cell-specific isomerisation of invertebrate neuropeptides.Neuroscience82:935–942
50.
SoyezDNoelPYVan DeijnenJEMartinMMorelAPayenGG (1990) Neuropeptides from the sinus gland of the lobster Homarus americanus: characterization of hyperglycemic peptides.Gen Comp Endocrinol79:261–274
51.
SoyezDToullecJ-YOllivauxCGéraudG (2000) L to D amino acid isomerization in a peptide hormone is a late post-translational event occurring in specialized neurosecretory cells.J Biol Chem275:37870–37875
52.
SoyezDVan HerpFRossierJLe CaerJPTensenCPLafontR (1994) Evidence for a conformational polymorphism of invertebrate neurohormones.J Biol Chem269:18295–18298
53.
Spanings-PierrotCSoyezDVan HerpFGompelMSkaretGGroussetECharmantierG (2000) Involvement of crustacean hyperglycemic hormone in the control of gill ion transport in the crab Pachygrapsus marmoratus.Gen Comp Endocrinol119:340–350
54.
SubramoniamTPReichweinBDircksenHKellerR (1998) On the isolation and characterization of a crustacean hyperglycaemic hormone in the eyestalk of the shrimp Penaeus indicus.Aquaculture162:99–111
55.
SusantoNG (2000) Adaptation des écrevisses à l'eau douce: ontogenèse de l'osmorégulation chez Astacus leptodactylus. PhD thesis, Université Montpellier II, Montpellier
56.
SusantoNGCharmantierG (2001) Crayfish freshwater adaptation starts in eggs: ontogeny of osmoregulation in embryos of Astacus leptodactylus.J Exp Zool289:433–440
57.
Van DeijnenJEVekFVan HerpF (1985) An immunocytochemical study of the optic ganglia of the crayfish Astacus leptodactylus (Nordmann 1842) with antisera against biologically active peptides of vertebrates and invertebrates.Cell Tissue Res240:175–183
58.
Van HerpF (1998) Molecular, cytological and physiological aspects of the crustacean hyperglycemic hormone family. In CoastGMWebsterSGRecent Advances in Arthropod Endocrinology.Cambridge, Cambridge University Press, 53–70
59.
Van HerpFVan BuggenumHJM (1979) Immunocytochemical localization of hyperglycemic hormone (HGH) in the neurosecretory system in the eyestalk of the crayfish Astacus leptodactylus.Experientia35:1527–1528
60.
Van HerpFVan WormhoudtAVan VenrooyWAJBellon-HumbertC (1984) Immunocytochemical study of crustacean hyper-glycemic hormone in the eyestalks of the prawn Palaemon serratus (Pennant) and some other Palaemonidae, in relation to variations in the blood glucose level.J Morphol182:85–94
61.
WatsonRDLeeKJBordersKJDircksenHLillyKY (2001) Molt-inhibiting hormone immunoreactive neurons in the eyestalk neuroendocrine system of the blue crab, Callinectes sapidus.Arthropod Struct Dev30:69–76
62.
WebsterSGChungJS (2003) Ontogeny of peptidergic neurones during embryonic development of crabs.Comp Biochem Physiol134A:s27
63.
WebsterSGDircksenH (1991) Putative molt-inhibiting hormone in larvae of the shore crab Carcinus maenas L.: an immunocytochemical approach.Biol Bull180:65–71
64.
YasudaAYasudaYFujitaTNayaY (1994) Characterization of crustacean hyperglycemic hormone from the crayfish (Procambarus clarkii): multiplicity of molecular forms by stereoinvertion and diverses functions.Gen Comp Endocrinol95:387–398
65.
ZehnderH (1934) Über die Embryonalentwicklung des Fluskrebses.Acta Zool15:261–408