
Abstract
CD44 is a polymorphic transmembrane glycoprotein that binds hyaluronan and growth factors. Multiple isoforms of the protein can be generated by alternative splicing but little is known about the expression and function of these isoforms in normal development and differentiation. We have investigated the expression of CD44 during normal prostate epithelial cell differentiation. A conditionally immortalized prostate epithelial cell line, Pre2.8, was used as a model system. These cells proliferate at 33C but at 39C stop dividing and undergo changes consistent with early stages of cell differentiation. During the differentiation of these cells, the expression of the CD44 isoform v3-v10 was upregulated. Two layers of epithelial cells can clearly be distinguished in the human prostate, a basal layer expressing keratins 5/14 and a luminal layer expressing keratins 8/18. In prostate tissue the v3-v10 isoform was found predominantly in basal cells but also in keratin 14-negative, keratin 19-positive cells intermediate between the two layers. CD44 v3-v10 was also expressed in other keratin 14-negative prostate tissues, the ejaculatory ducts and prostatic urethra. Therefore, CD44 v3-v10 may be important as a cell surface marker for differentiating cells in the prostate epithelium.
CD44
CD44 has been implicated in a number of normal and disease processes, including intercellular adhesion and tumor metastasis. Normal human salivary gland was found to be negative for the v4/5 isoform but positive for variant exons v3 and v6, where they may be involved in the regulation of growth and renewal of the tissue (Fonseca et al. 2000). Keratinocytes express various CD44 isoforms, including forms containing exons v2-v10, v3-v10, v4-v10, v6-v10, and v8-v10 (Brown et al. 1991; Hudson et al. 1995). Previous reports of CD44 expression in non-malignant human prostate have shown that basal and intermediate epithelial cells express CD44 (Heider et al. 1995; Noordzij et al. 1997; Tran et al. 2002) but little is known about changes in expression of variant isoforms during different stages of epithelial differentiation.
Expression of variant isoforms has been implicated in various human malignancies, including colorectal (Heider et al. 1993b), stomach (Heider et al. 1993a), breast (Matsumura and Tarin 1992), squamous cell carcinoma (Hudson et al. 1996), and prostate cancer (Nagabhushan et al. 1996). In the prostate, CD44s and v6 expression is inversely correlated with tumor stage, grade, and ploidy (Nagabhushan et al. 1996). In addition, there have been reports that loss of CD44s and v6 may be useful prognostic markers in prostate cancer patients treated by radical prostatectomy (Noordzij et al. 1997; Kallakury et al. 1998). Further studies have revealed reduced CD44 expression in prostate cancer metastasis as well as in the corresponding primary tumors (Noordzij et al. 1999). More recently, Iczkowski et al. (2003) demonstrated an increase in expression of variants v7-v9 in prostate cancer. In rat, expression of the v4-v7 isoform is involved in pancreatic carcinoma and metastasis (Gunthert et al. 1991).
The aims of this study were to investigate the expression of CD44 and its isoforms during epithelial cell differentiation in the human prostate. We have developed a cell culture model of differentiation using human prostate epithelial cells isolated from a patient with benign prostatic hyperplasia (BPH) (Daly-Burns et al. unpublished data). Primary cells were conditionally immortalized using a temperature-sensitive SV40 large T-antigen to produce the cell line Pre2.8. At the permissive temperature of 33C the cells are able to proliferate, but when switched to 39C proliferation ceases and the cells undergo morphological changes consistent with the early stages of differentiation. At the higher temperature there is an increase in cell size accompanied by decreased keratin (K) 14 and increased expression of K8, K19, and prostate stem cell antigen (PSCA), a marker for prostate epithelial transit-amplifying cells (Tran et al. 2002). The expression of CD44 during differentiation of prostate epithelial cells was investigated in Pre2.8 cells and in prostate tissue and primary prostate epithelial cell cultures.
Materials and Methods
Cell Line Culture
The human prostate epithelial cell line Pre2.8 was maintained in PrEGM growth medium (Cambrex Bio Science; Wokingham, UK) at 33C in 5% CO2. The cells were grown in flasks pretreated with 10 μg/ml collagen (First Link UK; Birmingham, UK). Cells were maintained at 33C or transferred to 39C for 4, 7, 10, or 14 days before harvesting of RNA and protein extraction or fixation for immunocytochemistry.
Primary Culture of Human Prostate Epithelial Cells
Human prostate tissue was obtained from patients undergoing transurethral resection of the prostate for BPH. Cells were isolated as described previously (Hudson et al. 2000). Briefly, minced prostate tissue was digested in 0.5 mg/ml collagenase type 1A (Sigma Aldrich; Gillingham, UK) in RPMI 1640 (Invitrogen; Paisley, UK) at 37C for 18–20 hr. The digested tissue was centrifuged at 170 × g for 20 sec and the pellet of epithelial acini and ducts digested to a single cell suspension in trypsin/versene (Invitrogen). Epithelial cells were resuspended in PrEGM medium and plated into 6-cm tissue culture dishes at 3000 cells/dish in the presence of a feeder layer of irradiated 3t6 cells.
RT-PCR
Total cellular RNA was isolated from cultured cells or from fresh tissue using the RNEasy Minikit (Qiagen; Crawley, UK) and 1 μg was reverse-transcribed and first-strand cDNA synthesized using Superscript II (Invitrogen) according to the manufacturer's instructions. The reverse transcription reactions were then diluted fivefold and 2 μl was used for PCR amplification. The PCR conditions used were as follows: 94C for 5 min followed by thirty cycles of 94C for 30 sec, 60C for 1 min, and 72C for 1 min 30 sec, with a final extension at 72C for 5 min. PCR products were analyzed by electrophoresis on 1.5% agarose gels stained with ethidium bromide (1 μg/ml). CD44 exon-specific primers were as used previously (Hudson et al. 1995).
DNA Sequencing
PCR products were gel-purified in 1% agarose using a QIAquick PCR Purification Kit (Qiagen). Purified PCR products were sequenced using the dRhodamine Cycle Sequencing Kit (Applied Biosystems; Warrington, UK) on an ABI Prism automated sequencer (PerkinElmer Life Sciences; Beaconsfield, UK). CD44-specific oligonucleotides were used for initiation of sequencing of the purified PCR products.
Western Blotting Analysis
Epithelial cultures were rinsed three times with PBS and incubated on ice for 15 min in the presence of extraction buffer consisting of 1% NP40, 0.4% sodium deoxycholate, 66 mM EDTA, 1 mM phenylmethyl sulphonyl fluoride, and 10 mM TRIS (pH 7.4) (Sigma Aldrich). (Isacke et al. 1990). Protein extracts were centrifuged for 15 min at 4C in an Eppendorf microfuge at 13,000 rpm and the pellets discarded. Total protein concentration was determined using the Bradford assay (Bio-Rad Laboratories; Hemel Hempstead, UK). Protein samples (30 μg) were resolved on a 7.5% polyacrylamide gel and transferred to Immobilon-P membrane (Millipore; Watford, UK). Immunodetection of specific proteins was performed as follows. Nonspecific antibody binding was blocked for 30 min in 5% skimmed milk powder in PBS containing 0.1% Tween-20 (PBST; Sigma Aldrich). The membranes were then incubated for 1 hr with E1/2, anti-CD44, in PBST containing 2% skimmed milk powder, followed by peroxidase-conjugated anti-mouse IgG for 45 min. The membranes were washed in PBST between antibody incubations and antibody binding was detected using the ECL chemiluminescence kit (Amersham Biosciences UK; Chalfont St Giles, UK).
Immunohistochemistry
Prostate tissue from patients undergoing transurethral resection of the prostate (TURP) for the treatment of BPH or from six male cadavers (aged 15 to 36 years; mean 25.2 years) were cut at 3 μm and mounted on Vectabond-treated slides (Vector Laboratories; Peterborough, UK). Antigen retrieval was carried out on deparaffinized tissue sections by boiling in antigen retrieval buffer (Vector Laboratories) for 30 min in a microwave oven. Cultured cells were fixed in formaldehyde on ice for 10 min. Tissue sections or cells were blocked in normal horse serum at room temperature for 30 min before incubation overnight at 4C for tissues or 1 hr for cells, with either E1/2 (a gift from C. Isacke; Isacke et al. 1986), VFF-327 anti-CD44v3, VFF-8 anti-CD44v5, VFF-7 anti-CD44v6, VFF-17 anti-CD44v7/8, VFF-14 anti-CD44v10 (all from Serotec; Oxford, UK (Heider et al. 1993b; Koopman et al. 1993), LP2K anti-keratin 19 (a gift from EB Lane; Stasiak et al. 1989), or LLOO2 anti-keratin 14 (a gift from EB Lane; Purkis et al. 1990). The CD44 antibodies were used at a dilution of 1:100 and the keratin antibodies were supernatants diluted 1:10. After thorough washing in PBS, sections were incubated with isotype-specific fluoroscein isothiocyanate- (FITC) or tetramethyl rhodamine-isothiocyanate (TRITC)-conjugated secondary anti-mouse antibodies (Southern Biotech; Cambridge Bioscience, Cambridge, UK) at RT for 1 hr. Nuclei were stained by incubation with a 1-μg/ml solution of Hoechst 33,258 (Sigma Aldrich) for 5 min. Primary antibody was omitted from negative control tissue sections or cells. Stained sections and cells were mounted in Gelvatol (Monsanto; St Louis, MO) and examined under an Hg-arc Zeiss Axiophot fluorescence microscope. The microscope was coupled to a Coolview 12 cooled charge-couple device (CCD) camera (1024 × 1024, 12-bit pixels; Photonic Science, Robertsbridge, UK) controlled by Image Pro-Plus software v3.0 (Media Cybernetics; Rockville, MD).
Results
CD44 Isoforms Expressed by the Prostate Epithelial Cell Line Pre2.8
To determine which CD44 splice variants are expressed during the differentiation of Pre2.8 cells, RT-PCR was carried out on total RNA extracted from cells grown at permissive and non-permissive temperatures. A schematic diagram of the CD44 gene and the positions of the primers used for RT-PCR is shown in Figure 1A. Primers P1 and P2 were used to identify both standard and variant CD44 isoforms present. Multiple bands were produced at 33C and 39C, and Figure 1B shows a time course of 4, 7, 10, and 14 days after transfer to the higher temperature. All lanes show a band corresponding to the 549-bp standard form of the CD44 gene, which lacks all variable exons. Three further major CD44 isoforms are expressed at the two growth temperatures. However, there is an increase in the relative expression level of the largest isoform at the non-permissive temperature.
To identify the CD44 exons expressed in each isoform, exon-specific RT-PCR was also carried out using a primer located on the 5’ side of the constant region (primer P1) together with a 3’ primer for each of the variant exons v1-v10 (Figure 1C). From the number of bands per lane and the stepwise increase in band size, it is possible to determine which CD44 isoforms are expressed. No product was seen with the v1 primer but all other lanes contained one or more PCR products. The PCR product labeled (i) in Figure 1B appears to contain exons v2-v10, band (ii) exons v6-v10, and the band labeled (iii) exons v8-v10. Band (iv) represents the standard isoform of CD44. PCR products (i-iv) from cells grown at 33C or at 39C for 14 days, as well as BPH cell cultures, were sequenced and the composition of bands (ii), (iii), and (iv) confirmed, although the main band (ii) actually contains v3-v10 and not v2. There is therefore an additional minor band containing v2. A schematic of the main variant forms identified is shown in Figure 1A. Because variant exons v3-v5 were observed only in the largest band, v3-v10, we chose to use v5 as a marker and any positive staining for v5 will indicate cells expressing this isoform.
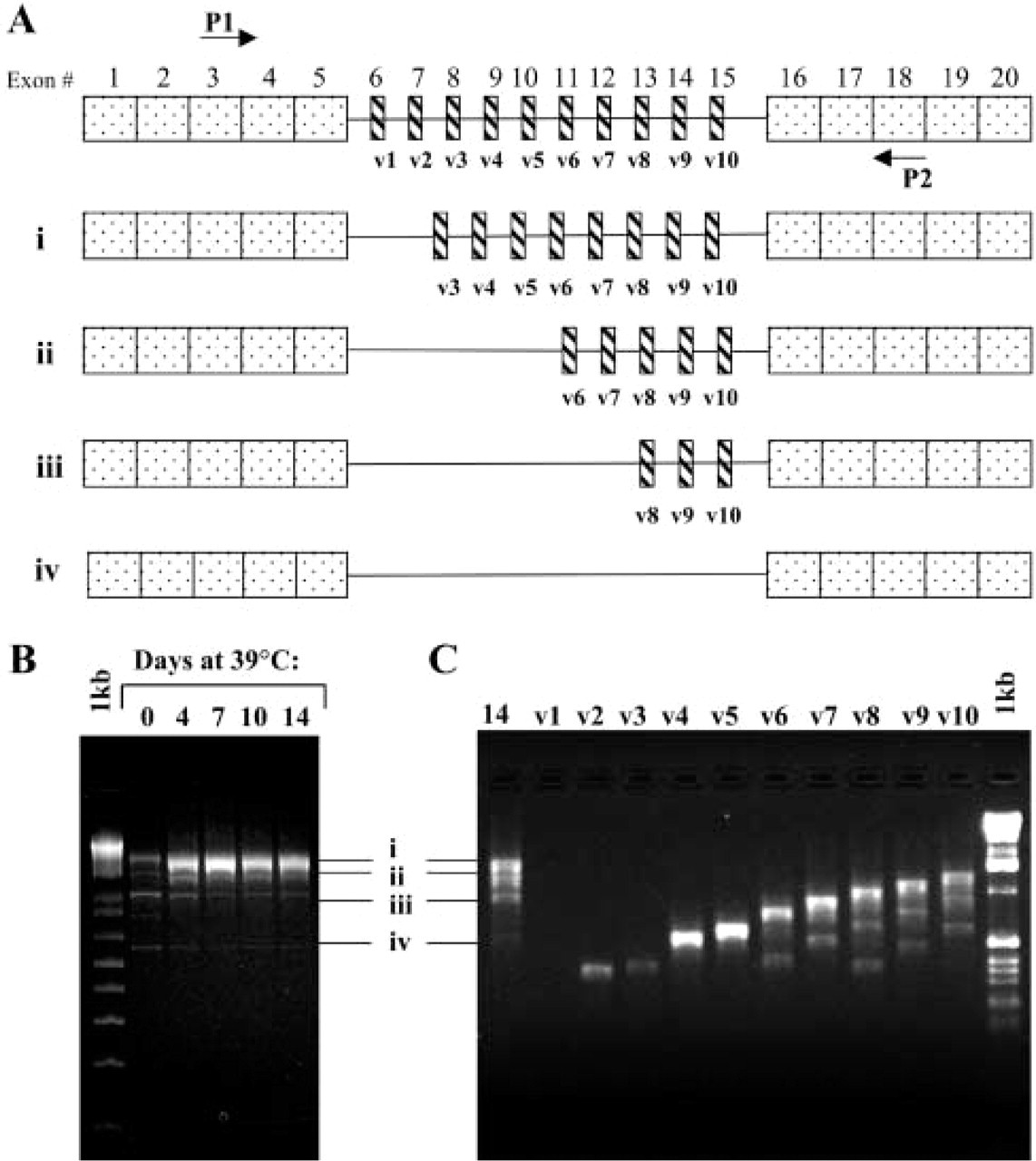
RT-PCR analysis of CD44 in Pre2.8 cells. PCR products were resolved on a 1.5% agarose gel. (
CD44 Protein Expression in Pre2.8 and BPH Cell Cultures
Extracts of Pre2.8 cells and primary cultures of BPH were resolved on SDS-PAGE gels, transferred to Immobilon P membrane, and probed with the antibody E1/2, which recognizes all isoforms of CD44. Equal amounts of protein (30 μg) were loaded in all lanes. Weak expression of the standard 90-kD CD44s was observed in all samples (Figure 2B, arrow), although in some cases it was seen only on longer exposure of the autoradiogram. Three other CD44 isoforms were expressed at 33C by the Pre2.8 cells. The most abundant form has a molecular mass of ≃160 kD, with the least abundant variant being ≃250 kD. As the cells differentiate at 39C, there is a shift in the levels of CD44 protein expression, with the largest isoform being more abundant at this temperature, and by day 14 this is the major CD44 variant expressed (Figure 2A). The CD44 isoforms also have an altered electrophoretic mobility, with a lower apparent molecular weight at 39C than at 33C. A comparison of the bands produced by Pre2.8 with primary epithelial cell culture from BPH tissue revealed a similar pattern of isoforms, with three variant bands and the standard form. The variant bands were expressed at similar levels and they had the same apparent molecular weight as Pre2.8 cells at 33C (Figure 2B).
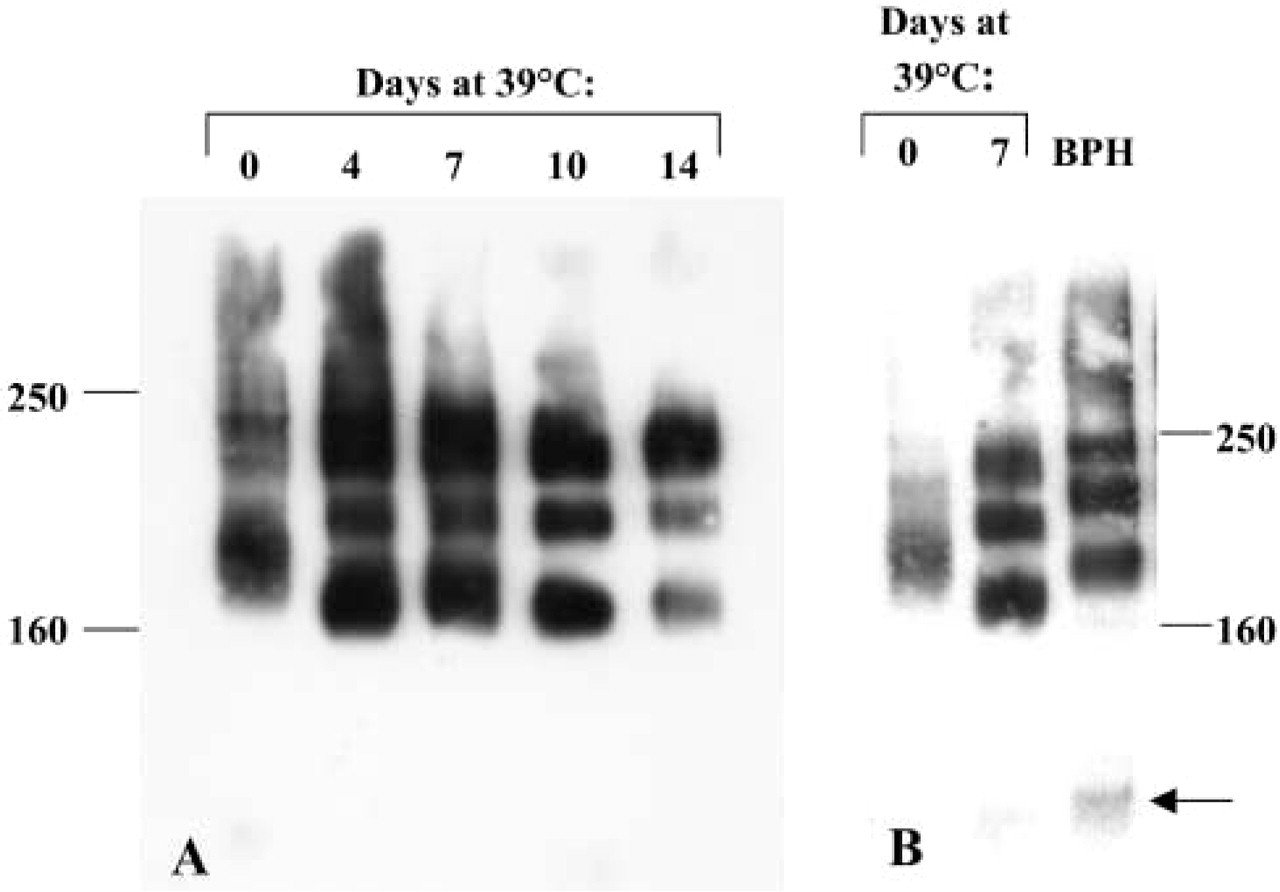
CD44 protein expression in Pre2.8 and BPH cells. (
Cell Surface Expression of CD44 Protein During Differentiation of Pre2.8 Cells and in BPH Cell Cultures
IHC was used to investigate the cell surface expression and distribution of CD44 in Pre2.8 cells. At 33C, when the cells are predominantly proliferative, weak expression of total CD44 protein is observed at cell–cell boundaries (Figure 3A). Only occasional very weak inter-cellular staining for v5-containing isoforms was detected (Figure 3B), although there was strong staining in some cells that appear to be overlying other basal cells. When the cells are switched to the non-permissive temperature of 39C, the increase in cell size is accompanied by an increase in total CD44 protein expression, with strong intercellular staining of almost all cells (Figure 3C). Strong expression of v5 is now detected at cell–cell borders, indicating that at least some of the newly expressed CD44 consists of the v3-v10-containing isoform (Figure 3D). Primary cultures of BPH epithelial cells consist of a mixed population of cells at various stages of the differentiation process. Total CD44 is expressed on the surface of the majority of cells, while the strongest expression of CD44v5 is by a minority of cells that, again, are overlying other cells (Figures 3E and 3F).
CD44 Expression in Prostate Tissue
To confirm that the CD44 isoforms expressed by prostate cells were also expressed in prostate tissue, RNA was isolated from fresh samples of BPH and normal prostate. DNA sequencing of RT-PCR products revealed the same pattern of isoform expression as found in the cell cultures. We therefore examined tissues for the distribution of the major v3-v10 isoform. Because exons v3, v4 and v5 will be found only where the v3-v10 isoform is expressed, an antibody to v5 was used as a marker. Eight BPH samples were stained for K14, K19, and CD44 v5 and the sections were scored for distribution of staining. A typical example of the pattern of double staining for K14 and CD44 v5 is shown in Figure 3. Positive staining for CD44 v5 was observed in all eight samples and in many areas was co-expressed with K14 in basal epithelial cells. CD44 v5 was expressed in basal cells that were negative for K14 in all samples (example shown in Figure 3G). Interestingly, CD44 v5 was also expressed in epithelial cells located in an intermediate position above the basal layer but below the luminal layer (Figure 3G). These cells are taller than the K14 positive basal cells and appear to be leaving the basal layer. This expression pattern is almost identical to that observed previously (Hudson et al. 2001) for K19 in human prostate tissue (Figure 3H).
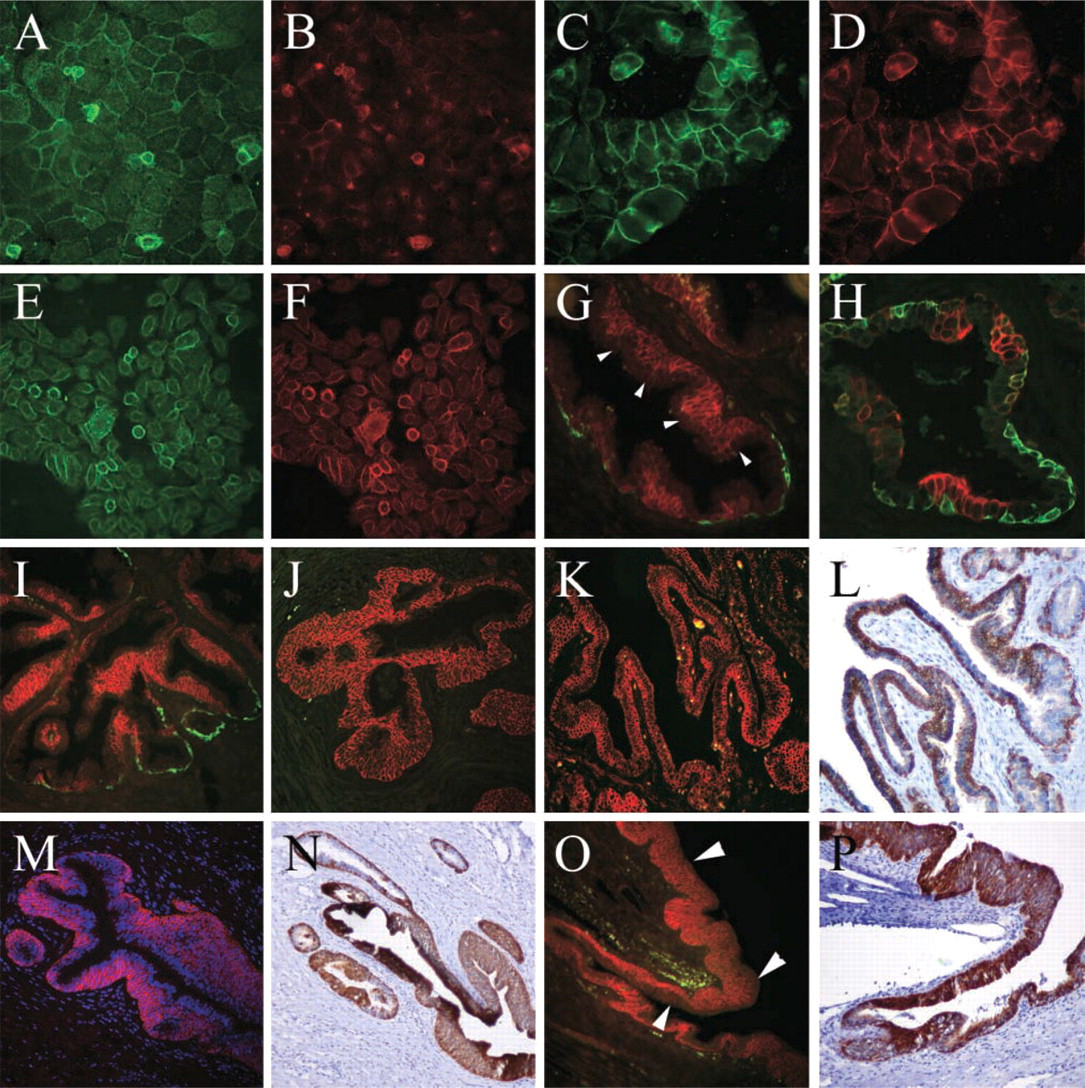
Immunofluorescent staining for cell surface CD44 in prostate cells and tissue. Pre2.8 cells grown at 33C (
To confirm that the staining patterns were not due to aberrant growth of cells in BPH, a further six tissue samples were obtained from tissues collected from young male cadavers. Alternating staining patterns for CD44v5 and K14 were found in all cases and in all three glandular zones, central, peripheral, and transition. In both BPH and normal tissue there were areas of basal cell hyperplasia with multiple K14-negative cell layers expressing high levels of CD44 v5 (Figures 3I and 3J). Some cases showed small areas of CD44v5-negative, K14-positive basal cells (Figure 3I), whereas others did not express K14 (Figure 3J). In all cases a single luminal layer could be distinguished overlying the CD44v5-positive cells.
Other K14-negative prostate-associated epithelial tissues also revealed interesting staining patterns. Ejaculatory ducts within the prostate showed strong basal staining for both CD44v5 (Figure 3K) and K19 (Figure 3L), while prostatic urethra stained strongly in all but the outermost cell layer for CD44v5 (Figure 3M, arrowheads) and in all layers for K19 (Figure 3N). Proximal prostatic ducts, adjacent to the urethra, are colonized by the transitional epithelium, composed of several layers of basal cells and a single layer of larger luminal cells (umbrella cells). In these regions CD44v5 is detected in all cells except the luminal cells (Figure 3O, arrowheads), while K19 is expressed by most cells. In areas where there are only two layers, K19 stains only the basal cells (Figure 3N, upper left). The distribution of the CD44 variant isoforms was further examined by staining the normal tissues with antibodies to CD44v3, v6, v7/8, and v10. Anti-v10 failed to stain at cell–cell boundaries in any of the tissues with high background nuclear staining. The other three antibodies, however, had staining patterns similar to that seen for CD44v5. K14-positive CD44-negative cells were found with all the antibodies, implying that all variant isoforms are preferentially expressed by more differentiated K14-negative basal cells.
Discussion
The prostate is the most common site of benign and malignant neoplastic disease in the human male. Both prostate cancer and BPH are disorders of cell differentiation and proliferation. Epithelial differentiation pathways have been postulated by several investigators (Isaacs and Coffey 1989; Bonkhoff and Remberger 1996; Hudson et al. 2001). However, very little is known about their control in the normal prostate. Therefore, an understanding of these processes will help elucidate the underlying mechanisms of prostate disease.
CD44 has been implicated in the progression of many types of tumor, including prostate cancer. However, little has been documented about the distribution of the CD44 variant isoforms in non-malignant human prostate tissue (Heider et al. 1995; Noordzij et al. 1997). The aim of the present study was to investigate the expression of CD44 during normal human prostate epithelial cell differentiation. RT-PCR analysis and DNA sequencing revealed four CD44 isoforms expressed by prostate tissue, BPH and normal, and by both the immortalized Pre2.8 cells and primary cultures of BPH. The shortest isoform corresponds to the standard form of the gene, whereas the three other isoforms represent alternatively spliced CD44 variants. None of these variants expressed exon v1, which is consistent with the report that this exon contains a stop codon (Screaton et al. 1993). The CD44 transcripts expressed were also observed at the protein level as determined by Western blotting analysis, the largest isoform being expressed most abundantly by day 14 at 39C when Pre2.8 cells are further along the differentiation pathway (Daly-Burns et al. unpublished data). Three major isoforms were expressed at similar levels in primary cultures of BPH epithelial cells. However, these cultures contain a mixed population of proliferating and differentiating cells. Therefore, we used Pre2.8 cells growing at permissive and non-permissive temperatures to provide an in vitro model in which differentiation could be induced.
Although sequencing of RT-PCR products revealed that the CD44 variants expressed by the cells at the two temperatures were all of equal size, an apparent shift in molecular weight was observed by Western blotting analysis. This may be due to differences in the posttranslational modification of the CD44 protein. CD44 contains a number of N- and O-linked glycosylation sites, all of which contribute to further diversity in protein expression. Our results show that the isoforms expressed by Pre2.8 cells grown at the non-permissive temperature have an increased electrophoretic mobility, and therefore a lower molecular mass, compared with those grown at 33C and with the primary BPH cultures. This difference may be the result of reduced glycosylation of the CD44 variants, perhaps caused by aberrant activity of glycosylation enzymes at 39C.
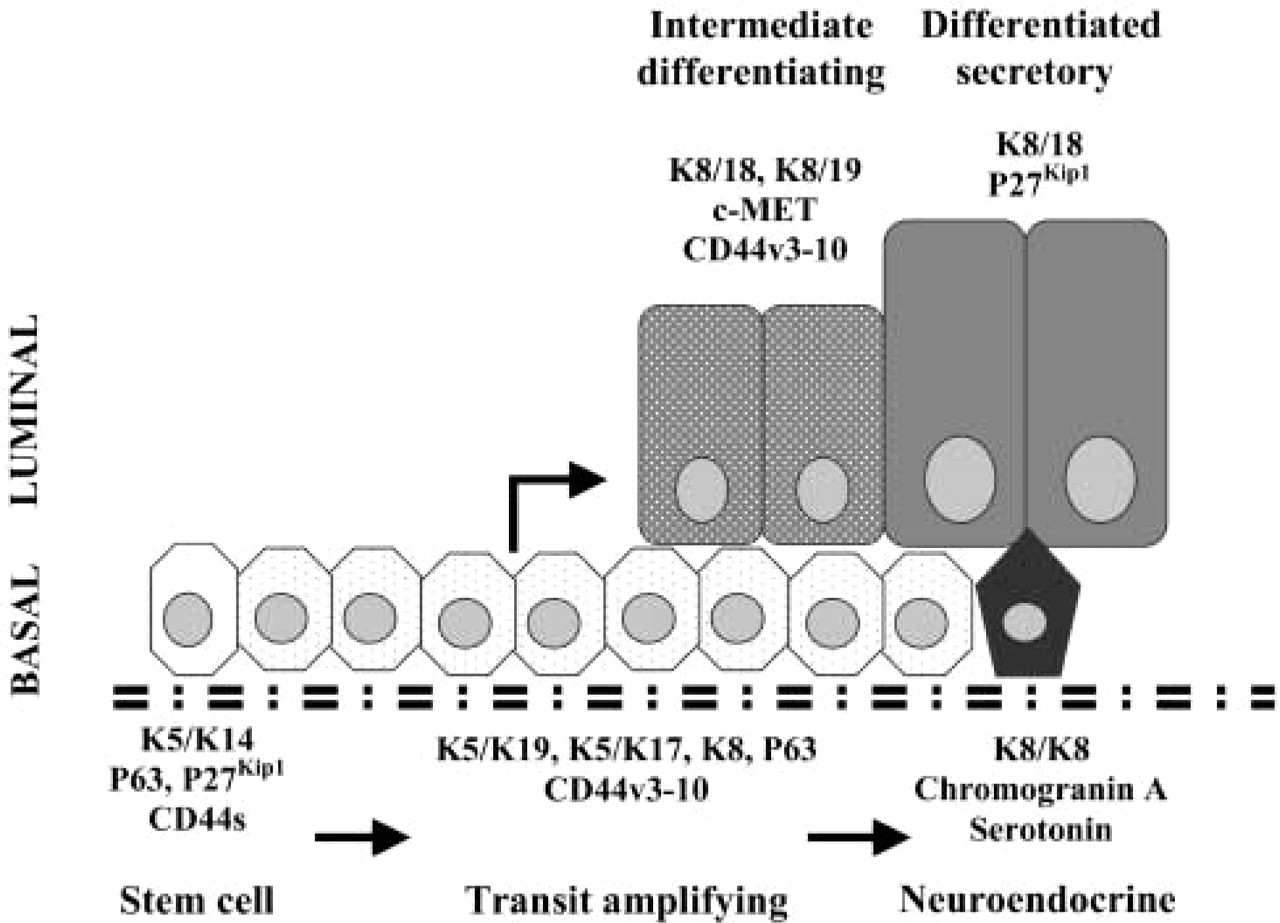
Hypothetical differentiation pathway for human prostatic epithelial cells based on keratin and CD44 expression patterns. Basal stem cells (K5 and K14 only) give rise to an intermediate transit amplifying population that expresses K5 with K19 and K17. These cells differentiate into luminal cells with transient co-expression of K19, K8 and K18 before complete differentiation into secretory cells expressing K8/K18 only. The stem cell population also expresses the short form of CD44 while the transit populations express variant isoforms including CD44v3–10. All basal cells express P63 while stem cells and secreted luminal cells express P27kip1. Intermediate luminal cells transiently express c-MET. In an alternative pathway, transit-amplifying cells differentiate into chromogranin A- and serotonin-expressing neuroendocrine cells. (Redrawn from Hudson et al. 2001, with permission from the Histochemical Society, 2004.)
Our in vitro studies suggested that the CD44 v3-v10 isoform might play a role in the differentiation of normal prostate epithelial cells. On switching to the non-proliferative temperature, upregulation of differentiation-associated markers such as keratin 8, p21, and prostate stem cell antigen (PSCA) occurs (Daly-Burns et al. unpublished data). To confirm whether the v5-containing CD44 isoform may also play a role in vivo, we looked at the expression of the v5-containing variant in prostate tissue specimens. Consistent with reports that CD44 isoforms are expressed by the proliferating compartments of epithelia, positive staining for CD44 v5 was seen in basal epithelial cells in BPH tissues (Mackay et al. 1994). However, many v5-positive cells were negative for K14, a marker for the least-differentiated basal cells. In addition, v5-positive staining was seen in epithelial cells located in a position intermediate between the basal and luminal cell layers. These cells were larger and taller than the characteristically small spindle-shaped basal cells, suggesting that the v5-positive cells may be in the process of differentiating and moving from the basal to the luminal layer. We have previously shown that intermediate cells also stain positively for K19 and have postulated that they may represent an epithelial cell population that is in the process of differentiation (Hudson et al. 2001). Other workers have shown an intermediate population that expresses low levels of K8 together with K5 (van Leenders et al. 2000) and lacks p27kip1, a protein that is expressed by all other epithelial cells in normal prostate (De Marzo et al. 1998). Late intermediate cells have also been shown to express prostate stem cell antigen (Tran et al. 2002), but there have been no cell surface markers for early intermediate cells. Figure 4 shows a hypothetical differentiation pathway for human prostatic epithelial cells that illustrates where the various differentiation markers, including keratins and CD44 isoforms, are expressed (adapted from Hudson et al. 2001).
The expression of CD44 isoforms appears to be cell type-specific. For example, it has been reported that normal pancreatic tissue expresses CD44 variants v3, v6, and v6–10, whereas in pancreatic carcinomas additional splice variants containing exon v5 are detected (Gansauge et al. 1995). This is in contrast to our study in which we have described v5-containing CD44 isoforms to be expressed by normal prostate epithelial cells. Variant isoforms of CD44 are expressed in a variety of other normal epithelial tissues. CD44 v3-v10 is expressed in both proliferating and terminally differentiated cultures of normal keratinocytes (Hudson et al. 1995). This variant plays a role in intercellular keratinocyte adhesion within the epidermis through binding of v5 and v6 to hyaluronan, a major ligand for CD44. It is possible therefore that this isoform plays a similar functional role in prostate epithelium. Recently, it was shown that luminally located intermediate cells in normal prostate also express c-met, the hepatocyte growth factor (HGF) receptor (van Leenders et al. 2002). Interestingly, it had previously been demonstrated that HGF-induced signal transduction is promoted through association of HGF with CD44v3-v10 (van der Voort et al. 1999). This raises the intriguing possibility that the expression of this CD44 contributes to changes in cell motility required by differentiating cells as they move between the basal and luminal cell layers.
These results provide evidence, of early differentiation-associated changes in CD44 expression during normal prostate epithelial cell differentiation. Little is known of the transition between basal and luminal cells in the prostate, and the findings indicate that the CD44 v3-v10- containing isoform may be a marker for the early stages of this differentiation process. CD44 has been used as a target for separating basal from luminal prostate cells using immunomagnetic bead-based cell sorting (Collins et al. 2001). The identification of a CD44 isoform as a cell surface marker specific for the transit-amplifying population in prostate could provide a valuable tool for the further characterization of the differentiation process.
Footnotes
Acknowledgements
Supported by the Wellcome Trust (TA and DH, grant 062869), the US National Institutes of Health (JRM, RO1 AG14960), the European Association of Urologists (IL), and the Bob Champion Cancer Trust (FB and DH).
We thank Clare Isacke and Birgitte Lane for kind gifts of antibodies.