
Abstract
Apolipoprotein D (Apo D) is a secreted lipocalin in the nervous system that may be related to processes of reinnervation and regeneration. Under normal conditions, Apo D is present in the central nervous system in oligodendrocytes, astrocytes, and some scattered neurons. To elucidate the regional and cellular distribution of Apo D in normal human brain, we performed double immunohistochemistry for glial fibrillary acidic protein (GFAP) and Apo D in samples of postmortem human cerebral and cerebellar cortices. Most of the GFAP-positive cells in the gray matter had features of protoplasmic astrocytes and were mainly Apo D-positive. Apo D staining was mostly confined to the cell soma and proximal processes, whereas GFAP extended to a rich and extensive array of processes. The fibrous astrocytes in the white matter were immunoreactive for GFAP but not for Apo D. In the white matter, Apo D was mainly detected in oligodendrocytes and extracellularly in the neuropil. The results of the present study support a specific behavior for each astrocyte type. These findings suggest that Apo D expression may be cell-specific, depending on the particular tissue physiology at the time of examination.
A
Apo D is a member of the lipocalin superfamily and is a component of high-density plasma lipoprotein (HDL). It is also present in breast cyst fluid, lachrymal secretions, apocrine axillary secretions, cerebrospinal fluid (CSF), and urine (Rassart et al. 2000). In peripheral tissues, Apo D is widely expressed, mainly by fibroblasts, in a large variety of mammal tissues (Boyles et al. 1990b; Provost et al. 1990; Smith et al. 1990; Séguin et al. 1994). Increments of Apo D in the peripheral nervous system (PNS) have been related to the repair process after a lesion (Boyles et al. 1990a). It has been suggested that Apo D probably has a role in binding and transporting cholesterol and its esters during regeneration, in association with other apolipoproteins such as Apo E and Apo A-I (Spreyer et al. 1990). However, Apo D protein is also able to bind other molecules, such as as arachidonic acid, porphyrins, and steroid hormones (Rassart et al. 2000). In the CNS, Apo D is localized in neurons, oligodendrocytes, astrocytes, and in pial and perivascular cells (Boyles et al. 1990b; Smith et al. 1990; Patel et al. 1995; Navarro et al. 1998,2001). The increment of Apo D levels in some neuropathologies (Suresh et al. 1998; Terrisse et al. 1998; Kalman et al. 2000; Belloir et al. 2001; Thomas et al. 2001; Mahadik et al. 2002; Navarro et al. 2003) and in experimental brain lesions (Ong et al. 1997; Montpied et al. 1999) suggests that Apo D might also be a good marker for neuropathology. The expression of Apo D is apparently induced as soon as any type of injury occurs, which enables us to speculate that Apo D in the CNS may be involved in repair and regeneration secondary to neurodegeneration.
The synthesis and secretion of Apo D by primary astrocyte cultures of mouse brain have been described. Moreover, Apo D is upregulated by progesterone and oxysterol in these cultured astrocytes (Patel et al. 1995). These results suggest a potential role as a hydrophobic ligand carrier in astrocytes for Apo D and may be important in maintenance of cholesterol homeostasis during reinnervation and regeneration. On the basis of its capacity to bind cholesterol and arachidonic acid, it is probable that Apo D takes part in mechanisms that control the membrane phospholipid/cholesterol rate and therefore cellular homeostasis and viability.
Because of the important role of astrocytes in the repair and maintenance of neuronal homeostasis and structure, our aim is to study the location and distribution of Apo D in human brain glial cells by immunohistochemical (IHC) techniques to elucidate the significance of Apo D expression in the different astrocyte types of the human nervous system.
Materials and Methods
Human tissues were provided by the Pathologic Anatomy Department of The General Hospital of Asturias. Twenty subjects between 30 and 58 years old without neurological or psychological disorders were used in this study. Pieces of human brain from different areas (frontal, parietal, and occipital cortices, striatum, hippocampus, and cerebellar cortex) were obtained at autopsy, and fixed by immersion in 10% buffered formalin. After fixation, pieces were dehydrated, cleared in butyl acetate, and embedded in paraffin. Sections 10 μm thick were obtained and attached to gelatin-covered slides.
Sections were deparaffinized in xylene and rehydrated, and a double immunostaining for Apo D protein and GFAP was carried out according to the following protocol. The sections were treated sequentially with Triton X-100 (0.1%, 5 min) at room temperature (RT), washed in distilled water, treated with H2O2 (3%, 5 min) in a wet chamber at RT, washed in distilled water, and treated with PBS for 2 min. Nonspecific binding was blocked by incubation with bovine serum. Incubation with a specific antibody against Apo D (1:2000 dilution) was carried out overnight at 4C (provided by Dr. C. López-Otín, Departamento de Bioquímica y Biología Molecular, Universidad de Oviedo; see López-Boado et al. 1994 and Navarro et al. 1998). The immunoreactivity was detected with an Extravidin-biotin-alkaline phosphatase staining kit (Sigma Extra-1A). Enzymatic activity was shown by incubation with Vector blue substrate (Vector SK-5300; Burlingame, CA). Slides were rinsed in PBS, placed in a plastic Coplin jar filled with 0.01 M sodium citrate buffer (pH 6), and incubated in a household microwave oven. Microwave treatment involves completely blocking contaminating staining in the double-labeling technique, using primary antibodies from the same species (Lan et al. 1995). Incubation with a specific antibody against GFAP (Sigma G-9269, 1:1000 dilution) was carried out overnight at 4C. The immunoreactivity was detected using the Extravidin-biotin-peroxidase Staining kit (Sigma Extra-3). After several washes, peroxidase was visualized by a red AEC reaction (Sigma A-6926) (0.5 mg AEC, 50 μl dimethylformamide, 10 μl H2O2 (3%) in 940 μl acetate buffer). The sections were not counterstained and were mounted in aqueous mounting medium. The usual specificity control tests were carried out.
Results
All specimens of normal human brain used in the present work exhibited a similar pattern of IHC staining for Apo D, and the labeling was more intense in white than in gray matter in all areas sampled. As expected, strong immunoreactivity for Apo D was also detected in fibroblasts and perivascular cells of the choroid plexus and in fibroblasts of the pial layer. Moreover, Apo D staining was also found in perivascular but not in endothelial cells of blood vessels. In constant and slight staining of scattered neurons was also observed. Scattered Apo D-positive glial cells were observed in gray matter opposite to the many Apo D-positive glial cells found in white matter. To detect astrocytes, double IHC for GFAP and Apo D was achieved. GFAP-positive astrocytes were observed in all layers of the cerebral and cerebellar cortices, but mainly in white matter.
No IHC differences between sampled areas of cerebral cortex were observed. Positive staining for GFAP was found in the cytoplasm and processes of scattered astrocytes of the gray matter in cerebral cortex (Figure 1A). Some of these cells showed immunoreactivity for Apo D mainly concentrated in cytoplasmic granules (Figure 1B). These double-positive cells were mainly observed in molecular and pyramidal layers of the cerebral cortex. Apo D-positive astrocytes were most frequently observed in association with blood vessels, and their morphology resembled that of protoplasmic astrocytes that were highly comparable to those described with the Golgi technique (Figure 1B). On the other hand, the white matter appeared more stained for GFAP and Apo D than the gray matter. However, co-localization of both antibodies in astroglial cells there was not observed (Figures 1C and 1D). Strong positive immunostaining for Apo D was observed in other glial cells that were round in shape with an eccentric nucleus and few straight processes, resembling oligodendrocytes (Figure 1D). Significant immunore-activity for Apo D was found in neuropil surrounding the white matter (Figures 1C and 1D).
The three layers of the cerebellum presented a similar pattern of double immunostaining (Figure 1D). Astrocytes immunoreactive for GFAP, similar in morphology to those observed in human cerebrocortical white matter, were sparsely found throughout the cerebellar white matter. These fibrous astrocytes have no reactivity to Apo D (Figure 1F). As in cerebral white matter, the Apo D immunostaining was found mainly in oligodendrocytes (Figure 1F). GFAP-positive astrocytes were regularly distributed throughout the granular layer of the cerebellar cortex. Only a few of these cells showed a slight co-localization of both proteins. Most of the astroglial cells were Apo D-immunonegative (Figure 1G). Sparsely distributed Apo D-immunoreactive Purkinje cells or granular cells were seen in all sampled human cerebellar cortices (Figure 1G). The other subpopulation of GFAP-positive astrocytes in the human cerebellum was identified as Bergman glia based on the position of their cell bodies, near the granular layer/Purkinje cell interface, and the extension of their feather-like processes across the width of the molecular layer to contact the pia mater. Like other astrocytes in the cerebellum, no immunoreactivity for Apo D was observed in Bergman glia (Figure 1H). Finally, the molecular layer, subjacent to the glia limitans of the cerebellum, displayed immunoreactivity for GFAP and Apo D (Figure 1H). Co-localization of both proteins was not found in this layer. GFAP staining was present only in Bergman processes, and only a few sparse neurons were slightly positive for Apo D (Figure 1H).
Discussion
The present study showed that Apo D is present in glial cells of cerebellar and cerebral cortices of normal young human brains. These results are consistent with previous IHC studies revealing that Apo D was present in mammalian neuroglial cells, astrocytes, and oligodendrocytes (Boyles et al. 1990b; Provost et al. 1990; Navarro et al. 1998; Kalman et al. 2000; Belloir et al. 2001). Moreover, the number of Apo D-positive cells in the gray matter was very low compared with that in the white matter. The main cells that express Apo D in white matter are probably oligodendrocytes, on the basis of their morphology. Fibrous astrocytes in white matter usually do not exhibit Apo D immunoreactivity, whereas reactive astrocytes in gray matter frequently express Apo D. These data suggest a differential expression for Apo D in both astrocyte subpopulations. Although oligodendrocytes are the predominant glial cells in white matter tracts, fibrous astrocytes are also present. However, in normal human brain we have observed that fibrous astrocytes of white matter are not Apo D-positive, whereas this protein is high expressed by oligodendrocytes. Therefore, oligodendrocytes might be the main cells that control the expression and role of Apo D in white matter. This hypothesis is supported by other authors who consider the white matter as the major site of Apo D gene expression in the human and rabbit nervous system, probably due to oligodendrocytes (Provost et al. 1990). The expression and possible secretion of Apo D by oligodendrocytes of the rat spinal cord was described in some studies (Boyles et al. 1990b; Schaeren-Weimers et al. 1995). Experiments in rodents show the possible implication of Apo D in the regeneration and re-myelination of nerves and consequently a role in lipid transport (Boyles et al. 1990a). One of the genes that are specifically expressed in oligodendrocytes it codes for apo D (Schaeren-Weimers et al. 1995). On the other hand, Apo D protein levels were increased in Alzheimer's disease and in the Niemann–Pick mouse, which has a genetic disorder affecting cellular cholesterol transport (Suresh et al. 1998; Kalman et al. 2000; Belloir et al. 2001). These two types of disease present common phenotypic features, neuron degeneration and white matter damage, suggesting that neural cells must be vulnerable to the disruption of cholesterol trafficking. Related to its role as a lipid-binding protein, the expression of Apo D in oligodendrocytes might be involved in binding or/and transport of the lipid molecules (cholesterol or fatty acids) necessary for the maintenance of myelin.
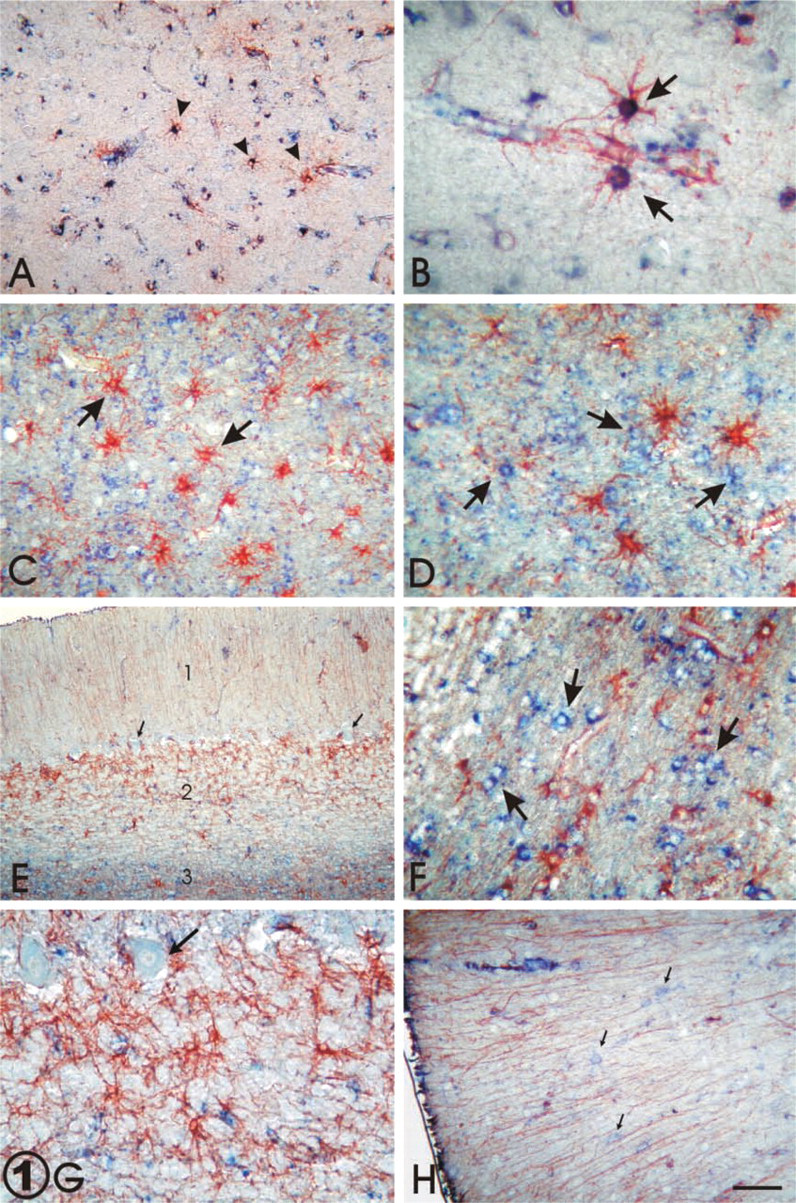
Double IHC staining for GFAP (red) and Apo D (blue). (
In human cerebral and cerebellar gray matter, we have found few GFAP-positive astrocytes, and most of them co-localized Apo D. Moreover, protoplasmic and fibrous astrocytes show clear differences in the expression of Apo D. Protoplasmic astrocytes of the gray matter show co-localization of GFAP and Apo D, especially when they are activated, whereas fibrous astrocytes do not exhibit Apo D staining. In cerebellar cortex, specialized protoplasmic astrocytes, i.e., the Bergman glia, are rarely immunopositive for Apo D. These glial cells behaved in the same way as fibrous astrocytes of the white matter. The different behavior of protoplasmic vs fibrous astrocytes, with regard to Apo D synthesis, suggests that astrocytes can be arranged in different compartments with different roles in brain homeostasis. Glial activation results in increments in the size and number of activated cells and in the expression of a new panel of genes (Topp et al. 1989; Diedrich et al. 1991). The activation of protoplasmic astrocytes is proved by the increasing amount of GFAP in their processes and cytoplasm, and recently we have shown, by non-radioactive in situ hybridization, an overexpression of Apo D in human reactive astrocytes (del Valle et al. 2003). The increased expression of Apo D, similar to that which occurs with GFAP in neurodegenerative lesions and aging (Eng 1988; Kimelberg and Norenberg 1994; Berciano et al. 1995; Raivich et al. 1999; del Valle et al. 2003), could be related to the enhanced synthesis of different transport molecules as a cellular response to stressors (Dandoy-Dron et al. 1998). Cellular changes in protoplasmic astrocytes are directly related to the formation of a physical barrier between damaged and healthy tissues (Eng 1988; Kimelberg and Norenberg 1994; Nichols 1999). Axon or dendrite injury in gray matter would activate astroglial cells and upregulate synthesis of growth factors, GFAP, and Apo D. These reactive astrocytes are progressively substituted by fibrillary astrocytes in brain lesions and this process may be controlled by cytokines and growth factors (Giulian et al. 1988; Laping et al. 1994; Asada et al. 1995; Klein et al. 1997; Ghirnikar et al. 1998; Raivich et al. 1999). IL-1 appears to play a role in modulating glial proliferation and in secretion of trophic factors and guidance of new nerve fibers (Parish et al. 2002). It is possible that Apo D is involved in repair or regeneration processes in the CNS and that its expression in activated astroglia could be upregulated by IL-1, as occurs in human breast cancer cells (Blais et al. 1994). Our results suggest that the presence of Apo D in protoplasmatic astrocytes and its absence in fibrous ones must be related to the repair and regeneration mechanisms achieved by reactive astrocytes.
The functional role of Apo D in the CNS is poorly known. Apo D is a member of the lipocalin family and is considered a “multiligand-multifunctional” protein, and it has been suggested that Apo D could achieve diverse functions in different tissues and organs. Particularly in the CNS, the role and the molecular mechanisms that control Apo D expression would be different in the various types of neural cells. Our results show a heterogeneous expression of Apo D in astrocytes, which support a specific behavior for each astrocyte type. The Apo D in protoplasmic astrocytes probably acts as a lipid transporter protein in neurites promoting responses.
Footnotes
Acknowledgements
Supported by grants from Fondo de Investigación Sanitaria Española (PI020324, Red C03/06).