
Abstract
A unique feature of the retina is the presence of photoreceptors, which require an enormous amount of oxygen for the conversion of light to an electrical signal. Hypoxia-inducible factor-1 alpha (HIF-1α) is a transcription factor that is the master regulator of cellular adaptation to low oxygen tension. Only in hypoxic conditions is HIF-1α protein stabilized and translocated to the nucleus, where it induces transcription of target genes involved in oxygen delivery and energy metabolism. We hypothesized that HIF-1α is constitutively stabilized and active in the normal human retina. We investigated the cellular distribution of HIF-1α and the expression of its downstream targets, vascular endothelial growth factor (VEGF), glucose transporter 1 (GLUT-1), and carbonic anhydrase IX (CAIX), by immunohistochemistry and immunoblotting in the retina of normal rats and human donor eyes. Both human and rat retinas displayed prominent staining of HIF-1α in nuclei of most cell types in inner and outer nuclear layers and the ganglion cell layer, a cellular distribution pattern which was confirmed in human retina by immunoblotting of nuclear extracts. A negative correlation was found between HIF-1α protein levels and postmortem times. In human retina, staining of VEGF, GLUT-1, and CAIX was found. Our observations indicate that active HIF-1 signaling occurs constitutively in the normal human and rat retina, suggesting that HIF-1 has a physiological role in the retina.
T
The outer retina, consisting of photoreceptors, is completely avascular, receiving oxygen and other essential nutrients by free diffusion from the choroid. The choroid is the best-perfused tissue in the human body, but it is barely able to provide sufficient oxygen and nutrients to meet the metabolic needs of the photoreceptors. This is illustrated by loss of dark adaptation when oxygen levels in the blood are low, for example, in healthy individuals at altitudes above 1000 m and in a variety of pathological conditions involving systemic hypoxia, such as polycythaemia vera and partial carotid occlusion (Havelius et al. 1997a,b,2000).
It has been proposed that the peculiar feature of the retina to be hypoxic in physiological conditions is an important driving force in the pathogenesis of conditions such as diabetic retinopathy and age-related macular degeneration (Lahdenranta et al. 2001; Arden et al. 2005; De Gooyer et al. 2006).
Hypoxia-inducible factor-1 (HIF-1) is a crucial player in the regulation of cellular oxidative metabolism (Semenza 2003; Ziello et al. 2007). This heterodimeric nuclear transcription factor is composed of an α and a β subunit. Both subunits are constitutively expressed in all cells, with the exception of cells in the peripheral blood, but HIF-1α is immediately degraded by an oxygen-dependent mechanism. Under conditions of hypoxia, HIF-1α is stabilized, dimerizes with HIF-1β, and translocates to the nucleus, where, depending on the cell type, it induces the transcription of a wide variety of genes, including vascular endothelial growth factor (VEGF), glucose transporter 1 (GLUT-1), erythropoietin-1 (EPO-1), carbonic anhydrase IX (CAIX), and glyceraldehyde-3-phosphate dehydrogenase (Forsythe et al. 1996; Jiang et al. 1996; Gleadle and Ratcliffe 1997; Iyer et al. 1998).
HIF-1α has mainly been investigated in relation to cancer (Bos et al. 2001; Horrée et al. 2007a,b). Owing to its presumed specificity for tumor hypoxia, it is considered to be a potential therapeutic target in cancer patients (Williams et al. 2001; Semenza 2003; Ziello et al. 2007). For years, HIF-1α was assumed to be expressed only under normal conditions in the developing embryo (Wang and Semenza 1993; Iyer et al. 1998; Zelzer et al. 1998; Talks et al. 2000; Poulaki et al. 2002). Recently, HIF-1α was also found in the nuclei of cells in a number of normal human tissue types as well: epidermis, dermal glands and hair follicles, epithelium of esophagus and colon, and cartilage (Talks et al. 2000; Giles et al. 2006; Pfander and Gelse 2007; Rosenberger et al. 2007; Boutin et al. 2008).
In the retina, HIF-1α expression has been investigated in relation to ischemia (Tang et al. 2006; Zhu et al. 2007), diabetes (Poulaki et al. 2002; Calvert et al. 2004; Kalesnykas et al. 2008), and glaucoma (Tezel and Wax 2004). However, expression in normal adult human retina has not been reported. Given the tenuous nature of oxygen tension in the retina, its possible role in retinal disease (Arden et al. 2005), and the constitutive expression in the retina of VEGF (Witmer et al. 2003) and GLUT-1 (Mantych et al. 1993), we hypothesize that HIF-1α is constitutively stabilized in the normal retina. To this end, we investigated HIF-1α distribution and the expression of its downstream target genes in normal human and rat retina.
Materials and Methods
Subjects
Donor eyes were kindly provided by the Corneabank, Amsterdam, The Netherlands, after removal of corneal buttons for transplantation. Eyes were either frozen in liquid nitrogen and stored at −80C for Western blotting (
Perfusion Fixation of Rat Retina
Experiments were carried out on male Wistar rats weighing 300–350 g. The animals were anesthetized intraperitoneally with KMA mixture [ketamine (100 mg/ml), medetomidine (1 mg/ml), atropine (0.5 mg/ml) in saline]. Either the carotid arteries were briefly perfused in situ with heparin, immediately followed by 4% para-formaldehyde under physiological pressure, by opening the jugular vein to fix the retina as rapidly as possible (
Immunohistochemistry
Paraffin-embedded human and rat retinas were sectioned (section thickness, 5 μm). Staining, using monoclonal mouse anti-HIF-1α (clone 54, dilution, 1:50; BD Transduction Laboratories, San Diego, CA), was performed as described previously (Bos et al. 2003). Polyclonal goat anti-VEGF (dilution, 1:50; R and D Systems, Abingdon, UK), polyclonal rabbit anti-GLUT-1 (dilution, 1:200; Dako, Glostrup, Denmark), and polyclonal rabbit anti-CAIX (dilution, 1:1000; Novus, Littleton, CO) were used as decribed by Horrée et al. (2007a,b). Sections were counterstained with hematoxylin. Control incubations were performed in the absence of primary antibodies.
Immunoblotting
Protein was isolated from frozen human retinas with a nuclear extraction kit (Active Motif; Rixensart, Belgium) according to the manufacturer's instructions. Twenty μg of protein was subjected to 6% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), followed by Western blot analysis. Blots were incubated overnight with anti–HIF-1α (dilution, 1:250) and anti–HIF-1β (dilution, 1:1500; BD Transduction Laboratories) as loading control. Secondary antibodies were horseradish peroxidase-conjugated goat anti-mouse IgG + IgM (dilution 1:5000; Biosource, Camarillo, CA). Enhanced chemiluminescence (Amersham Biosciences, Buckinghamshire, UK) was used for visualization as described by the manufacturer. Each sample was analyzed twice in separate runs. Intensity of HIF-1α and HIF-1β bands was quantified by densitometry (AlphaEase; AlphaInnotech Corp., San Leandro, CA), and their ratio was calculated.
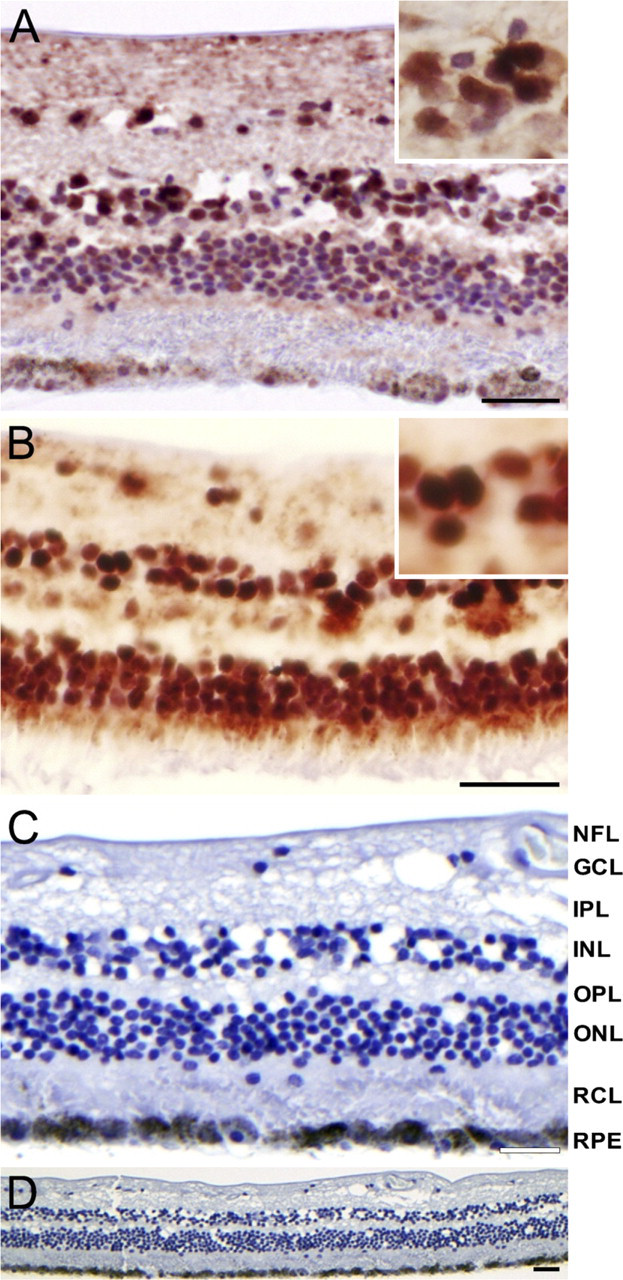
Immunohistochemical staining of hypoxia-inducible factor-1 alpha (HIF-1α) in human retina. Nuclear HIF-1α staining was present in ganglion cell layer (GCL), inner nuclear layer (INL), and outer nuclear layer (ONL) in central retina
Results
Immunohistochemical Analysis of Human Tissue Samples
Nuclear staining of HIF-1α was observed in all human retinas (Figure 1). Nuclear staining intensity increased from central to peripheral retina. At least 75% of all nuclei of the ganglion cell layer (GCL), more than 50% of nuclei in the inner nuclear layer (INL), and less than 50% of nuclei in the outer nuclear layer (ONL) were stained. Retinal staining was absent after control incubations (Figures 1C and 1D).
Downstream target proteins of HIF-1α were also expressed in normal retina (Figure 2). VEGF staining was present in cells of the GCL and in the nerve fiber layer (NFL). GLUT-1 staining was observed in the cytoplasm of cells from the outer retina toward the NFL. CAIX immunostaining was present in cells of the GCL and NFL.
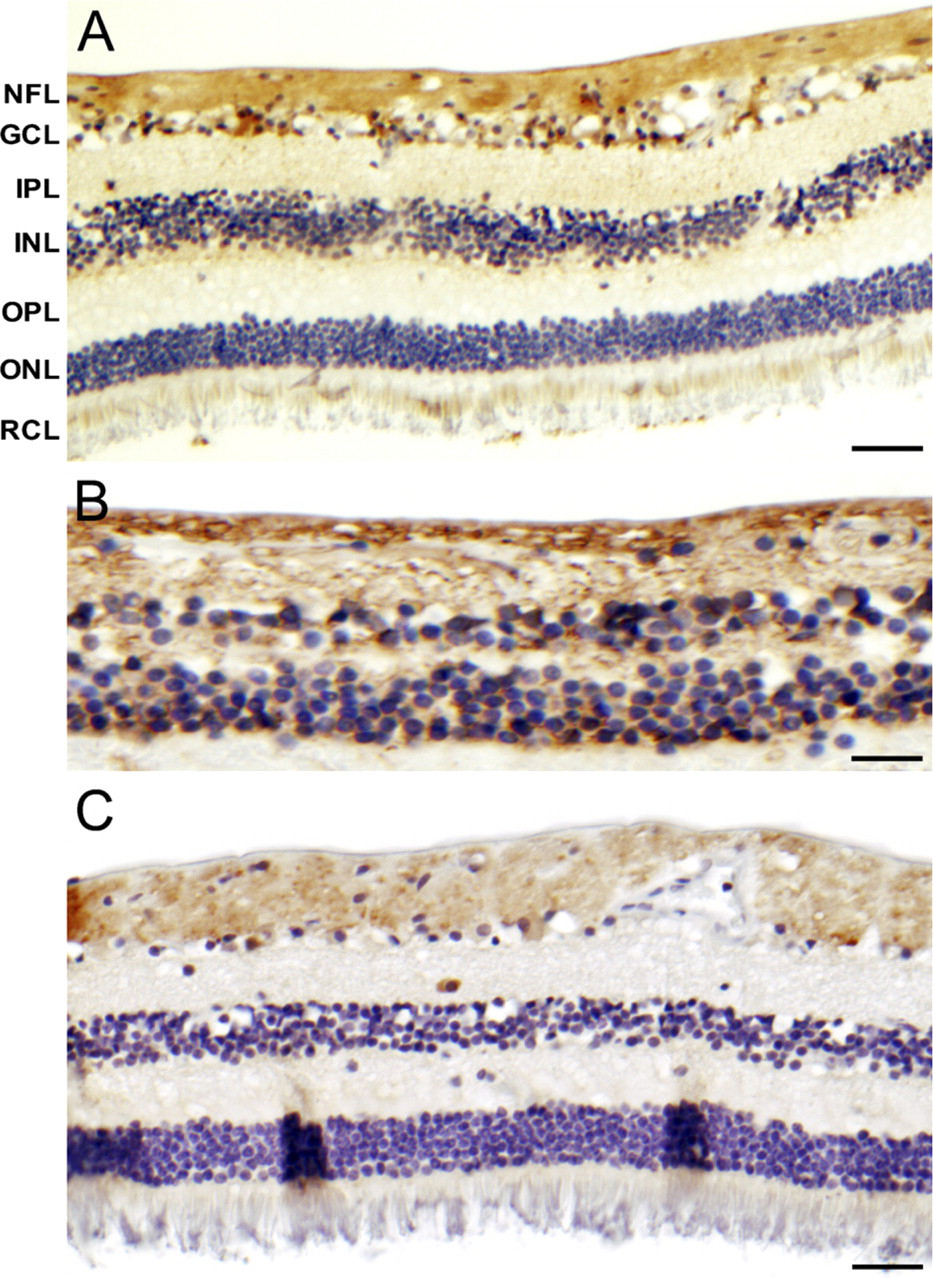
Immunohistochemical staining of vascular endothelial growth factor (VEGF)
Immunohistochemical Analysis of Rat Tissue Samples
To exclude the possibility that the nuclear localization in the human tissue samples was the result of postmortem hypoxia, rats were perfused in vivo with paraformaldehyde through the carotid artery to immediately fix the retina. The jugular vein was opened to allow the outflow of the fixative. For comparison, eyes were kept for 4 or 24 hr postmortem at room temperature. Again nuclear staining in the GCL, the INL, and the ONL of HIF-1α was present in rat retina, whereas no staining was present after control incubation (Figure 3). Overall staining was more intense, as compared with human retina, probably because a higher antibody concentration was used. In the immediately perfusion-fixed eyes, the amount of nuclei stained in the three retinal nuclear layers was similar to that in human and relatively higher than in eyes that were kept for 4 or 24 hr postmortem in phosphate-buffered saline (Figure 3).
Immunoblot Analysis of Human Retina Nuclear Extracts
To confirm the presence of HIF-1α protein in human retina, nuclear extracts of total retina of 15 patients were analyzed by immunoblotting. An example of one of the blots is shown in Figure 4A. Stabilized HIF-1α was present in various amounts in all nuclear extracts, whereas HIF-1β was present in higher and more constant amounts. The positive control of cells incubated at low levels of oxygen showed strong HIF-1α expression. Nuclear HIF-1α protein levels were negatively correlated with the postmortem time (Figure 4B). Age, sex, and disease history of the patients did not correlate with HIF-1α protein levels.
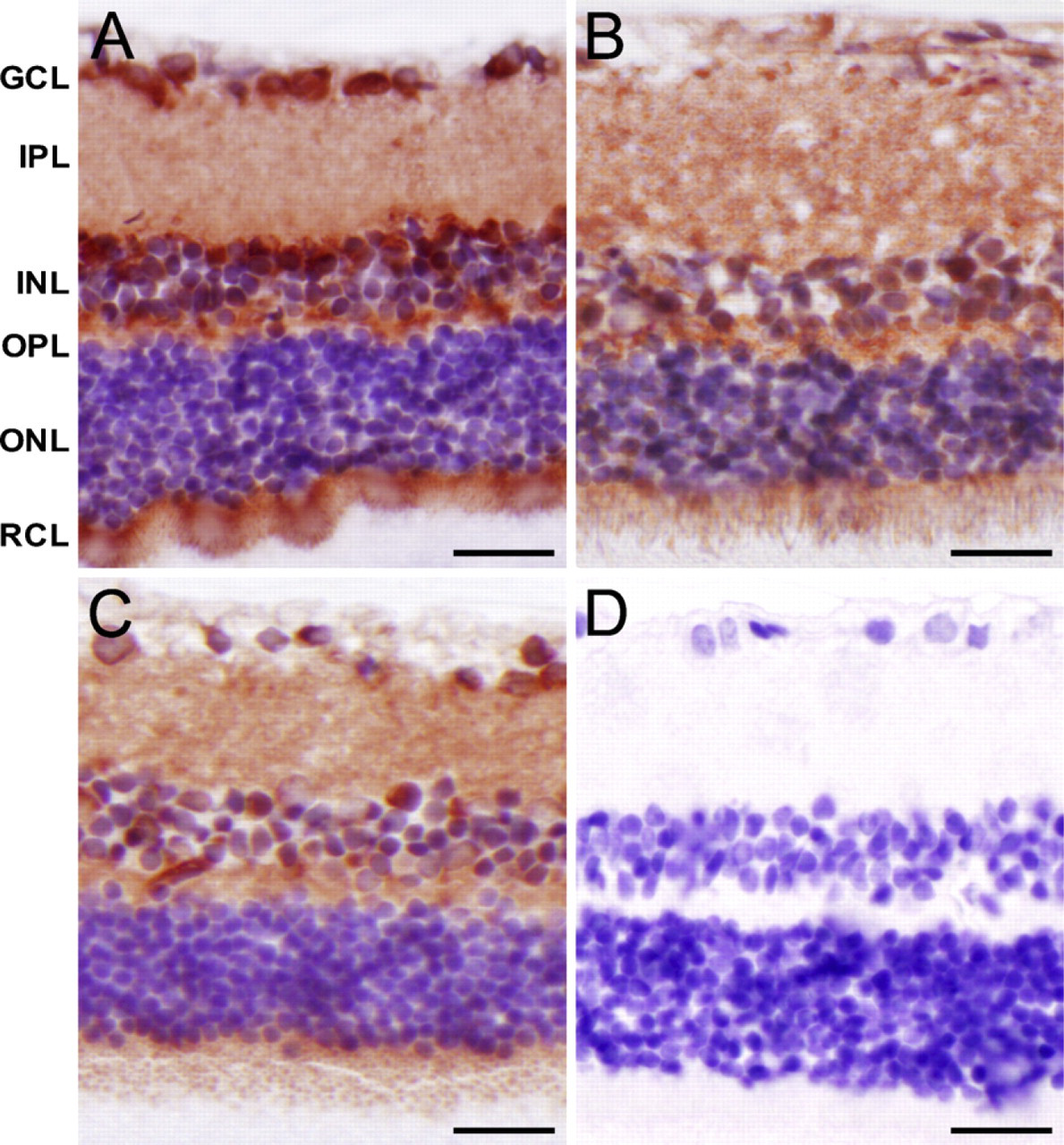
Immunohistochemical staining of HIF-1α in rat retina that was immediately perfusion-fixed
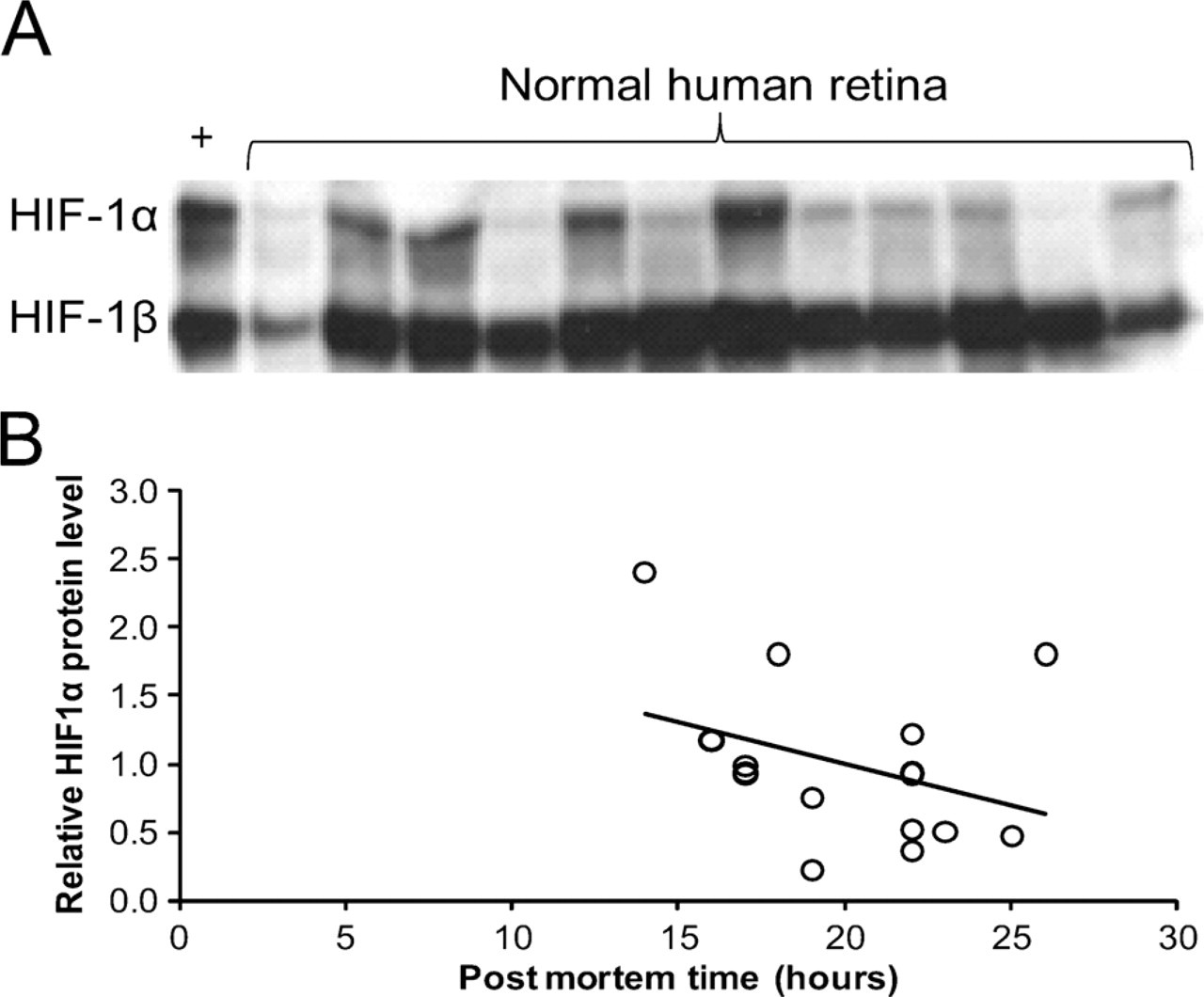
Retinal nuclear expression of HIF-1α protein.
Discussion
Stabilized HIF-1α is assumed to be present only in cellular nuclei during embryogenesis, where its activity is involved in vascular development (Iyer et al. 1998; Talks et al. 2000). In adult tissues, stabilized HIF-1α has only been found under pathological conditions associated with tissue hypoxia (Wang and Semenza 1993) and diabetes (Poulaki et al. 2002). Recently, it was reported that specific cells in certain normal epithelial and cartilaginous tissues also express HIF-1α (Giles et al. 2006; Pfander and Gelse 2007; Rosenberger et al. 2007; Boutin et al. 2008). In this study, we demonstrate that stabilized HIF-1α is also present in normal human retina. Our findings in rats confirm this, and they are in line with earlier reports showing the presence of nuclear HIF-1α protein in non-diabetic rat retina (Poulaki et al. 2002; Calvert et al. 2004; Kalesnykas et al. 2008). The nuclear staining pattern and the immunoblotting data of nuclear extracts indicate that stabilized HIF-1α was located in the nuclei. This strongly suggests that in these retinal cells, HIF-1α is dimerized with HIF-1β and able to induce gene transcription (Jiang et al. 1996).
We considered the obvious possibility that stabilization of HIF-1α is a postmortem effect due to tissue hypoxia after circulatory arrest. Although this cannot be ruled out with complete certainty by our study, it is, however, unlikely. We have never observed nuclear staining in human postmortem or surgically obtained material of other organs, irrespective of time after death, cause of death, or time before fixation or freeze-fixation (Bos et al. 2003; Horrée et al. 2007a,b). Furthermore, a similar HIF-1α staining pattern was found in the rat retina that was immediately fixed and in which postmortem effects were virtually eliminated. Interestingly, increasing postmortem times led to a decrease in nuclear HIF-1α staining in the rat retina. and likewise, a significant negative correlation was found between HIF-1α protein expression and postmortem times in human retina. Therefore, it seems more likely that stabilized nuclear HIF-1α is degraded postmortem rather than increased due to postmortem hypoxia.
The retinal expression of VEGF, GLUT-1, and CAIX, three downstream targets of HIF-1, indicates the presence of nuclear HIF-1 transcriptional activity in normal retina (Kim et al. 1999). All three proteins have cell-protective properties, especially under hypoxic conditions (Grimm et al. 2004,2006; Potter and Harris 2004; Tang et al. 2006; Bernhardt et al. 2007; Zhu et al. 2007). These three proteins had differential but partly overlapping staining patterns, with the GCL and NFL layers staining most prominently. This staining pattern is in agreement with the observed HIF-1α staining, inasmuch as the percentage of nuclei stained increases from outer to inner retina. This is also in agreement with the pO2 gradient of the retina, which declines from the outer retina to the inner retina, with a relatively hypoxic inner retina with a pO2 of only ×25 mm Hg (Wangsa-Wirawan and Linsenmeier 2003). Furthermore, these data are in agreement with those of De Gooyer et al. (2006), who demonstrated hypoxia by pimonidazole in the INL and GCL of normal mouse retina that was significantly reduced in the retina of rhodopsin knockout mice lacking rod cells.
Expression of stabilized HIF-1α in normal retina is probably due to the tenuous state of oxygen tension indigenous to the retina. The dark-adapted retina, in particular, is hypoxic because of the enormous oxygen demand of rods (Lahdenranta et al. 2001; Arden et al. 2005).
We have previously proposed that rod-driven hypoxia is an important factor in the pathogenesis of conditions such as diabetic retinopathy and age-related macular degeneration (Arden et al. 2005). Against the background of physiological hypoxia, the threshold toward pathological hypoxia is probably lowered. A small further increase of hypoxia caused by the diabetic milieu and/or a compromised vasculature is likely to have pathological consequences through upregulation of the expression of VEGF and other factors. Our present finding of stabilized HIF-1α in the retina provides evidence that even in the normal eye, the downstream biochemical effects of hypoxia are already activated, which supports the potential role of rod-driven hypoxia as a causal factor in eye disease.
We found a gradual increase in HIF-1α staining intensity from central to peripheral retina, in line with the previously reported observation that in the peripheral retina, the venous oxygen tension is lower (Alder et al. 1991; Wangsa-Wirawan and Linsenmeier 2003) and that the retinal microvasculature is more sparse than in the central retina (Toussaint et al. 1961). This suggests that physiological rod-driven hypoxia is even more profound in the peripheral retina. This may explain why in patients with diabetic retinopathy, retinal vascular occlusion and ischemia usually tend to develop first in the peripheral retina.
The large interindividual variation in nuclear HIF-1α levels in human retina can partly be explained by the variation in postmortem time (14–26 hr) and its effect on nuclear HIF-1α. Furthermore, HIF-1α is constitutively expressed in the cytoplasm and continuously degraded, unless it is stabilized by hypoxia, and it is translocated to the nucleus. Therefore, individual/local variation in oxygen levels in the human retina may affect the nuclear HIF-1α levels.
It is important to note that our findings indicate that caution should be taken when considering HIF-1α as a therapeutic target for ocular diseases such as diabetic retinopathy and age-related macular degeneration, because it appears to have a physiological role in the maintenance of retinal function. This also applies to anti-tumor therapies targeting HIF-1α.
Our findings suggest that HIF-1α not only plays a role in pathology, but also regulates cell survival, inasmuch as many of its downstream effector genes are neuroprotective. In the absence of hypoxia and subsequent HIF-1α signaling, neuroprotection may be insufficient and the retina may be more vulnerable. This may provide a new explanation for why in conditions with rod loss due to degeneration or to genetic defects such as retinitis pigmentosa, not only do the rods die, but eventually other neurons such as cones do so as well (Usui et al. 2009).
In summary, we provide evidence that HIF-1 signaling is constitutively active in the normal human retina. The retina is distinct from other tissues because of the presence of photoreceptors, which require enormous amounts of oxygen. The present study indicates that this causes not only physiological hypoxia, but also hypoxic molecular signaling via HIF-1α. These are unique features when compared with those of other normal tissues.
Footnotes
Acknowledgements
This work was supported by grants from the Dutch Diabetes Fund (grant number 1998, 131); the Edward and Marianne Blaauw Foundation; the Landelijke Stichting voor Blinden en Slechtzienden, Utrecht; the Blindenpenning Foundation, Amsterdam; the Society for the Blind, Rotterdam; and the Society for the Blind, Gelderland, The Netherlands.
The authors kindly thank Adrie Maas for perfusion of rats.