
Abstract
Multiple studies have shown that dentin matrix protein 1 (DMP1) is essential for bone and dentin mineralization. After post-translational proteolytic cleavage, DMP1 exists within the extracellular matrix of bone and dentin as an NH2-terminal fragment, a COOH-terminal fragment, and the proteoglycan form of the NH2-terminal fragment (DMP1-PG). To begin to assess the biological function of each fragment, we evaluated the distribution of both fragments in the rat tooth and bone using antibodies specific to the NH2-terminal and COOH-terminal regions of DMP1 and confocal microscopy. In rat first molar organs, the NH2-terminal fragment localized to predentin, whereas the COOH-terminal fragment was mainly restricted to mineralized dentin. In the growth plate of bone, the NH2-terminal fragment appeared in the proliferation and hypertrophic zones, whereas the COOH-terminal fragment occupied the ossification zone. Forster resonance energy transfer analysis showed colocalization of both fragments of DMP1 in odontoblasts and predentin, as well as hypertrophic chondrocytes within the growth plates of bone. The biochemical analysis of bovine teeth showed that predentin is rich in DMP1-PG, whereas mineralized dentin primarily contains the COOH-terminal fragment. We conclude that the differential patterns of expression of NH2-terminal and COOH-terminal fragments of DMP1 reflect their potentially distinct roles in the biomineralization of dentin and bone matrices.
Keywords
W
The importance of DMP1 in biomineralization has been shown through mice and human genetic studies. In mice, a lack of DMP1 results in poor mineralization of bone and dentin, whereas mutations in the DMP1 gene in humans result in osteomalacia (Ye et al. 2004, 2005; Feng et al. 2006). Interestingly, DMP1 expression was also observed in malignant tumor cells (Fedarko et al. 2001; Fisher et al. 2004; Ogbureke et al. 2007). Biochemical studies and cDNA analysis indicate that the full-length rat DMP1 protein is composed of 473 amino acids rich in Asp, Glu, and Ser. This composition shows that DMP1 is highly acidic, a property that is necessary for calcium binding because it provides a microenvironment for mineral precipitation (Qin et al. 2003).
It is nearly impossible to detect and extract the full-length protein from the extracellular matrix (ECM) from bone and dentin because of the very limited amount of intact DMP1 in both tissues. However, the ECM of both tissues does contain significant amounts of processed NH2-terminal and COOH-terminal DMP1 fragments (Qin et al. 2003). Recently, a proteoglycan form of the NH2-terminal fragment, designated as “DMP1-PG,” was discovered in bone and dentin ECM (Qin et al. 2006). Biochemical analysis showed that the COOH-terminal fragment of rat DMP1 contains 41 phosphates, whereas the NH2-terminal fragment has 12 phosphates. Tarataix et al. (2004) conducted in vitro mineralization studies and found that the COOH-terminal fragment of DMP1 extracted from rat bone acts as a hydroxyapatite nucleator. Similar data were obtained by Gajjerman et al. (2007), who used the COOH-terminal fragment of rat DMP1 in an in vitro study in the presence of type I collagen; the same group observed that the NH2-terminal fragment inhibited mineral formation. Additional, detailed studies conducted by the same group proved that DMP1 signaling functions for osteoblastic cell lineage differentiation and maturation; these authors showed that DMP1 overexpression enhances changes in cells morphology and stimulates expression of marker genes, specific for the process of biomineralization (Narayanan et al. 2001, 2006). Considering the abundance of processed DMP1 fragments in the bone and dentin ECM compared with only trace amounts of the full-length protein (unpublished data), we speculate that DMP1 processing is an activation event, converting the full-length precursor into active fragments at the correct time and place to initiate and/or regulate mineralization.
The biochemical variations of the three major DMP1 fragments found in the bone and dentin ECM indicate that they may serve distinct functions during biomineralization. A thorough assessment of their patterns of distribution during the mineralization of dentin and bone will hence provide important clues about their physiological roles, both unique and overlapping, during the process of mineralization. To gain deeper insight into the role of DMP1, we analyzed the distribution of the NH2-terminal and COOH-terminal fragments of DMP1 in rat teeth and growth plates at different ages using antibodies specific to the NH2-terminal and COOH-terminal DMP1 regions.
Materials and Methods
Antibodies
Two polyclonal antibodies were produced in rabbits using the NH2-terminal (859) and COOH-terminal (857) regions of DMP1 (Sigma Genosys; The Woodlands, TX). Oligopeptides with the sequences of LGPEEGQWGGPSKLDSDEDS (NH2-terminal, mouse DMP1 residues 101–121) and AYHNKPIGDQDDND (COOH-terminal, residues 471–485) were used. This “anti-DMP1-N antibody” called 859, was used as the standard antibody in the double-staining IHC and FRET analyses to identify the NH2-terminal fragment of DMP1. The monoclonal antibody, 8G10.3 specific for the COOH-terminal fragment of DMP1 isolated from rat bone (Baba et al. 2004) was designated as the “anti-DMP1-C antibody” and was employed in this study as the standard antibody to identify the COOH-terminal fragment of DMP1 in the IHC and Forster resonance energy transfer (FRET) analyses. The polyclonal antibody raised against oligopeptide AYHNKPIGDQDDND was designated as the “anti-DMP1-C-857 antibody” used to detect the DMP1 COOH-terminal fragment in Western immunoblots. Other primary antibodies included a monoclonal antibody, 9B6.3 (Qin et al. 2006), which specifically recognized the NH2-terminal fragments of DMP1 [including the proteoglycan form of the NH2-terminal fragment (DMP1-PG)] in Western immunoblots, along with two polyclonal antibodies that recognize either the NH2-terminal or the COOH-terminal regions of DMP1 [polyclonal antibody 784 against the NH2-terminal fragment of DMP1: gift from Dr. Lynda Bonewald of the University of Missouri, Kansas City, MO; and LF-148 polyclonal antibody against the COOH-terminal fragment of bovine DMP1, gift from Dr. Larry Fisher, National Institutes of Health (NIH)/National Institute of Dental and Craniofacial Research]. Primary antibodies used in this study are presented in Table 1.
Tissue Preparation for IHC
At the ages of 1, 7, 21, 35, and 56 days, Sprague-Dawley rats (Harlan; Indianapolis, IN) were perfused from the ascending aorta with 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4). The entire head, tibia, and humerus were dissected and immersed in the same fixative for 2 days at 4C, followed by decalcification in 8% EDTA (pH 7.4) at 4C. Samples coming from P1 animals were demineralized for 4 days, whereas those from P7 thru P56 were demineralized for 2–5 weeks. The progress of demineralization was followed with X-ray analysis. Although a good fixation achieved by perfusion with 4% paraformaldehyde followed by 48 hr of postfixation can prevent the loss of mineral-related proteins from the demineralization process, we cannot completely rule out the possibility that certain amounts of DMP1 and/or its processed fragments in the mineralized tissues might be lost during EDTA demineralization. Nevertheless, such experiments, as described in this study, can only be performed on demineralized hard tissues.
Antibodies used in this study

This polyclonal antibody was used to detect the NH2-terminal fragment of DMP1 by immunofluorescence and FRET analysis (primary utilization) and Western immunoblotting (secondary utilization) in this study. This antibody was recently generated in our laboratory (see Materials and Methods). The amino acid sequence of this immunizing oligopeptide is highly conserved among rat, mouse, bovine, and human DMP1.
This monoclonal antibody was used to detect the COOH-terminal fragment of DMP1 by immunofluorescence and FRET analysis. The characterization of this antibody has been reported (Qin et al. 2006).
This polyclonal antibody was used to detect the COOH-terminal fragment of DMP1 by Western immunoblotting analysis in this study. This antibody was recently generated in our laboratory (see Materials and Methods). The amino acid sequence of this immunizing oligopeptide is highly conserved among rat, mouse, bovine, and human DMP1.
This monoclonal antibody was used to detect the NH2-terminal fragment of DMP1 by Western immunoblotting analysis. The characterization of this antibody has been described previously (Qin et al. 2003).
The amino acid sequence of this immunizing oligopeptide is extremely highly conserved among rat, mouse, bovine, and human DMP1 (gift from Dr. Lynda Bonewald, University of Missouri, Kansas City, MO).
This polyclonal antibodies were used to detect the COOH-terminal fragment of DMP1 by Western immunoblotting analysis and were a gift from Dr. Larry Fisher of the NIH.
DMP1, dentin matrix protein 1; FRET, Forster resonance energy transfer.
The tissues were processed for paraffin embedding, and 8-μm sagittal serial sections were prepared for IHC. All the animal experiments were approved by the Baylor College of Dentistry Institutional Animal Care and Use Committee.
IHC
Reagents used for the IHC procedures were from Invitrogen (Carlsbad, CA), unless stated differently. The paraffin sections of the head, tibia, and humerus were treated with hyaluronidase (1 mg/ml) for 1 hr at 37C. After 1 hr of blocking with a solution (PBS, 2% BSA, 10% normal goat serum, and 0.1% Triton), the sections were incubated for 1 hr with the primary antibodies (both at the dilution of 1:200) at room temperature followed by 1 hr of incubation with the Alexa 488-labeled goat anti-rabbit and Alexa 546-labeled goat anti-mouse F (ab′)2 fragment secondary antibodies (1:600) at room temperature. Finally, the sections were treated with ToPro-3 (1:500) for 5 min and coverslipped with Slow Fade. Three sets of negative controls were used for the double-staining experiments. In the first set, the primary antibody was replaced with non-immunized rabbit serum. In the second set, the synthetic oligopeptide with the sequence of GLGPEEGQWGGPSKLDSDEDS (NH2-terminal) was incubated with the anti-DMP1-N antibody at a molar ratio (IgG/peptide) of 1:100 at 4C. After overnight incubation, this solution was used to replace the primary, anti-DMP1-N antibody for IHC staining. In the third set, the anti-DMP1-C antibody (8G10.3) was incubated overnight with the pure 57-kDa fragment of DMP1 isolated from rat bone at a molar ratio of 1:100 (IgG/protein) at 4C before being applied to the sections. All the sections were examined with a SP2 confocal microscope (Leica; Solms, Germany). The negative control sections and sections stained with the individual fluorophores were used to establish the settings (see below for details).
FRET Analysis
FRET analysis was used to better evaluate the distribution and potent colocalization of both types of fragments derived from the parental full-length DMP1. FRET is a biophysical technique that allows one to distinguish between the biased colocalization of two fluorophores visible on the IHC section as colocalized because of the thickness of the section (overlapping layers) and the real physical closeness of the two molecules. FRET can be used to establish the distance between two fluorophores: one called a “donor” and the other called an “acceptor” (Jares-Erijman and Jovin 2003). The emission wavelength of the donor fluorophore (Alexa 488 in this study) overlaps with the excitation wavelength of the acceptor fluorophore (Alexa 546), thus increasing the fluorescence emission of the acceptor (Konig et al. 2006). The energy transfer from donor to acceptor also causes shortening of the fluorescence lifetime and quenching of the donor; however, because the energy transfer is proportional to the inverse sixth power of the distance between the two fluorophores, FRET can be effective only when the donor and acceptor are no more than 10 nm apart. An estimate of the distance separating the donor and acceptor can be made by comparing the fluorescence efficiencies of these two fluorophores before and after photobleaching the acceptor. Photobleaching reduces the acceptor's capacity to capture the donor emission, thus increasing the measurable fluorescence intensity of the donor. The estimated distance can be used to indicate whether the two molecules (labeled with two different fluorophores) are colocalized or in different locations in vivo. To measure the FRET efficiency in this study, the acceptor photobleaching method was applied using the confocal microscope. The excitation of the donor fluorophore (Alexa 488) was performed with a 488-nm Ar laser at 25% intensity. The acceptor fluorophore (Alexa 546) was excited by means of a 543-nm NeHe laser at 51% intensity. The detection wavelength for Alexa 488 was set at 498-522 nm and for Alexa 546 was set at 558-608 nm. The photo-multiplier tube settings were fixed between 550 and 670 V. All the settings were kept constant throughout the entire experiment. To prevent potential cross-talk, each fluorophore was exposed sequentially by using only one laser at a time. The acceptor was bleached 15 times at the highest zoom (x64) at 100% power of the 543-nm laser.
All calculations for the FRET acceptor photobleaching application were performed with the Leica CLSM software, and the results were collected in the Leica system report format. At least five measurements from every structure were scored. The mean of the FRET efficiency measurements of every structure was calculated using Excel (Microsoft, Redmond, WA).
Extraction and Separation of NCPs From Bovine Predentin and Dentin (Fast Protein Liquid Chromatography) and SDS-PAGE
To obtain a sufficient amount of proteins derived specifically from the predentin and dentin, we selected bovine molars as the source of both types of tissues. The molars from a 6-month-old calf were cut in half along the long axis. The predentin layer (total, 1.2 g) was manually separated under a dissecting microscope and dissolved in an ice-cold solution of 4 M guanidium-HCl (Gdm-HCl) containing protease inhibitors. The solution was changed after 24 hr as described previously (Qin et al. 2003), and the Gdm-HCl extract of the predentin was subjected to chromatographic separation. The dentin (total, 1.2 g) was removed under the dissecting microscope, mechanically ground into pieces, and dissolved in ice-cold 0.5 M EDTA in 4 M Gdm-HCl containing protease inhibitors (twice for 24 hr). The one-step extraction protocol routinely used in our laboratory was performed with some modifications (Qin et al. 2006). The Gdm-HCl alone (predentin) and EDTA/Gdm-HCl (dentin) extracts were separated using a Q-Sepharose column (Amersham Biosciences; Piscataway, NJ) connected to a fast protein liquid chromatography (FPLC) system. The NCPs were eluted with NaCl in 6 M urea with a gradient ranging from 0.1 to 0.8 M NaCl. The protein concentration in each of the 99 collected chromatographic fractions (0.5 ml each) was assessed with the BCA kit (Pierce Biotechnology; Rockford, IL), and the proteins in each fraction were further separated with SDS-PAGE and analyzed with Stains-All staining and Western immunoblotting. Gradient gels of 5-15% SDS-PAGE were used for all the experiments.
Western immunoblots were performed using a standard protocol. Briefly, the proteins in the SDS-PAGE gels were transferred to a PVDF membrane (Immobilon F; Millipore, Bellerica, MA) at 150 V in transfer buffer (0.025 M Tris-HCl, 0.2 M glycine, 20% methanol) for 90 min. The blots were blocked at room temperature in Tris-buffered saline (TBS) containing 5% non-fat bovine milk for 2 hr and were incubated at 4C overnight in TBS containing 1.5% non-fat bovine milk and either of the primary antibodies: 9B6.3 that recognizes the NH2-terminal fragment of DMP1 or anti-DMP1-C 857 that recognizes the COOH-terminal portion of DMP1. For either of these primary antibodies, the dilution was 1:5000. The blots were washed three times for 10 min each in TTBS (TBS + 0.1% Tween-20), followed by incubation in TBS containing 1.5% non-fat bovine milk and a 1:3000 dilution of a horseradish peroxidase-conjugated secondary antibody (Cell Signaling Technology; Danvers, MA). The blots were again washed three times for 10 min each in TTBS and incubated with a color development reagent (Amersham ECL Plus Western Blotting Detection Reagent; GE Healthcare, Buckinghamshire, UK). Protein bands were assayed with a Kodak film imaging system (Rochester, NY).
Results
Immunolocalization of the NH2-terminal and COOH-terminal Fragments of DMP1 in the Tooth
Rat mandibular first molars were evaluated at different postnatal ages (days D1, D7, D14, D21, D35, and D56)
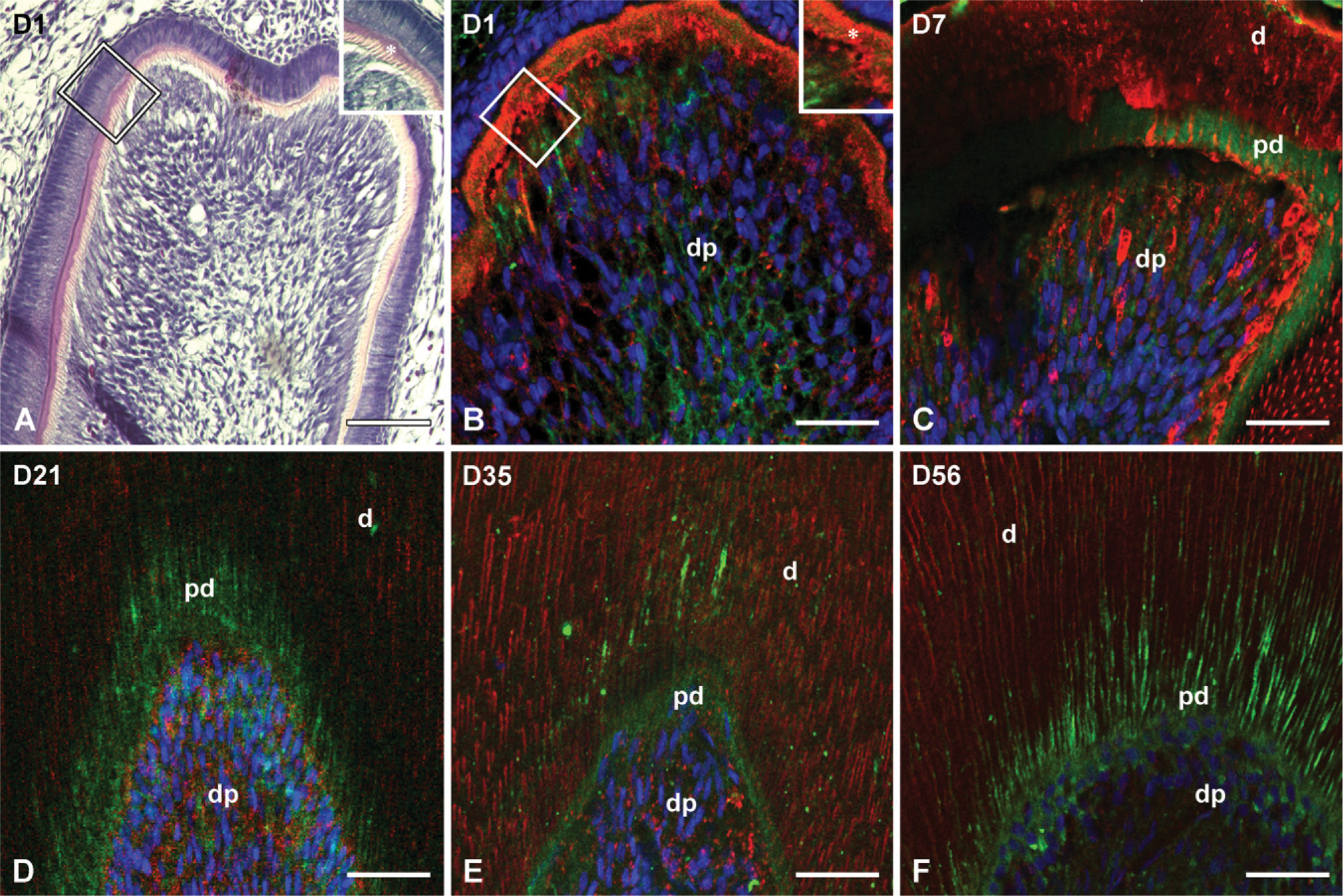
In D1, deposition of the ECM by the polarized odontoblasts was clearly noticeable at the cusp tip region, whereas the predentin (Figure 1A) layer covered up to one half of the height of the central cusp tip where only a very thin layer of mineralized dentin was visible (Figure 1A). Thus, a clear difference in the distribution of the NH2-terminal and the COOH-terminal fragments in the dentin (Figure 1B) compared with the predentin was indistinguishable (Figure 1B).
The interrupted first molar at day 7 showed a clear demarcation between the predentin and mineralized dentin layer (Figures 1C and 1D). The difference in the immunolocalization of the NH2-terminal (Figure 1) and the COOH-terminal (Figure 1) fragments of DMP1 in the predentin vs the mineralized dentin was prominent (Figure 1C). The immunoreactivity for the anti-DMP1-N antibody was observed in the odontoblasts and the predentin, whereas the dentin stained almost exclusively with the anti-DMP1-C antibody. Immunostaining for the DMP1 COOH-terminal fragment appeared as strips at advanced stages of dentinogenesis (Figures 1C-1F).
(
As postnatal development progressed at D21, the first molars reached the functional occlusion and the mineralized dentin appeared thicker. The pattern of distribution of the NH2-terminal and COOH-terminal fragments of DMP1 in the predentin and the mineralized dentin remained unchanged, however, until the end of the experiment at day 56. As shown in Figures 1D-1F, the NH2-terminal fragment of DMP1 was predominantly distributed in the predentin, whereas the COOH-terminal fragment accumulated in the mineralized dentin (Figures 1D-1F).
Immunolocalization of the NH2-terminal and COOH-terminal Fragments of DMP1 in the Growth Plate of Bone
Because the expression level of DMP1 in bone is much higher than that of the tooth, in the growth plate of rat long bone at day 1, the immunoreactivity for both the NH2-terminal and COOH-terminal fragments of DMP1 was stronger compared with the tooth. Thus, the difference in the immunodistribution of those two fragments in the bone was already distinct at day 1. The anti-DMP1-N antibody stained strongly in the resting, proliferation, and prehypertrophic zones of the growth plate but not in the ossification zone. The immunostaining appeared to concentrate in the ECM of the cartilage (Figure 2A). In contrast, the immuno-reactivity for the anti-DMP1-C antibody was primarily detected in the mineralization front and ossification zone; it mainly localized along the lamina lacuna of the primary spongiosa (Figures 2A and 2B). At day 1, the development of the secondary ossification center of the humerus epiphysis had not developed well enough to be distinguishable among the individual structures. At this stage, the cartilage of the epiphysial secondary ossification center stained exclusively with the anti-DMP1-N antibody (Figures 2A and 2B). The colocalization of the NH2-terminal and COOH-terminal fragments of DMP1 was limited to the layer of hypertrophic chondrocytes, which aligned along the mineralization front (Figure 2B).
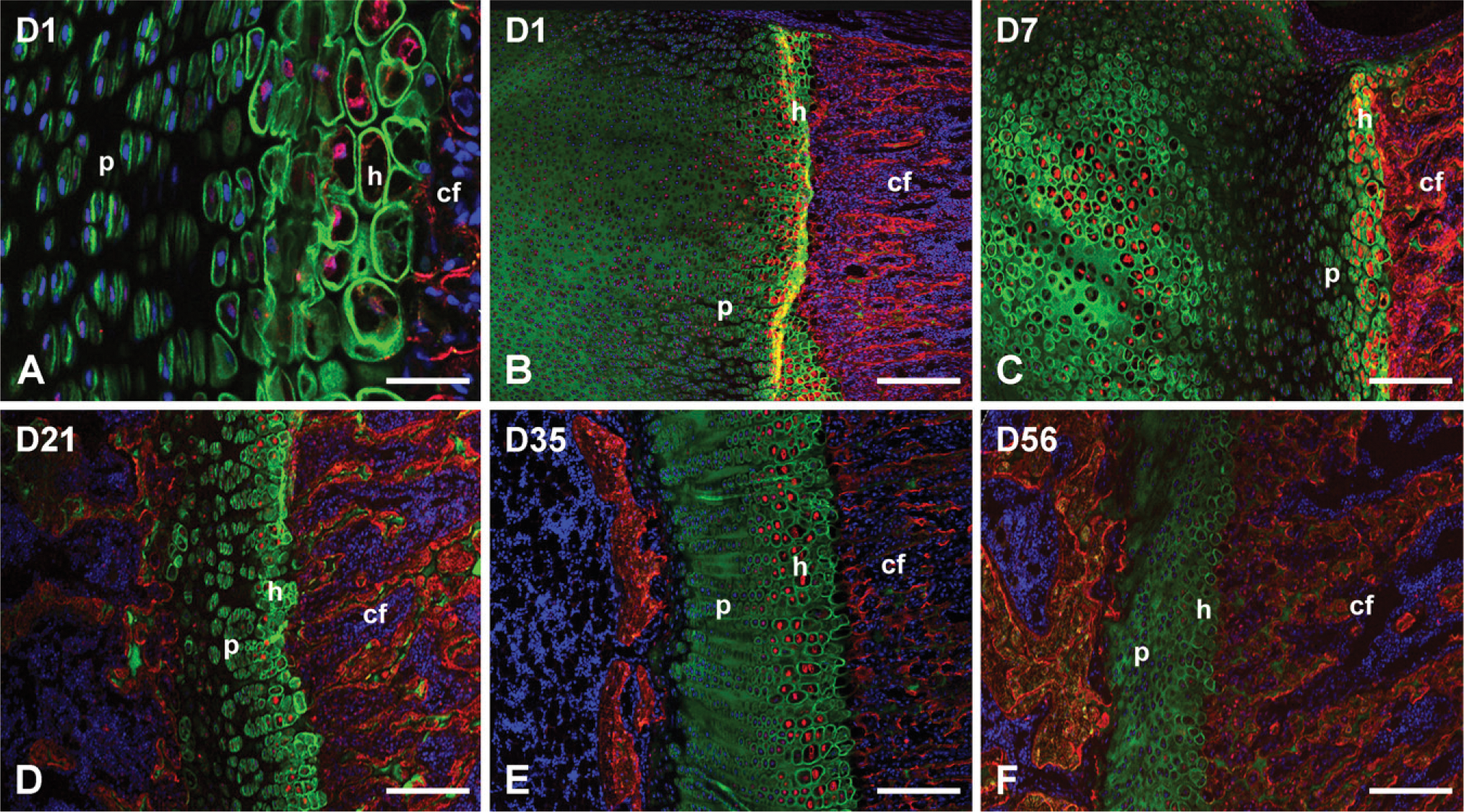
IHC staining of the NH2-terminal and COOH-terminal fragments of DMP1 in bone. Sections from the epiphyseal regions of rat humerus at the ages of day 1 (
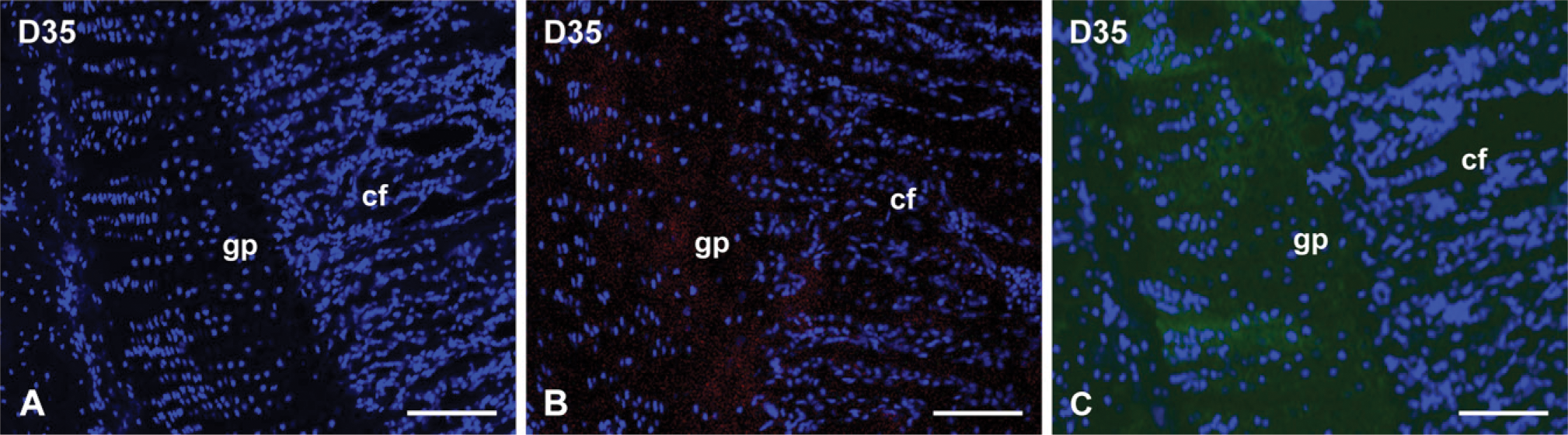
From day 7 to the end of the experiment (D56), the difference in distribution between the NH2-terminal and the COOH-terminal fragments of DMP1 in the growth plate followed the same pattern as for the 1-day-old rats (i.e., the NH2-terminal fragments of DMP1 primarily localized in the proliferating and prehypotrophic zones of the growth plate, whereas the COOH-terminal fragments accumulated in the mineralization front and the ossification zone; Figures 2C-2F). These distribution patterns were also evident in the developing secondary ossification center at day 7 (Figure 2C). No signal was detected in the negative control sections devoid of primary antibodies, (Figure 3A). In negative controls made with preincubation of the synthetic oligopeptide or purified 57-kDa fragment, a very weak signal specific for the appropriate primary antibody was detected because of incomplete blocking (Figures 3B and 3C).
FRET Analysis
The FRET analysis showed that, in the tooth, the NH2-terminal and COOH-terminal fragments of DMP1 colocalized (i.e., <10 nm apart) in the odontoblasts and predentin, whereas in the bone, they colocalized in the hypertrophic zone of the growth plate (Table 2). These results remained consistent throughout the entire observation period.
Extraction and Separation of NCPs and Western Immunoblotting Analysis of DMP1 Fragments in Bovine Predentin and Dentin
Q-Sepharose chromatography separated the NCPs of predentin or dentin extract into 99 fractions (Figures 4A and 4B). NCPs extracted from predentin eluted from the Q-Sepharose column as two sharp peaks, whereas those extracted from dentin eluted as three peaks. The widest middle peak detected in the dentin extracts did not show up in the extracts derived from predentin. The subsequent SDS-PAGE and Western immunoblotting analyses of all extracted fractions showed the differences in the distribution of the NH2-terminal and COOH-terminal fragments of DMP1 in the bovine predentin and dentin. Fractions 75-93 of the predentin extracts contained broad smears (Figure 5A) that were immunoreactive to the anti-DMP1-N antibody and to the monoclonal antibody 9B6.3. Such broad molecular mass components (eluting in later fractions) corresponded to the proteoglycan form of the NH2-terminal fragment of rat DMP1 (Qin et al. 2006). Protein smears were not observed in the corresponding fractions of the dentin extracts (Figure 5B). Western immunoblotting of predentin chromatographic fractions 51-69 showed an ∼40-kDa protein band that was immunoreactive with the anti-N-terminal fragment monoclonal antibody (Figure 5C); this protein band corresponded with the 37-kDa (NH2-terminal) fragment of rat DMP1. In the corresponding chromatographic fractions of dentin extract (Figure 5D), this ∼40-kDa protein band was much weaker than in those fractions from the predentin extract. Western immunoblotting using antibodies against the COOH-terminal fragment of DMP1 for predentin extract showed the presence of an ∼70-kDa protein band in fractions 51-69, which was immunoreactive with both the anti-DMP1-C 857 and LF-148 antibodies, whereas in the corresponding fractions (51-69) from the dentin extract, this ∼70-kDa protein band was much stronger than that in the predentin (data not shown).
(
FRET analysis
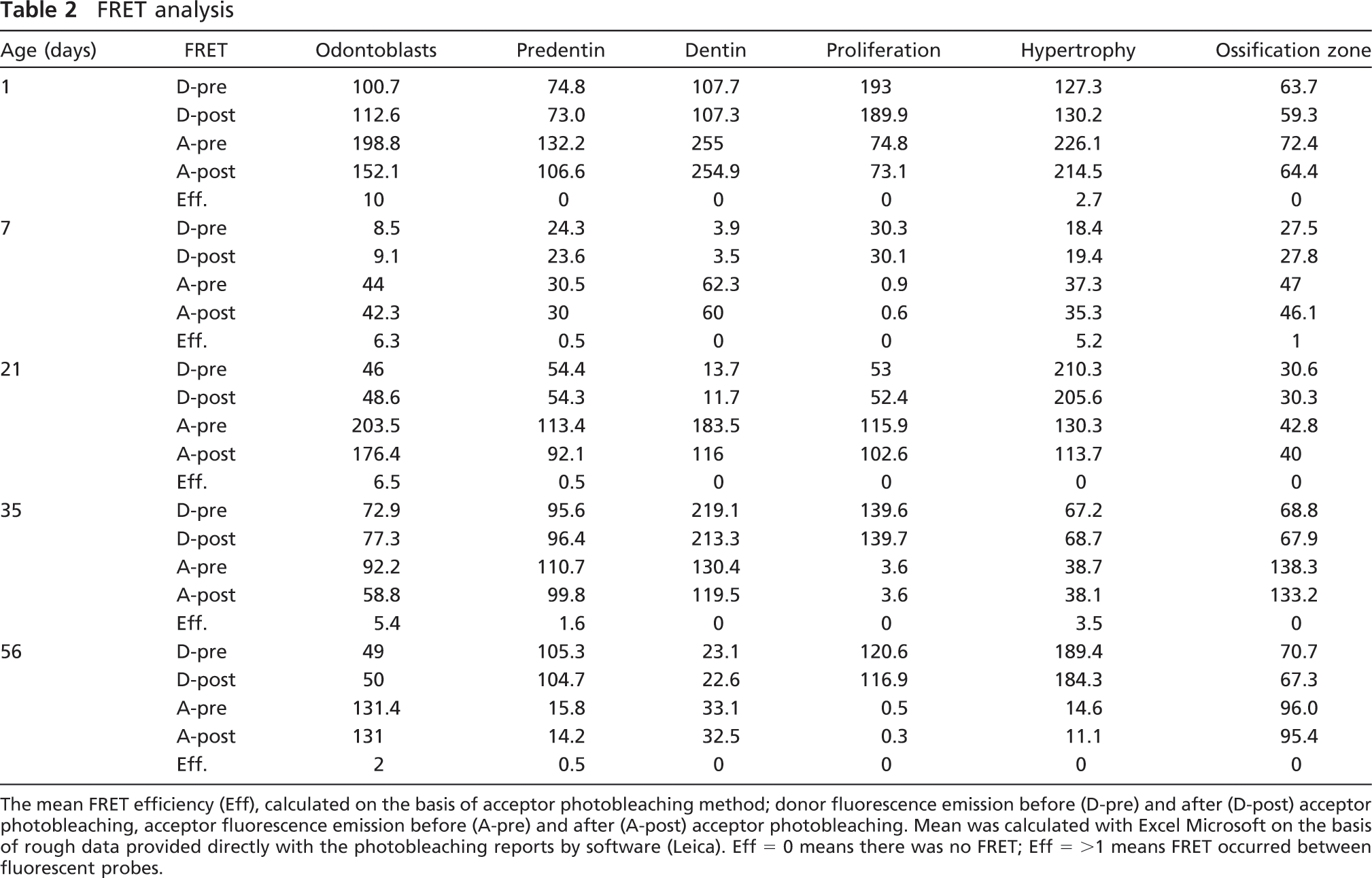
The mean FRET efficiency (Eff), calculated on the basis of acceptor photobleaching method; donor fluorescence emission before (D-pre) and after (D-post) acceptor photobleaching, acceptor fluorescence emission before (A-pre) and after (A-post) acceptor photobleaching. Mean was calculated with Excel Microsoft on the basis of rough data provided directly with the photobleaching reports by software (Leica). Eff = 0 means there was no FRET; Eff = >1 means FRET occurred between fluorescent probes.
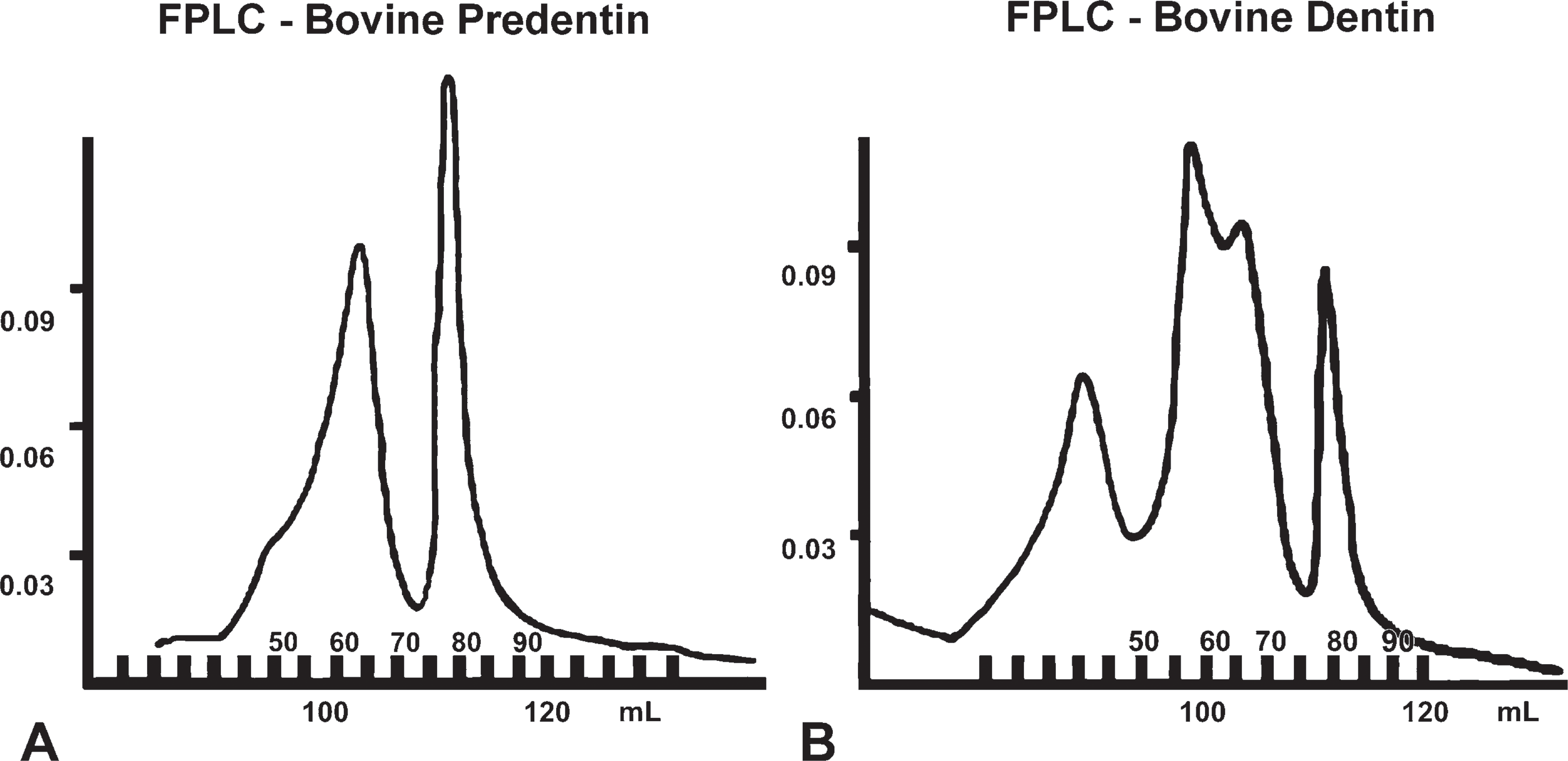
Q-Sepharose chromatographic separation [Forster resonance energy transfer (FPLC)] of non-collage-nous proteins (NCPs) from the extracts of predentin (A) and dentin (
Discussions
The findings from this study suggested that the two NH2-terminal and COOH-terminal fragments of DMP1 may play different roles in dentinogenesis and endochondral osteogenesis.
Distribution of the COOH-terminal fragment of DMP1 suggests a role in the dentin and bone mineralization. Our IHC and biochemical findings clearly showed that the COOH-terminal fragment of DMP1 is limited to the mineralization zones. Our findings were strengthened by FRET analysis and align with several in vitro observations that showed that the COOH-terminal fragment isolated from rat bone acts as a promoter for hydroxyapatite formation (Tartaix et al. 2004; Gajjeraman et al. 2007). Moreover, our observations are consistent with the amino acid sequence and protein chemistry analyses, which indicated that the DMP1 COOH-terminal fragment would be suitable for biomineralization. This fragment contains 41 phosphorylated, negatively charged serine residues, which are targets for calcium ion precipitation and subsequent mineral deposition (Qin et al. 2004). The fact that dentin phosphoprotein (DPP, a COOH-terminal fragment of dentin sialophosphoprotein), another highly phosphorylated protein known to be a strong promoter for hydroxyapatite formation, is preferentially localized at the mineralization front between predentin and dentin (Butler and Ritchie 1995) provides further indirect support to our hypothesis that the COOH-terminal fragment of DMP1 promotes biomineraliztion.
In the tooth, the pattern of the COOH-terminal fragment distribution was similar to that of DPP, which was previously confirmed to be the promoter of dentin mineralization (Butler et al. 2002). In neonate rats, the COOH-terminal fragment of DMP1 accumulated at the predentin-dentin border and spread to the mineralizing regions of dentin, which aligns the previously reported observation by Massa et al. (2005). In the older groups, staining of several sections showed the striplike appearance that suggests that the peritubular dentin might stain stronger with the anti-DMP1-C antibody compared with the intertubular dentin.
In the epiphyseal growth plates and the secondary ossification center of a long bone, the COOH-terminal fragment seems to be much more abundant in the mineralization front and the ossification zone of the epiphysis. These areas are rich in the degenerating ECM of the cartilage and are targets for the vascular invasion and subsequent cascade of events leading to final mineralization. In the hypertrophic chondrocytes, the COOH-terminal fragment seemed to concentrate inside the cells. Presence of DMP1 inside the cells might reflect the regulatory function of the protein reported by Nayaranan et al. (2001), which showed DMP1 direct stimulation of the expression of the mineralization-related genes alkaline phosphatase (ALP), osteocalcin (OC), and bone sialoprotein (BSP). Moreover, since Ye et al. (2005) reported on the impact of DMP1 on chondrocyte apoptosis, it seemed possible that the COOH-terminal fragment might be involved in the apoptosis signaling pathway. Clearly further studies involving the functional analysis of molecules engaged in the cascade of events leading to the programmed cells death in hypertrophic chondrocytes are warranted.
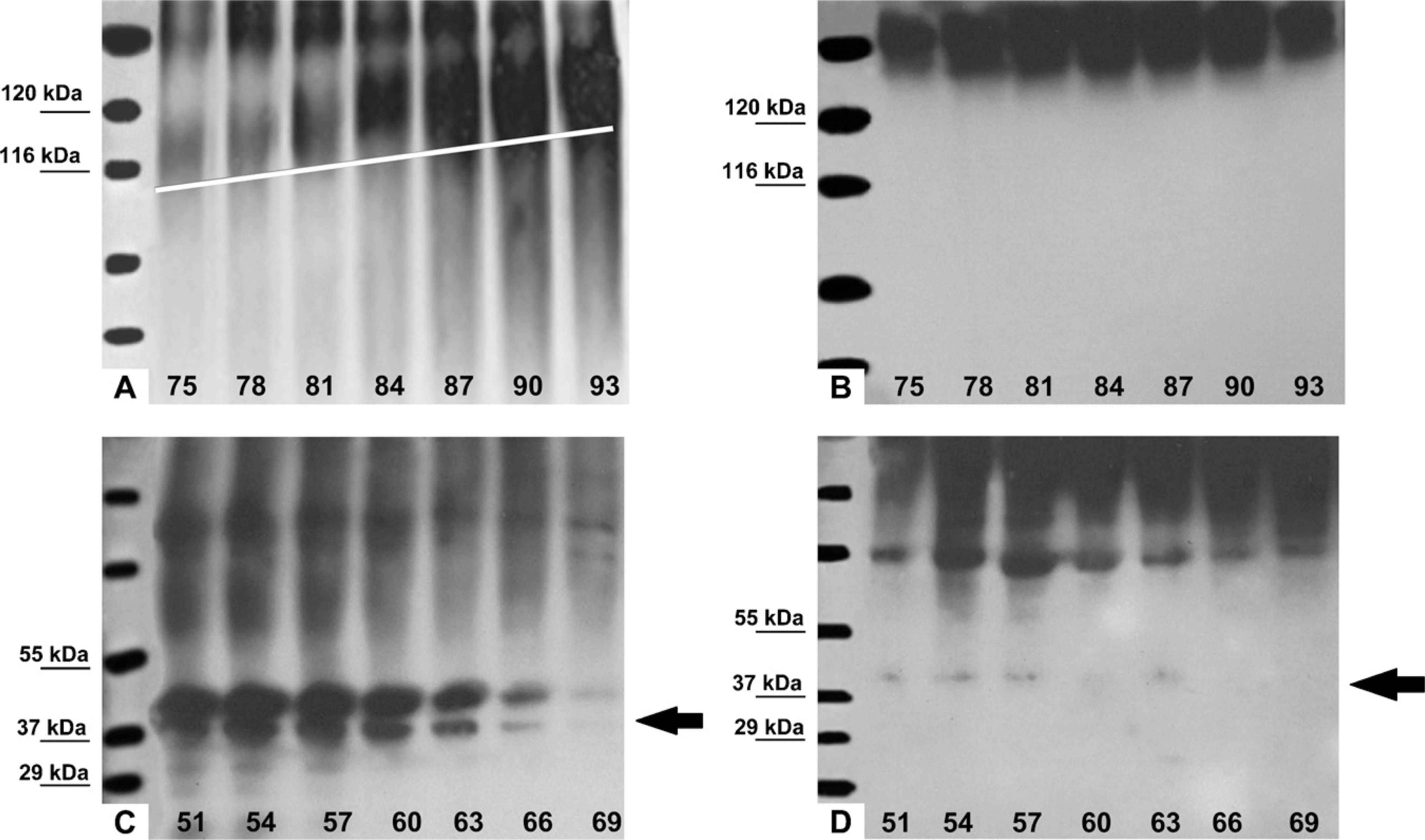
Western blot analyses of the distribution of DMP1 fragments in bovine predentin vs dentin using an antibody specific to the NH2-terminal fragment of DMP1. (
To date, DMP1 studies have seemed to focus on the role of the COOH-terminal fragment, because its biochemical properties are more suitable for biomineralization. To the best of our knowledge, we are reporting the detailed distribution of the NH2-terminal fragment in dentin and bone for the first time. The finding that the specific distribution of the NH2-terminal fragment of DMP1 was consistently restricted to the non-mineralized regions of the tooth predentin and the growth plate of bone (proliferation and prehypertrophic zone) seems to correlate with its biochemical properties and putative function. Moreover, the distribution pattern followed the age-dependent changes of the narrowing growth plate. Further analysis of the biochemical results showing the abundance of DMP-PG in the fractions extracted from the predentin led us to the working hypothesis that the proteoglycan fragment of DMP1 (DMP-PG) may be the biologically important form of the NH2-terminal fragment, whereas the “pure” 37-kDa fragment may be a remnant remaining after DMP-PG degradation. We postulate that the DMP1-PG secreted into the predentin, along with other proteoglycans, prevents this tissue in some manner from being mineralized too rapidly. Before the mineralization of the collagen fibrils and the conversion of the predentin to dentin, a major portion of the DMP1-PG is metabolized or removed. Some of it remains as the 37-kDa fragment. Thus, there is very little or no DMP1-PG in the mineralized dentin. This theory corroborates previous studies using [35S] sulfate labeling, which showed that as much as ∼50% of nascent proteoglycans were lost when the predentin was converted to dentin (Prince et al. 1984; Steinfort et al. 1994; Butler and Ritchie 1995). This hypothesis was further supported by the observation that the gradient concentration of chondroitin 4-sulfate decreases toward the dentin layer where mineralization increases (Takagi et al. 1990; Septier et al. 2001). It seems very likely that DMP1-PG interacts with other proteoglycans in the organization and maturation of the collagenous scaffold of the cartilage and predentin to appropriately prepare the spatially oriented sites for mineral deposition. Thus, the counterbalancing effect of the NH2-terminal and the COOH-terminal fragment of DMP1 may play an important role to some degree in dentinogenesis and osteogenesis. The additional FRET analysis showing the lack of colocalization of the NH2-terminal fragment and the COOH-terminal fragment of DMP1 in the resting and proliferation zones of the growth plate provided strong evidence that the NH2-terminal fragment of the DMP1 accumulates exclusively in the non-mineralized portions of bone. The DMP1 NH2-terminal fragment distribution limited to the resting, proliferating, and hypertrophic chondrocytes of metaphysis resembles the distribution of the matrix Gla protein. Matrix Gla protein is expressed in the zones of endochondral ossification and cartilage acting as an inhibitor of mineralization and preventing osteoblasts from overmineralization (Schreiweis et al. 2007). This body of evidence suggests that the NH2-terminal fragment of DMP1 may act as a mineralization inhibitor or cartilage protector against the too rapid, premature mineralization promoted by the COOH-terminal fragment of DMP1.
The IHC double-staining method used in this study allowed the NH2-terminal and the COOH-terminal fragments of DMP1 to be viewed in the same area, which when coupled with the results of the FRET analysis, can confirm or deny the evidence of their physical colocalization. The use of both methods showed that, in the tooth, the NH2-terminal fragment of DMP1 colocalized with the COOH-terminal fragment in the odontoblasts and predentin; however, the clear difference between both fragments distribution in the tooth became distinguishable at the later stage at 7 days, which is consistent with the developmental expression of DMP1 that has been previously reported (Hao et al. 2004). Because the odontoblasts synthesize DMP1, the coexistence of both fragments in these cells indicates the presence of the full-length protein at its origin. Our study showed that the NH2-terminal and COOH-terminal fragments of DMP1 colocalize in the predentin and tend to separate near the dentin, implying that the majority of the full-length DMP1 may be processed in the cells. It is likely that, after cleavage, the highly phosphorylated, negatively charged COOH-terminal fragment of DMP1 is immediately entrapped in the spatially oriented collagenous ECM framework and is involved in calcium binding and mineral nucleation. Thus, in our IHC study, the COOH-terminal fragment was distributed predominantly in the mineralizing dentin. In contrast to the COOH-terminal fragment, the proteoglycan form of the NH2-terminal fragment may contribute to the creation of the hydrated interfibrillar network of the predentin and protect it against mineralization. Based on our chronological studies, we speculate that the pulp-oriented recession of the mineralization front caused by gradual dentin deposition likely initiated a cascade of enzymatic events, resulting in the degradation of the GAG chain of DMP-PG. Based on the fact that matrix metalloproteinase-3 is involved in the degradation of proteoglycan core (Hall et al. 1999; Boukpessi et al. 2008) and the release of NCPs from the dentin matrix), we speculate that these two proteases may act in concert to drive the process of proteoglycan degradation. It is also possible that the increasing concentration of calcium ions stimulated the in situ activity of the chondroitinases (Prabhakar et al. 2006), subsequently catalyzing the glycosidic linkage between the hexosamines and uric acid and leading to the gradual shortening of the GAG chain until its final degradation. Consequently, the trace amount of the NH2-terminal fragment observed in the dentin probably indicated a “pure” 37-kDa fragment, which was likely the product of DMP-PG degradation.
Because of the stronger level of DMP1 expression in bone, the difference in both fragment distributions was relatively more striking at every chronological stage. In bone, the NH2-terminal and COOH-terminal fragments of DMP1 colocalized merely in the layer of hypertrophic chondrocytes bordering the mineralization front, the area where endochondral bone formation takes place; during this process, the hypertrophic chondrocytes die, leading to cartilage matrix degradation. The subsequent vascular invasion delivers osteoblast progenitors that initiate the deposition of bone after differentiation into the osteoblasts (van Munster et al. 2005). It seems reasonable to believe that, in the hypertrophic chondrocyte zone resembling predentin, both fragments of DMP1 “compete” for dominance at the border of the mineralization front. Thus, we believe that the colocalization of the NH2-terminal and COOH-terminal fragments of DMP1 in the hypertrophic chondrocyte zone signals bone production and the consequent invasion of the COOH-terminal fragment. As bone nucleation and mineralization progress, chondroitin kinase hydrolyzes the GAG chain of DMP-PG, and this proteoglycan of the NH2-terminal fragment is gradually withdrawn from the mineralizing bone.
Until now, our understanding of the role of DMP1 in biomineralization has been focused on its effects on mineral nucleation, growth, and maintenance, functions that seem to be associated with the COOH-terminal fragment. However, the biological roles of the NH2-terminal fragment have been largely ignored. Our studies suggest that the role of the NH2-terminal fragment may be essential for both dentin and bone mineralization. It seems very likely that both fragments work together in a sophisticated balance that controls the progress of biomineralization in vivo.
Footnotes
Acknowledgements
This work was supported by NIH Grant DE005092 (to CQ).
Dr. Izabela Maciejewska is on sabbatical leave at Baylor College of Dentistry from the Department of Dental Prosthodontics, Medical University of Gdansk, Poland.