
Abstract
The aim of this study was to evaluate the abundance of the organic anion transporter 5 (Oat5) and the sodium-dicarboxylate cotransporter 1 (NaDC1) in kidney and urine after renal ischemic reperfusion injury. Renal injury was induced in male Wistar rats by occlusion of both renal pedicles for 0 (Group Sham), 5 (Group I5R60), or 60 (Group I60R60) min. The studies were performed after 60 min of reperfusion. The expression of Oat5 and NaDC1 was evaluated by IHC and Western blotting. Oat5 and NaDC1 abundance and alkaline phosphatase activity (AP) were assayed in urine. A decreased expression in renal homogenates and apical membranes and an increase in urinary excretion of Oat5 and NaDC1 were observed in I60R60 rats, as well as alterations of other widely used parameters for renal dysfunction and injury (plasma creatinine, urinary AP activity, kidney weight, histological lesions). In contrast, in the I5R60 group, only an increase in urinary excretion of Oat5 and mild histopathological damage was detected. This is the first study on Oat5 and NaDC1 detection in urine. These results suggest that urinary excretion of Oat5 might be an early indicator of renal dysfunction, which is useful for detection of even minor alterations in renal structural and functional integrity.
R
Drugs and metabolites are eliminated from the body by metabolism and excretion. The kidney makes the major contribution to excretion of unchanged drugs and also to excretion of metabolites. Net renal excretion is a combination of three processes: glomerular filtration, tubular secretion, and tubular reabsorption. Several transport proteins are implicated in the renal tubular secretion and reabsorption of endogenous and exogenous compounds (Wright and Dantzler 2004). It has been reported that the expression of several transport proteins is downregulated in the presence of ischemia and reperfusion: e.g., organic anion transporters 1 and 3 (Oat1 and Oat3) (Kwon et al. 2007; Matsuzaki et al. 2007: Schneider et al. 2007; Di Giusto et al. 2008), type 3 Na+/H+ exchanger (NHE-3) (Wang et al. 1998; Kwon et al. 2000; Du Cheyron et al. 2003), type II Na-Pi cotransporter (NaPi-II) (Kwon et al. 2000; Rubinger et al. 2005), and aquaporins (Fernández-Llama et al. 1999; Kwon et al. 1999).
Oat5 from rats and mice has been cloned and characterized in an in vitro expression system as an organic anion/dicarboxylate exchanger, exhibiting a cation-independent, probenecid-sensitive transport capacity for ochratoxin A (OTA), estrone-3-sulfate (ES), and dehydroepiandrosterone sulfate (DHEAS), which was inhibited by some sulfate but not glucuronide conjugates (Youngblood and Sweet 2004; Anzai et al. 2005, 2006; Kwak et al. 2005). In addition, the mouse Oat5, but not the rat Oat5, was found to transport salicylate, whereas both transporters showed no significant affinity for p-aminohippurate (PAH) and various other anionic and cationic drugs (Youngblood and Sweet 2004; Anzai et al. 2005; Kwak et al. 2005). Rat Oat5 interacts with the five-carbon dicarboxylate α-ketoglutarate, but also with the four-carbon dicarboxylate succinate, and with chemically heterogenous anionic compounds, such as non-steroidal anti-inflammatory drugs, diuretics, bromosulfophthalein, and penicillin G (Anzai et al. 2005). RT-PCR studies in isolated tubules from the rat kidney localized the Oat5 mRNA predominantly in proximal tubule S2 and S3 segments (S3>S2) (Anzai et al. 2005), whereas, by IHC, the Oat5 protein was localized to the brush border of proximal tubule straight segment (S3) in the outer stripe of the rat and mouse kidney (Anzai et al. 2005; Kwak et al. 2005).
The sodium-dicarboxylate cotransporter 1 (NaDC1) protein is located on the apical membrane of the S1, S2, and S3 segments of the proximal tubule and in the small intestine (Sekine et al. 1998; Pajor 2006). The primary function of this transporter is to reabsorb filtered Krebs cycle intermediates, particularly citrate (Pajor 2006; Ho et al. 2007). The regulation of urinary citrate concentrations is of considerable importance because of the role of citrate as a calcium chelator. Hypocitraturia is associated with a tendency to form kidney stones (Pajor 2000). He et al. (2004) have shown that increased NaDC1 expression on the renal proximal tubule epithelial cells could play an important role in nephrolithiasis development. The activity of NaDC1 of the renal proximal tubule is increased in metabolic acidosis (Aruga et al. 2000), potassium depletion (Levi et al. 1991), and starvation (Pajor 2000).
It has been proposed that NaDC1 and Oat5 have an important role in the late (S2 to S3) segments of the proximal tubules; because the apical membrane Oat5 expression level is high and the basolateral membrane Oat3 expressions level is low, some organic anions, such as steroid sulfates that are glomerular filtrated or tubular secreted or effluxed by Mrp2/4 and/or Npt1(OATv1), are reabsorbed by Oat5 using the outward gradient of succinate and α-ketoglutarate generated by NaDC1 (Anzai et al. 2005).
In vitro studies have shown that renal proximal tubules can be salvaged from hypoxia-/reoxygenation-induced mitochondrial injury by the supplement of Krebs cycle intermediates during either hypoxia or reoxygenation (Weinberg et al. 2000a, b). Considering that Oat5 and NaDC1 are both involved in the transport of Krebs cycle intermediates and that little is known about the regulation of Oat5 and NaDC1, especially during pathological states, it was of interest to evaluate their abundance in urine and kidneys from rats with renal ischemic reperfusion injury.
Materials and Methods
Experimental Animals
Male Wistar rats from 110 to 130 days old were used throughout the study. All animals were allowed free access to standard laboratory chow and tap water and were housed in a constant temperature and humidity environment with regular light cycles (12 hr) during the experiment. All animals were cared for in accordance with the principles and guidelines for the care and use of laboratory animals (National Institutes of Health, Guide for Care and Use of Laboratory Animals, Publication 86-23, National Institutes of Health, Bethesda, MD, 1985). Animals were anesthetized with sodium thiopental (70 mg/kg body weight, IP). Both kidneys were exposed through flank incisions, mobilized by being dissected free from the perirenal fat. Both renal pedicles were occluded with a smooth surface vascular clip for 0 (Group Sham), 5 (Group I5R60, model of bilateral mild subclinical ischemia), or 60 min [Group I60R60, model of bilateral ischemia with acute renal failure (ARF)] (Mishra et al. 2003). Total ischemia was confirmed by observing blanching of the entire kidney surface. During the period of ischemia, the wound was closed temporarily to maintain body temperature. After the clips were removed, the kidneys were observed for an additional 2-5 min to ensure color change indicating blood reflow, and the wound was closed with 3-0 silk and surgical metal clamps. All the studies were performed after 60 min of reperfusion.
Measurement of Arterial Pressure
Systolic arterial pressure was measured in both experimental groups with a Harvard indirect rat tail blood pressure monitor (Harvard Apparatus; Millis, MA).
Biochemical Determinations
Blood was withdrawn from the femoral artery. At the end of the experiments, urine was sampled from the bladder. The urine samples were used for analyses of transporters abundance, alkaline phosphatase (AP), and creatinine concentrations. Serum samples were used to measure creatinine levels as indicative parameter of global renal function. Serum and urine creatinine levels were determined using a commercial kit (Creatinina; Wiener Laboratory, Rosario, Argentina).
Histopathological Studies
A set of experimental animals were used for histopathological studies. Histopathology of kidneys was performed after fixing the kidneys in 10% neutral-buffered formaldehyde solution for 4 hr and embedding in paraffin; 4-μm-thick sections were processed for routine staining with hematoxylin-eosin. A semiquantitative scoring system was used, similar to that previously described (He et al. 2004). The pathological alterations were graded as 0, 1, 2, 3: 0 = no visible lesions; 1 = mild dilation in some tubules without decreased cellular height; 2 = obvious dilation of many tubules and decreased cellular height; 3 = severe dilation of most tubules, flattened tubular epithelium, loss of brush border membranes, luminal debris, and cytoplasmic vacuolization.
Preparation of Apical Membranes From Kidney
Apical membranes were isolated from kidneys by Mg/EGTA precipitation as previously described (Montagna et al. 1998; Torres et al. 2003), with some modifications. The kidneys were cut off, minced, and homogenized in 30 g/100mL (v/w) of ice-cold 50 mM mannitol, 2 mM Tris HCl buffer (pH 7.10), and 5 mM EGTA for 5 min at top speed in a Sorvall Omnimixer. Homogenate samples were separated for analyses of marker enzymes activities, transporters abundance, and total protein levels. MgCl2 was added to the homogenate to a final concentration of 12 mM, and the mixture was stirred in an ice bath for 15 min. The homogenate was centrifuged (3000 × g, 15 min, 4C). The supernatant was carefully decanted and centrifuged again at 28,000 × g for 40 min at 4C. The pelleted material representing brush border membrane was resuspended in 50 mM manitol-10 mM HEPES-Tris (pH 7.50) and centrifuged for 15 min at 800 × g at 4C. The supernatant was finally centrifuged for 45 min at 28,000 × g. The brush border membrane pellets thus obtained were resuspended in 50 mM manitol-10 mM HEPES-Tris buffer (pH 7.40). Aliquots of the membranes were stored immediately at −70C for 2 weeks. Each preparation represented renal tissues from four animals. The enrichment and purity of these membranes were comparable to those previously reported (Montagna et al. 1998; Torres et al. 2003). Protein quantification of samples was performed using the method of Sedmak and Grossberg (1977).
Electrophoresis and Immunoblotting
Homogenates (50 μg of protein), apical membranes (40 μg of protein), and urine (10 μl) samples were boiled for 3 min in the presence of 1% 2-mercaptoethanol/2% SDS. Samples were applied to an 8.5% polyacrylamide gel, separated by SDS-PAGE, and electroblotted to nitrocellulose membranes. To verify equal protein loading and transfer between lanes, Ponceau red and/or antibody against human β actin were used as previously described (Shah et al. 2001; Rubinger et al. 2005; Brandoni et al. 2006a, b; Matsuzaki et al. 2007; Di Giusto et al. 2008). The nitrocellulose membranes were incubated with 5% non-fat dry milk in PBS containing 0.1% Tween 20 (PBST) for 2 hr. After being rinsed with PBST, the membranes were incubated with a rabbit polyclonal antibody against rat Oat5 (at a dilution of 1:800) and with commercial mouse monoclonal antibody against human β actin (1.25 μg/ml) or with a rabbit polyclonal antibody against rat NaDC1 (at a dilution of 1:800) with commercial mouse monoclonal antibody against human β actin (1.25 μg/ml). The specificity of Oat5 and NaDC1 antibodies has been described elsewhere (Sekine et al. 1998; Anzai et al. 2005). The membranes were incubated for 1 hr with a peroxidase-coupled goat anti-rabbit IgG (Bio-Rad; Hercules, CA) after further washing with PBST. Blots were processed for detection using commercial kit (ECL enhanced chemiluminescence system; Amersham, Buckinghamshire, UK).
IHC Microscopy
Kidneys from the three experimental groups of rats were briefly perfused with saline, followed by perfusion with periodate-lysine-paraformaldehyde solution (0.01 M NaIO4, 0.075 M lysine, 0.0375 M phosphate buffer, with 2% paraformaldehyde, pH 6.20), through a cannula introduced in the abdominal aorta. The kidney slices were immersed in periodate-lysine-paraformaldehyde solution at 4C overnight. The tissue was embedded in paraffin. Paraffin sections were cut. After deparaffining, the sections were incubated with 3% H2O2 for 15 min (to eliminate endogenous peroxidase activity) and then with blocking serum for 30 min. The sections were incubated with non-commercial polyclonal antibodies against Oat5 (diluted 1:100) or NaDC1 (diluted 1:500) overnight at 4C. The sections were rinsed with PBST. The sections were incubated with horseradish peroxidase (HRP)-conjugated secondary antibody against rabbit immunoglobulin for 1 hr. To detect HRP labeling, a peroxidase substrate solution with diaminobenzidine (0.05% diaminobenzidine in PBST with 0.05% H2O2) was used. The sections were counterstained with hematoxylin before being examined under a light microscope.
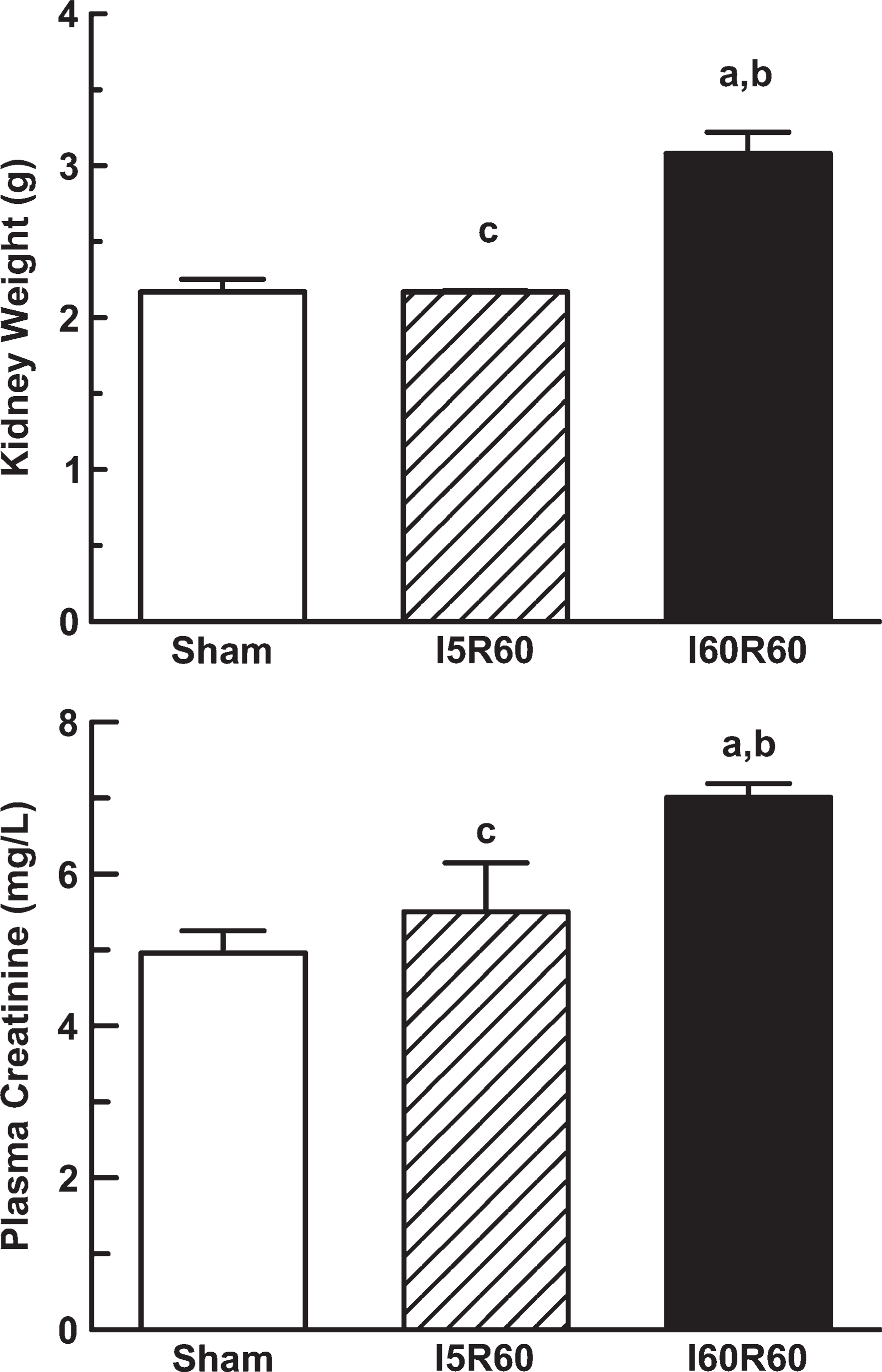
Kidney weight and creatinine plasma levels in Sham (n = 6), I5R60 (n = 3), and I60R60 (n = 4) rats. Results are expressed as mean values ± SEM. a p<0.05 vs Sham. b p<0.05 vs I5R60. c p<0.05 vs I60R60.
Materials
Chemicals were purchased from Sigma (St. Louis, MO) and were analytical grade pure. Mouse monoclonal antibody against human β actin was purchased from Alpha Diagnostic International (San Antonio, TX).
Statistical Analysis
Statistical differences between groups were evaluated by one-way ANOVA followed by the Newman-Keuls test. p<0.05 was considered statistically significant. The values are expressed as the means ± SEM. For these analyses, GraphPad software (San Diego, CA) was used.
For densitometry of immunoblots, samples from kidneys of I5R60 and I60R60 rats were run on each gel with corresponding Sham kidneys. The results were normalized for β actin density. The abundance of Oat5 and NaDC1 in the samples from the experimental animals was calculated as percentage of the mean Sham control value for that gel.
Results
Rats with bilateral mild subclinical ischemia showed kidney weights and plasma creatinine levels that were indistinguishable from Sham animals. In contrast, kidney weight and creatinine plasma levels increased significantly in the I60R60 group in comparison to the Sham rats as shown in Figure 1, reflecting significant renal dysfunction and kidney injury. No modifications were observed in body weight or systolic arterial pressure in all the experimental groups (data not shown).
By light microscope, mild dilatation in some tubules (score grade = 1) occurred in the I5R60 group. Severe dilation of most tubules, flattened tubular epithelium, loss of brush border membranes, luminal debris, and cytoplasmic vacuolization (score grade = 3) were observed in the I60R60 group as previously described in this experimental model (Brady et al. 1996; Green et al. 2000) (Figure 2).
As shown in Figure 3, there was a significant decrease in Oat5 abundance in the homogenates and in apical membranes from I60R60 rats. NaDC1 abundance also showed a statistically significant decrease in renal homogenates and apical membranes from the I60R60 group (Figure 4). No modifications were observed in Oat5 and NaDC1 abundance in the homogenates and in apical membranes after 5 min of bilateral ischemic-reperfusion injury as shown in Figures 3 and 4, respectively.
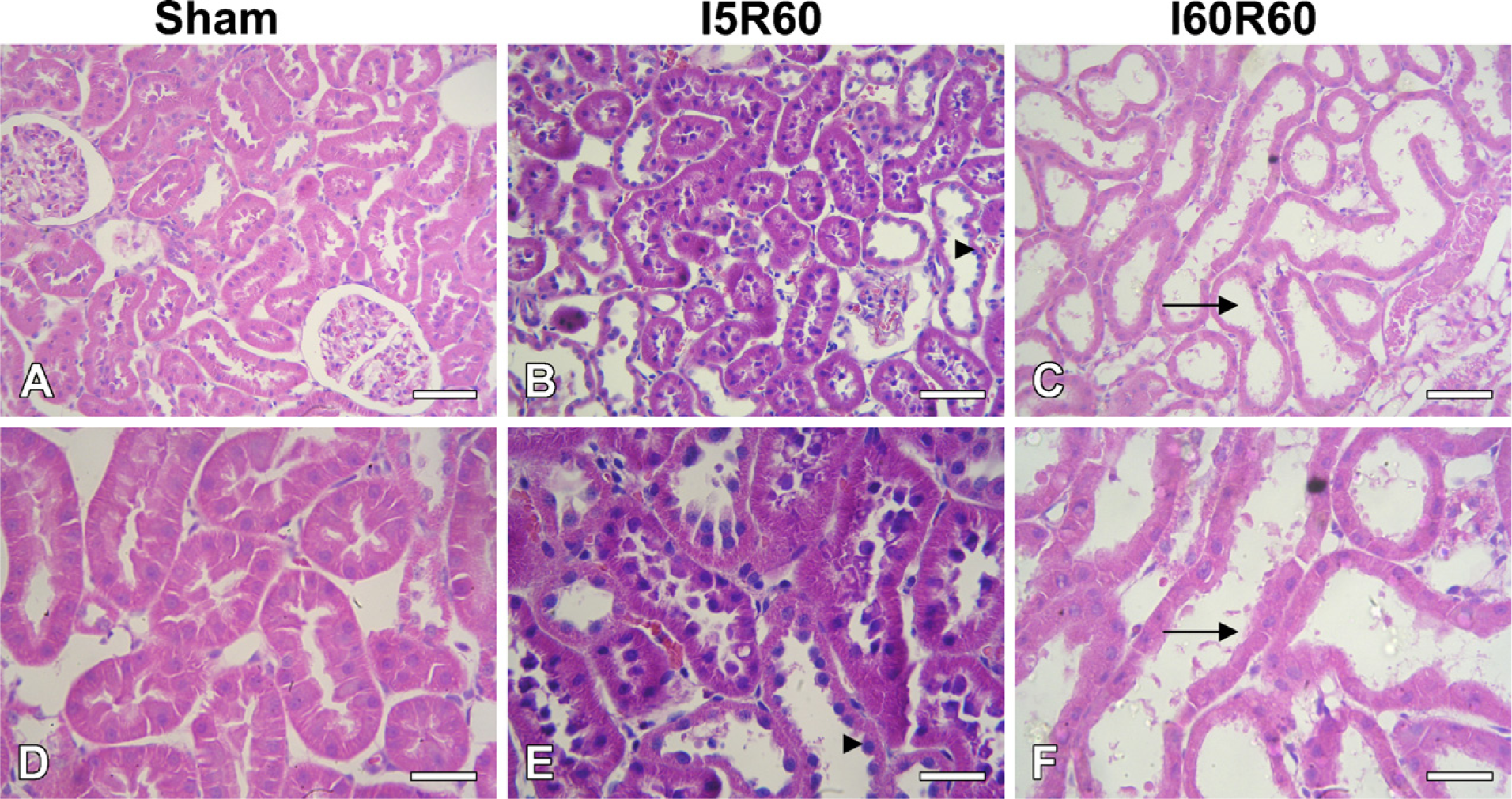
Histological studies in Sham (
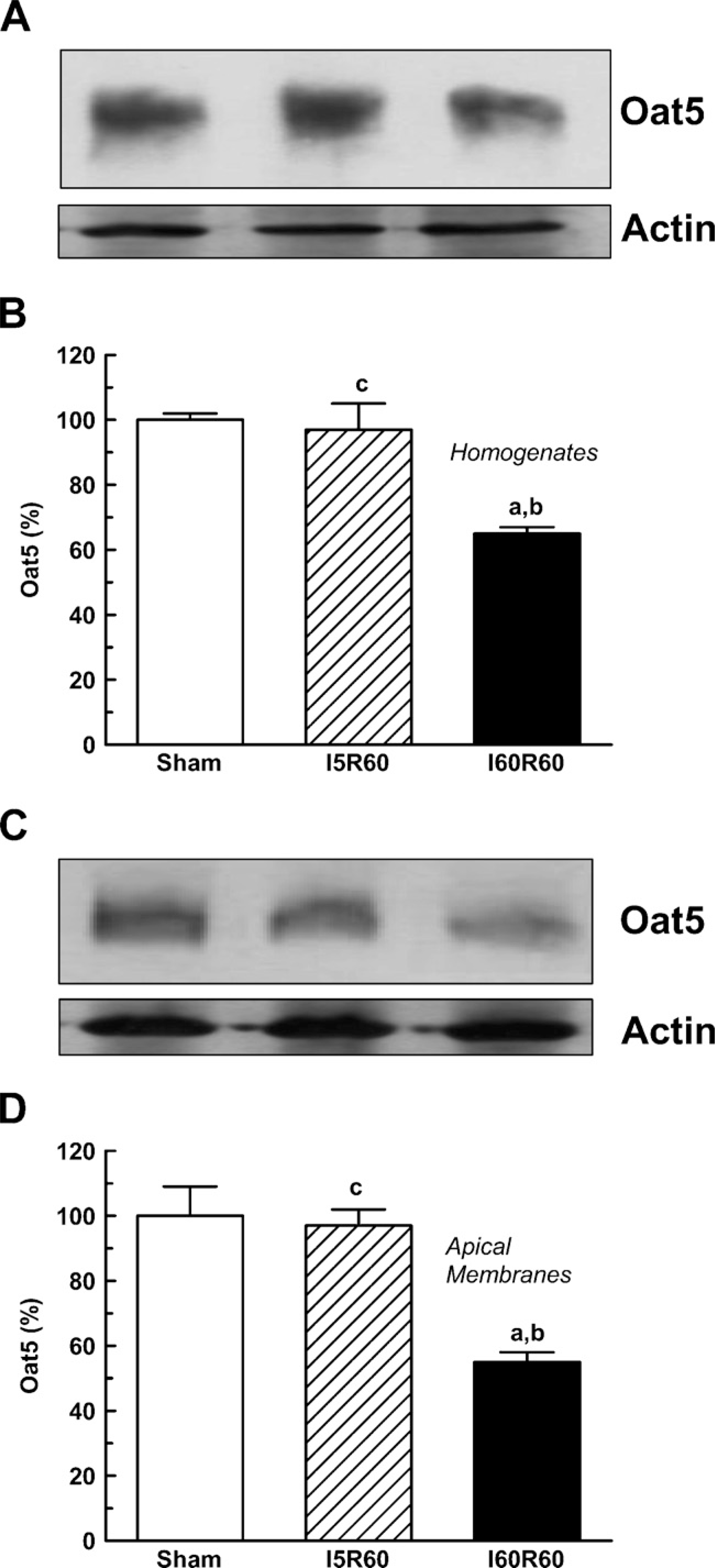
Renal homogenates (50-μg proteins) (
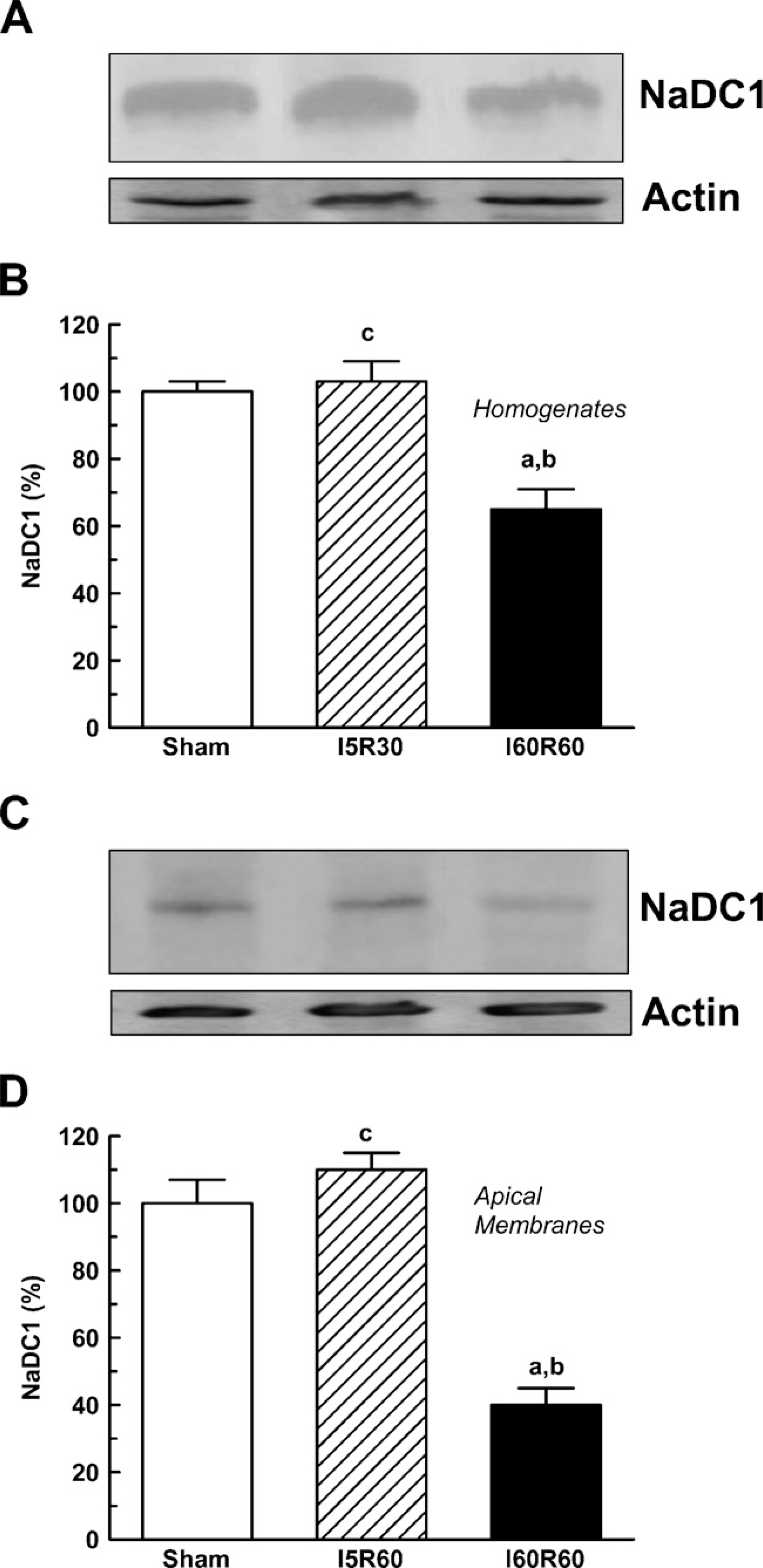
Renal homogenates (50-μg proteins) (
In Sham-operated rats, strong Oat5 labeling was associated with the apical plasma membrane domains in proximal tubule cells. Oat5 staining of proximal tubule cells was normal in I5R60 kidneys and was reduced in I60R60 ones (Figure 5). Oat5 labeling was consistent with the density observed by immunoblotting in each experimental group.
IHC showed abundant labeling of NaDC1 associated with apical plasma membranes of proximal tubule of Sham and I5R60 rat kidneys (Figure 6). In contrast, NaDC1 labeling was substantially weaker in I60R60 kidneys, with proximal tubules showing significantly reduced labeling of apical membranes. These results corroborated the data obtained by Western blotting.
Oat5 and NaDC1 abundance in urine was related to urinary creatinine concentrations to correct for variations in urine production as previously described for urinary transporters and enzymes (Heiene et al. 2001; Umenishi et al. 2002). Measurements of biomarker abundance alone are insufficient because normal physiological variations in water excretion can dilute or concentrate urinary proteins. Normalization on the basis of total protein amount is generally unsatisfactory because total protein excretion can vary broadly among various pathological states. Creatinine is excreted in the urine at relatively constant rates allowing it to be used to normalize urinary excretion of a particular protein. Figure 7A shows that Oat5 abundance was significantly higher in urine samples from the I5R60 and I60R60 groups (38% and 60%, respectively). In contrast, NaDC1 abundance in urine was comparable to Sham values in I5R60 rats and was significantly higher (65%) in I60R60 animals (Figure 7B). The activity of AP in urine has been widely used as a biomarker for renal dysfunction and injury (Price 1982; Heiene et al. 2001). Therefore, the urinary activity of AP was measured relative to urinary creatinine concentrations and compared with urinary Oat5 and NaDC1. As shown in Figure 7C, the activity of AP was only significantly increased in urine samples from I60R60 rats.
Discussion
ARF, caused either by renal ischemia or nephrotoxic agents, is typically characterized by oliguria, a severe reduction in glomerular filtration rate, and a variable fall in renal blood flow. The pathophysiology of ischemic ARF is complex and is not well defined. Structural and biochemical changes in the postischemic kidney that result in vasoconstriction, desquamation of tubular cells, intralaminar tubular obstruction, and transtubular backleakage of glomerular filtrate are pathophysiological mechanisms that have been reported (Brady et al. 1996; Green et al. 2000).
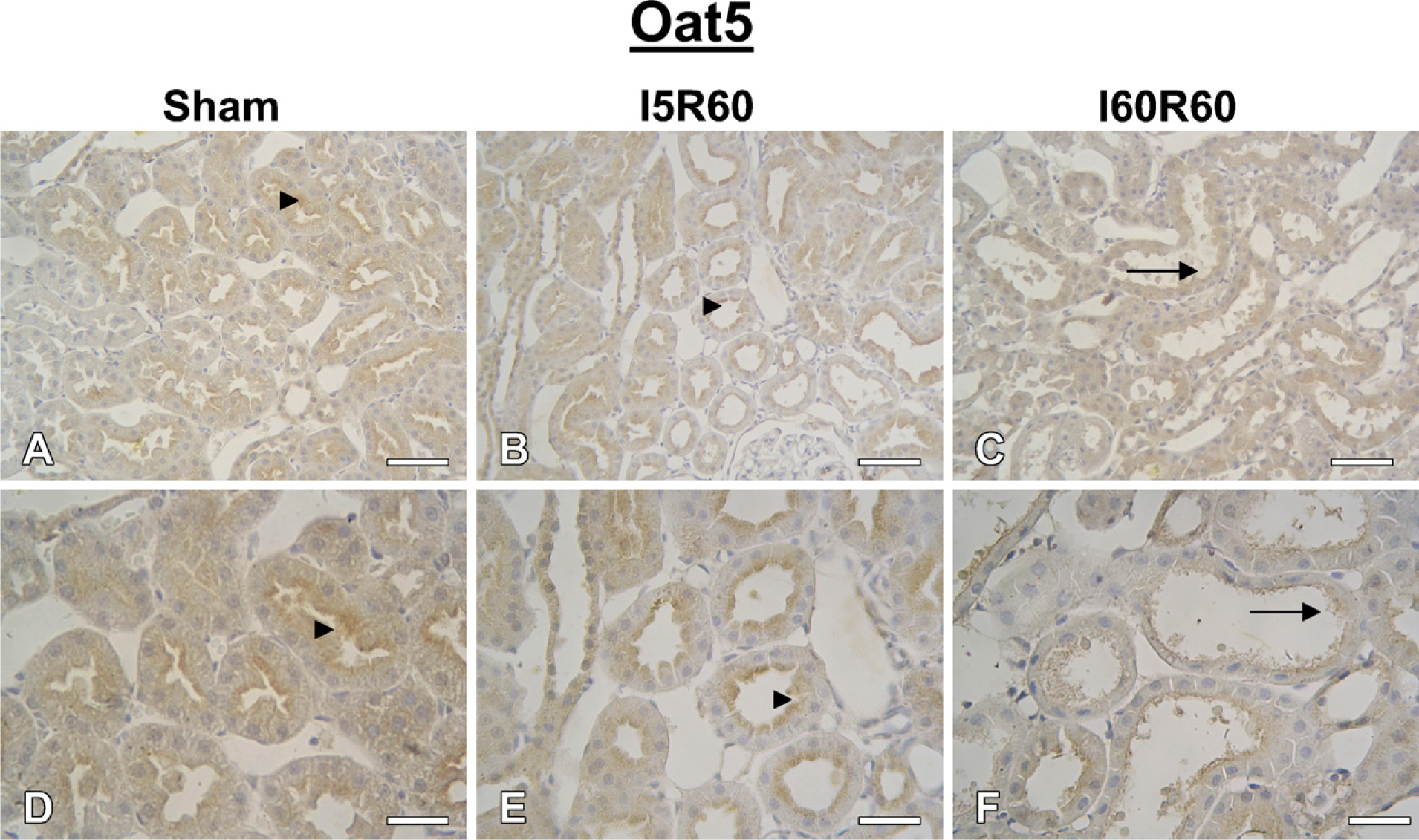
IHC for Oat5 in kidneys from Sham (
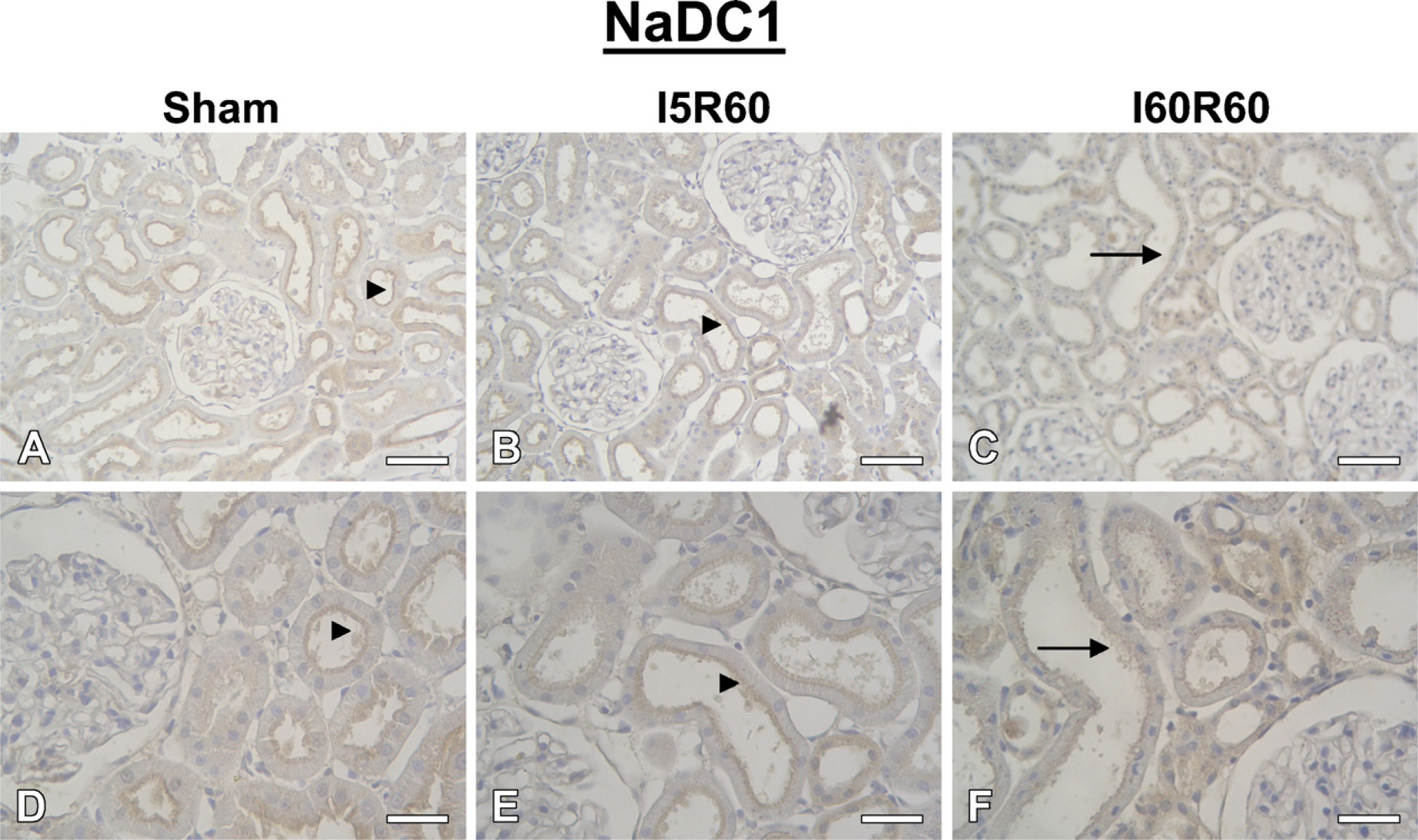
IHC for NaDC1 in kidneys from Sham (
NaDC1 is mainly expressed in the epithelial cells of the kidney proximal tubule and is thought to be involved in the reabsorption of various Krebs cycle intermediates from the tubular filtrate in the kidney. Krebs cycle intermediates are important substrates for renal metabolism because they account for 10-15% of oxidative metabolism in the kidney (Simpson 1983). The proposed function of NaDC1 in the kidney was originally based on its expression in the apical membrane of the proximal tubules and its transport functions examined in intact proximal tubules and membrane vesicles isolated from various species (Sekine et al. 1998; Pajor 2000, 2006). Ho et al. (2007) have recently developed NaDC1 gene knockout mice, which excreted significantly higher amounts of various Krebs cycle intermediates in their urine compared with their wild-type littermates. This result showed for the first time in vivo NaDC1′s role in reabsorbing various types of Krebs cycle intermediates from the tubular filtrate in the kidney.
Oat5 renal expression may have physiological relevance in homeostasis of ES and DHEAS in rodents. In the mammalian blood, both ES and DHEAS represent a pool (reservoir) of inactive steroids, which in various tissues can be enzymatically transformed into various compounds, such as estrone, estradiol, and DHEA, which all exhibit estrogenic actions (Eberling and Koivisto 1994; Ranadive et al. 1998). Because both steroids are reabsorbed in the mammalian kidney (Kellie and Smith 1957; Wright et al. 1978), proximal tubules in the rodents may internalize the filtered ES and DHES (in exchange for intracellular succinate and α-ketoglutarate) because of the expression of Oat5 in the luminal membrane. The internalized compounds may exit the tubules through Oat3 localized in the proximal tubule basolateral membrane (Youngblood and Sweet 2004; Anzai et al. 2006).
The downregulation of Oat5 and NaDC1 protein expression in homogenates and apical membranes from kidneys of I60R60 rats might be caused by the decrease in their synthesis, the increase in their degradation, the liberation of intracellular and apical membrane fractions into tubular fluid, or a consequence of cell shedding. The similar decrease of Oat5 and NaDC1 abundance observed in homogenates compared with apical membranes suggests no impairment in the intracellular distribution of this transporter.
ATP is known to be required for metabolic transformations such as protein synthesis and maintenance of cellular structures. The proximal tubule is largely dependent on oxidative metabolism for generation of ATP. In this regard, it has been reported that ATP levels were at 50% of control values after 40 min of ischemia and 60 min of reperfusion of rat kidneys (Coux et al. 2002). Moreover, it has been recently reported that IRIP (a new ischemia-reperfusion inducible protein) can negatively regulate the activity of a variety of transporters under normal and pathological conditions (Jiang et al. 2005). Therefore, ATP and IRIP levels might downregulate both Oat5 and NaDC1 in I60R60.
Previous studies have shown that Krebs cycle intermediates could confer resistance to renal proximal tubule cell injury induced by hypoxia/reoxygenation. In the normal kidney, proximal tubule cells are especially vulnerable to damage caused by renal ischemia and reperfusion, but some of them are able to survive the injury nevertheless. These surviving cells are thought to be important for renal recovery. Ho et al. (2007) wanted to examine whether Krebs cycle intermediates play a role in protecting these cells from renal ischemia and reperfusion injury. Because they have shown that NaDC1 is responsible for the transport of these intermediates in the kidney in vivo, they hypothesized that renal proximal tubule cells from NaDC1-deficient mice may have a depleted pool of Krebs cycle intermediates compared with wild-type mice, rendering them less resistant to damage and hence less able to aid renal recovery after ischemia-reperfusion injury. However, they showed that absence of NaDC1 had no effect on the severity of renal ischemic damage both morphologically and functionally. Therefore, they concluded that the ability to recover from renal ischemia and reperfusion was not dependent on NaDC1. However, Ho et al. (2007) did not take into account the fact that the expression of NaDC1 might also be decreased after ischemia and reperfusion in mice as we have just shown in rats.
The increased abundance of both Oat5 and NaDC1 in urine from I60R60 rats was expected because tubular cell necrosis is associated with liberation of intracellular and apical membrane fractions into tubular fluid (Edelstein et al. 1997). A similar behavior has been previously reported by Du Cheyron et al. (2003) for the NHE-3 protein, whose abundance is increased in urine from patients with ARF. It is important to note that this study is the first for Oat5 and NaDC1 detection in urine. Oat5 was also increased in urine from I5R60 rats. Because the expression of this protein was not modified in homogenates and in apical membranes in I5R60 rats, it is possible to suggest that, after mild subclinical renal ischemia, Oat5 urinary excretion is dependent of a selective apical pathway.
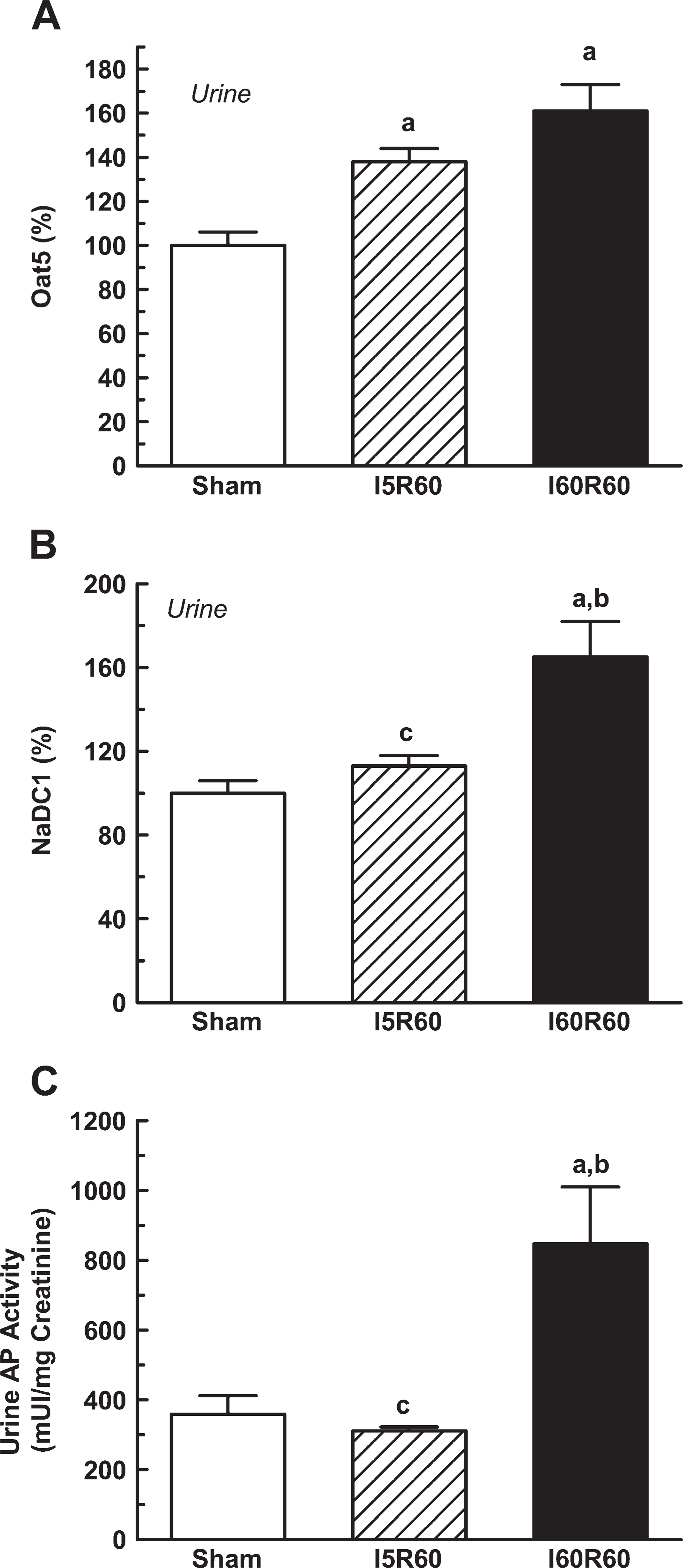
Oat5 abundance (
ARF is common in intensive care units, with a 10-30% incidence that is 5-fold higher than among medical ward patients (Groeneveld et al. 1991; Brivet et al. 1996). The mortality rate for hospitalized patients who develop ARF is approximately five times higher than without ARF (Levy et al. 1996). Despite the introduction of continuous renal replacement therapy and biocompatible membranes, the mortality associated with this diagnostic category has improved little over the last years. The routinely available clinical parameters of kidney disease—plasma creatinine and urea—do not in practice provide either a sensitive or specific indication of renal function, and show ARF well after the injury inducing ARF has occurred. Early detection of acute tubular necrosis could permit implementation of salvage therapies and improve patient outcomes in ARF. Many proximal tubular membrane proteins, enzymes, or usually reabsorbed low-molecular-weight proteins have been used as non-invasive indicators of tubular injury or glomerular filtration rate in specific diseases but have only been suggestive and rarely diagnostic (Muramatsu et al. 2002; Du Cheyron et al. 2003; Zahedi et al. 2003).
The major sodium transporters expressed along the nephron and aquaporin-2 have been detected in urine of rats and humans by means of antipeptide antibodies. The measurements of their renal excretion have begun to be exploited for the study of different pathologies (Wen et al. 1999; Ivarsen et al. 2003). Urinary abundance of the NHE3 has been proposed a novel non-invasive marker of renal tubule damage in patients with ARF (Du Cheyron et al. 2003). In our study, the dramatic urinary increase of Oat5 in the presence of bilateral mild subclinical ischemia and bilateral ischemia with ARF suggests that Oat5 might be a potential early non-invasive marker of this pathology. Plasma creatinine and urinary AP activity are widely used bio-markers for renal dysfunction and injury. These parameters were measured and compared with urinary Oat5 and NaDC1 to evaluate the efficiency of these transport proteins as early diagnostic indicators of kidney injury. In the I60R60 group, all these parameters were increased. In contrast, in the I5R60 group, there was no significant increase in plasma creatinine levels, urinary AP activity, or urinary NaDC1 abundance; on the other hand, there was an increase in urinary Oat5. Thus, the abundance of urinary Oat5 was significantly higher in I5R60 rats, indicating renal injury, although none of the conventionally used bio-markers of renal injury tested in this study were able to predict this.
Several relatively new biomarkers of ARF are being studied. Kim-1 (kidney injury molecule-1) is increased in urine 24 hr after 10 min of bilateral ischemia or 12 hr after 30 min of kidney ischemia in the rat (Vaidya et al. 2006). Cyr61 (cysteine-rich protein 61) was detected after 3-9 hr in urine after bilateral ischemia for 30 min (Muramatsu et al. 2002). NGAL (neutrophil gelatinase-associated lipocalin) was excreted in urine after 6 hr of reflow in mice with 10 min of bilateral ischemia and after 12 hr of reperfusion in rats with 30 min of bilateral renal clamping (Mishra et al. 2003). Therefore, the increase in urinary Oat5 far preceded the increase or the detection in urine of traditional and novel biomarkers of renal disfunction.
On the basis of our results, we suggest that further studies using animal experimental models of ARF and clinical studies of patients at high risk of developing ARF be undertaken to explore the utility of this potential marker. It will be necessary to determine whether it is useful for classification of disease, choice of therapeutic agents, assessment of prognosis, monitoring of a particular therapeutic regimen, and evaluation of nephrotoxicity of pharmaceutical agents in development.
In summary, modifications in renal expression and urinary excretion of Oat5 and NaDC1 were observed 60 min after bilateral ischemia of 60 min, as well as modifications of other widely used parameters for renal dysfunction and injury (plasma creatinine, urinary AP activity, kidney weight, histopathology). In contrast, in the I5R60 group, only an increase in urinary excretion of Oat5 and mild histopathological damage were detected.
These results suggest that urinary excretion of Oat5 might be an early indicator of renal dysfunction that will be useful for detection of even minor alterations in renal structural and functional integrity.
Footnotes
Acknowledgements
This study was supported by the following grants: Fondo para la Investigación Científica y Tecnológica (PICT 05-20201) and CONICET (PIP 5592).
The authors thank Prof. Juan C. Picena (Cátedra de Anatomía y Fisiología Patológicas, Facultad de Ciencias Médicas, UNR) and Alejandra Martínez (Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario) for contributions to the histological studies and Wiener Lab Argentina for the analytical kits.