
Abstract
In pancreatic beta cells, proinsulin (ProIN) undergoes folding in endoplasmic reticulum/Golgi system and is translocated to secretory vesicles for processing into insulin and C-peptide by the proprotein convertases (PC)1/3 and PC2, and carboxypeptidase E. Human beta cells show significant variation in the level of expression of PC1/3, the critical proconvertase involved in proinsulin processing. To ascertain whether this heterogeneity is correlated with the level of expression of the prohormone and mature hormone, the expression of proinsulin, insulin, and PC1/3 in human beta cells was examined. This analysis identified a human beta cell type that expressed proinsulin but lacked PC1/3 (ProIN+PC1/3−). This beta cell type is absent in rodent islets and is abundant in human islets of adults but scarce in islets from postnatal donors. Human islets also contained a beta cell type that expressed both proinsulin and variable levels of PC1/3 (ProIN+PC1/3+) and a less abundant cell type that lacked proinsulin but expressed the convertase (ProIN−PC1/3+). These cell phenotypes were altered by type 2 diabetes. These data suggest that these three cell types represent different stages of a dynamic process with proinsulin folding in ProIN+PC1/3− cells, proinsulin conversion into insulin in ProIN+PC1/3+cells, and replenishment of the proinsulin content in ProIN−PC1/3+ cells:
Introduction
Insulin is translated as preproinsulin and converted into proinsulin (ProIN) in the endoplasmic reticulum (ER)/Golgi system. 1 The maturation of proinsulin into insulin occurs in secretory granules and is mediated by the proprotein convertases (PC) 1/3 and PC2, and carboxypeptidase E (CPE),2 –6 with PC1/3 mediating the initial stage of the maturation process.7,8 The final products of proinsulin processing include insulin and C-peptide. 9 The process of insulin secretion is dependent on Ca2+ and requires the involvement of attachment proteins in a process similar to that in neurons. 10
It has long been known that beta cells are heterogeneous at the functional level.11,12 This heterogeneity includes differences in glucose metabolism, insulin synthesis and secretion 13 and variations in the level of key proteins. 14 The analysis of beta cell heterogeneity has recently received increased scrutiny due to the availability of techniques to probe variability at the transcription and translation level in isolated beta cells. 15 It was recently reported that human beta cells display significant variation in the level of PC1/3 expression. 16 To interrogate the significance of this variation, the coexpression of proinsulin, PC1/3, and insulin (IN) was examined by high-resolution confocal microscopy in islets from non-diabetic and diabetic donors. This analysis revealed the presence of a human beta cell type that expressed proinsulin, had undetectable levels of PC1/3 and very low levels of insulin (ProIN+PC1/3−); a second beta cell type that contained PC1/3 and insulin in addition to the prohormone (ProIN+PC1/3+); and a third cell type that lacked proinsulin but expressed PC1/3 and insulin (ProIN−PC1/3+). Although this analysis gives snapshots of the different human beta cell types, it can be speculated that they represent sequential changes in beta cell phenotypes, with folding of proinsulin in ProIN+PC1/3− cells, processing of proinsulin to maturity in ProIN+PC1/3+ cells, and restocking the cellular content of proinsulin in ProIN+PC1/3+ cells. Interestingly, mouse islets lack ProIN+PC1/3− cells, suggesting that this cell type may have evolved in humans to implement a strict control of proper insulin folding and aggregation.
These three cell types were also found in islets from donors with type 2 diabetes (T2D), along with islets populated by cells displaying partial or total loss in the expression of the hormone precursor and proconvertase. Notably, most ProIN+PC1/3− cells of control and T2D differ in the localization of proinsulin, which is perinuclear in the first and cytoplasmic in the second group, a displacement that may affect the function of ProIN+PC1/3− cells. Taken together, these findings suggest that preservation of the normal cell phenotypes and of their sequential interconversion is linked to normal beta cell function.
Methods
Paraffin sections of human pancreas were provided by Network for Pancreatic Organ Donors with Diabetes (nPOD). 1 To obtain sections of mouse pancreas, 6-month-old C57Bl6J female mice (n=6) were perfused through the heart with a solution of 4% paraformaldehyde in 1× PBS, and the pancreas was removed and processed for cryostat sectioning as described elsewhere. 17 Dr. Janice Brissette (SUNY Downstate Health Sciences University) provided the mice from her colony and euthanized them following animal protocols approved by the Institutional Animal Care and Used Committee of SUNY Downstate Health Sciences University. Animals were monitored until breathing had ceased, confirmed by lack of reflexes. Then, the thoracic cavity of the dead animals was opened to expose the heart and the animals were perfused. No specific protocol was required for perfusion of dead mice.
Immunohistochemistry
Monoclonal antibody to proinsulin was purchased from Novocastra (ProIN-1G4; Buffalo Grove, IL) and from the Developmental Biology Hybridoma Bank (GS-9A8; Nashville, TN). Antibody ProIN-1G4 has been reported to recognize the junction between B-chain and C-peptide.18,19 Manufacturer information indicates no crossreactivity of the antibody with bovine or porcine proinsulin, human C-peptide, or human, bovine, and porcine insulin. Antibody GS-9A8 reacts with an antigenic determinant localized to the site of linkage of the B-chain to the C-peptide (Arg-Arg) at positions 31–32. Antigenic determinants were identified by Madsen et al. 20 and its distribution, assessed by immune electron microscopy, is restricted to the Golgi region. 2 Rabbit antibody to PC1/3, a generous gift from Iris Lindberg (University of Maryland), was generated by D.F. Steiner (University of Chicago). This antibody recognizes the active 87 and 66 kDa forms of the enzyme.21,22 Guinea pig antibodies to insulin (Linco; EMD Millipore, Burlington, MA) colocalized in beta cells with mouse anti-insulin (05-1108; EMD Millipore) (GT, unpublished observation). The mouse monoclonal antibody binds to mature insulin and/or the des-31.32 proinsulin intermediate that has been processed by both PC1/3 and CPE, 19 suggesting similar properties for the Linco guinea pig antibody. It remains to be determined whether the two proinsulin antibodies and the insulin antibody that were used in this study also recognize the proinsulin intermediates produced during processing of the prohormone. Secondary antibodies anti-rabbit Alexa Fluor 488 IgG (green) and goat anti-mouse Alexa Fluor 594 (red) were purchased from Molecular Probes (Eugene, OR); donkey anti-guinea pig-Cy5 IgG (blue) was purchased from Jackson ImmunoResearch (West Grove, PA).
Antibody Concentrations
The concentration of mouse antibody to proinsulin GS-9A8 was 1:500; mouse antibody to proinsulin GS-9A8, 1:250; rabbit antibody to PC1/3, 1:500; guinea pig antibody to insulin, 1:500; and mouse antibody to insulin, 1:500. All secondary antibodies were used at a 1:200 dilution. All antibodies were diluted in 0.1 M Tris saline containing 1% lamb serum.
Fluorescence Microscopy
Images were taken with a Leica SP5 confocal microscope. Confocal Z-stack and single plane images were acquired with an Argon (488 nm), HeNe (543 nm), and HeNe (633 nm) laser source. One-micron-thick Z-stack images were acquired using a 40× Plan-Apochromat (NA1.25) objective. Images were scanned sequentially to prevent crosstalk between the three fluorophores and were scanned at a speed of 400 Hz. Case characteristics are presented in Table 1.
Case Characteristics
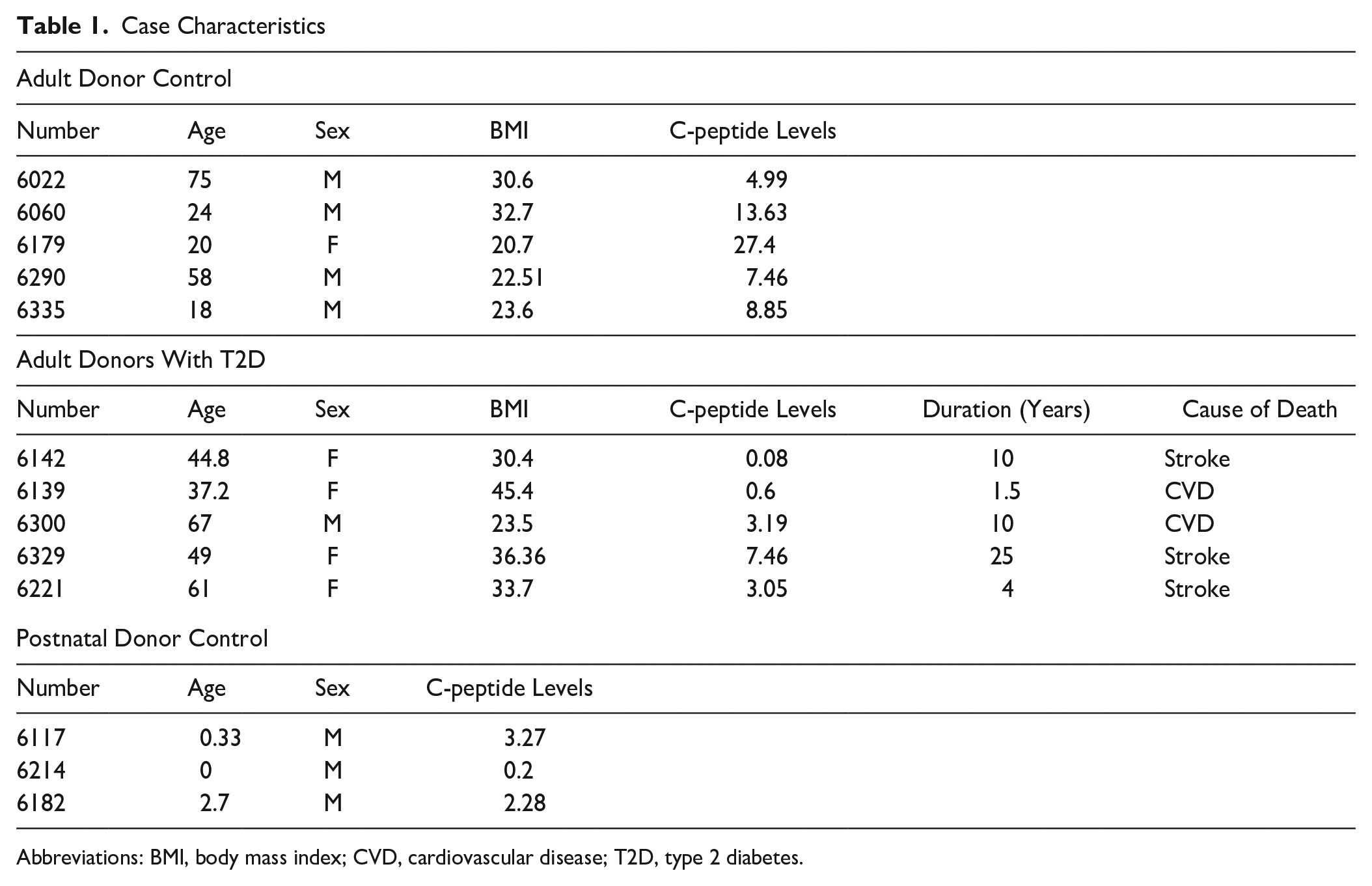
Abbreviations: BMI, body mass index; CVD, cardiovascular disease; T2D, type 2 diabetes.
Image Analysis
The percentage of each cell type in islets was determined using the multipoint tool of Image J (http://rsb.info.nih.gov).
Results
Pancreatic Islets of Adults Contain Three Beta Cell Types
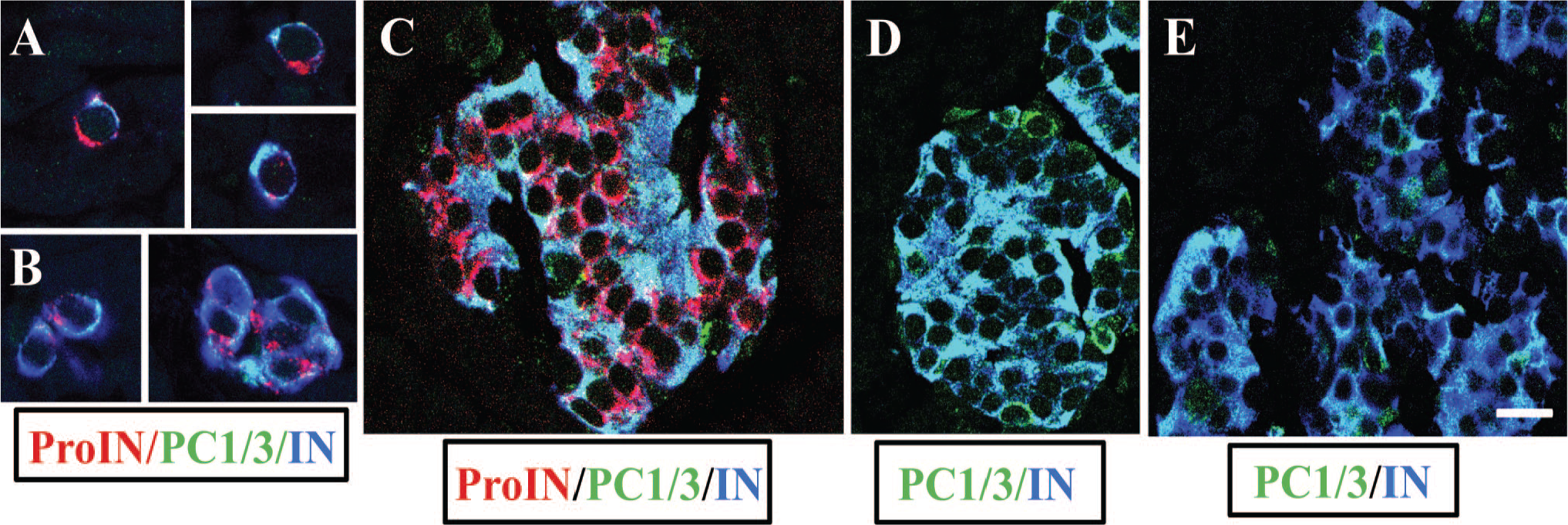
To ascertain whether all insulin cells coexpress proinsulin, PC1/3, and insulin, sections of pancreas from five donor controls were triple-labeled for simultaneous visualization of the three markers and examined by confocal microscopy. Results illustrated in Fig. 1 document the presence of cells that were ProIN+ (Fig. 1A) and had undetectable levels of PC1/3 (Fig. 1B). These cells were also characterized by the expression of very low levels of insulin (Fig. 1C). The overlap of A, B, and C images is shown in Fig. 1D, which establishes the presence of ProIN+PC1/3− beta cells in islets of donor control. The reduced level of insulin expression in ProIN+PC1/3− cells is also shown in Fig. 3A; in this instance, the blue color of the insulin cells (Fig. 3B) was changed to green (Fig. 3C) to facilitate the comparison of insulin staining in IN+ and INlow cells.
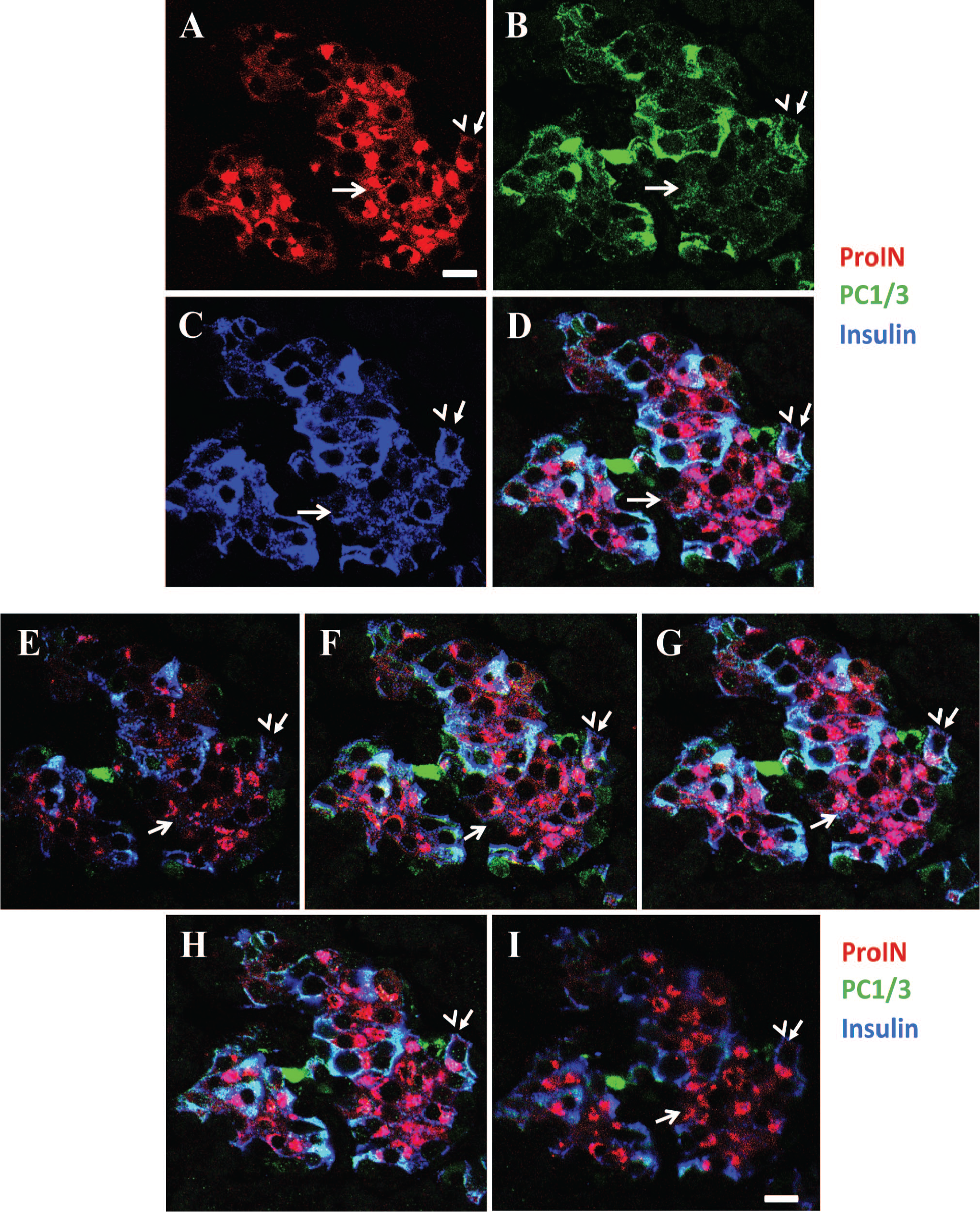
Human islets contain clusters of beta cells that express proinsulin but lack the processing enzyme. (A–D) Photomicrograph illustrates an islet from a donor control immunostained for proinsulin (with antibody ProIN-Ig4; A), PC1/3 (B), and insulin (C). Note that the cluster of ProIN+ cells (indicated with an arrow) has very low levels of PC1/3 and insulin. (D) The overlap of A + B + C. A ProIN+PC1/3+ cell is indicated with an arrow and arrowhead. Bar: 15 µm. (E–H) Z-stack analysis of 1 µm optical sections of the islet illustrated in Fig. 1 A-D. (I) PC1/3- proportion convertase 1/3. Note the persistence of the ProIN+PC1/3− phenotype throughout the 5 μm section. Bar: 20 μm. Abbreviations: PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
The second beta cell type found in human islets is depicted in Fig. 2A to D, which shows an islet in which most cells were ProIN+ (Fig. 2A), PC1/3+ (Fig. 2B), and IN+ (Fig. 2C). Figure 2D illustrates the overlap of the three images of this occasional homotypic islet. Of note, these cells expressed different levels of PC1/3, confirming previous findings in islets of normal donors. 16 A single ProIN+PC1/3+ cell is also shown in Fig. 1A to D. The third beta cell type found in human islets was the ProIN−PC1/3+ cell, which is illustrated in Fig. 3A and D to G. The ProIN+PC1/3− and ProIN+PC1/3+ cell types were found intermingled (Figs.1 and 3A) or in homotypic clusters of 5 to 10 cells (Fig. 2), whereas the ProIN−PC1/3+ beta cells were found isolated or in pairs (Fig. 3G). The presence of these cell types was evaluated by Z-stack analysis of 1 µm optical sections (Fig. 1E–I). This analysis indicated that the pattern of expression of the three cell markers persisted throughout the thickness (5 µm) of the section with no evidence of the presence of a gradient of expression. However, confirmation of these findings will require the development of techniques that will allow the analysis of expression of these markers in whole beta cells while maintaining normal islet structure and the reactivity of the antigens to the antibodies.
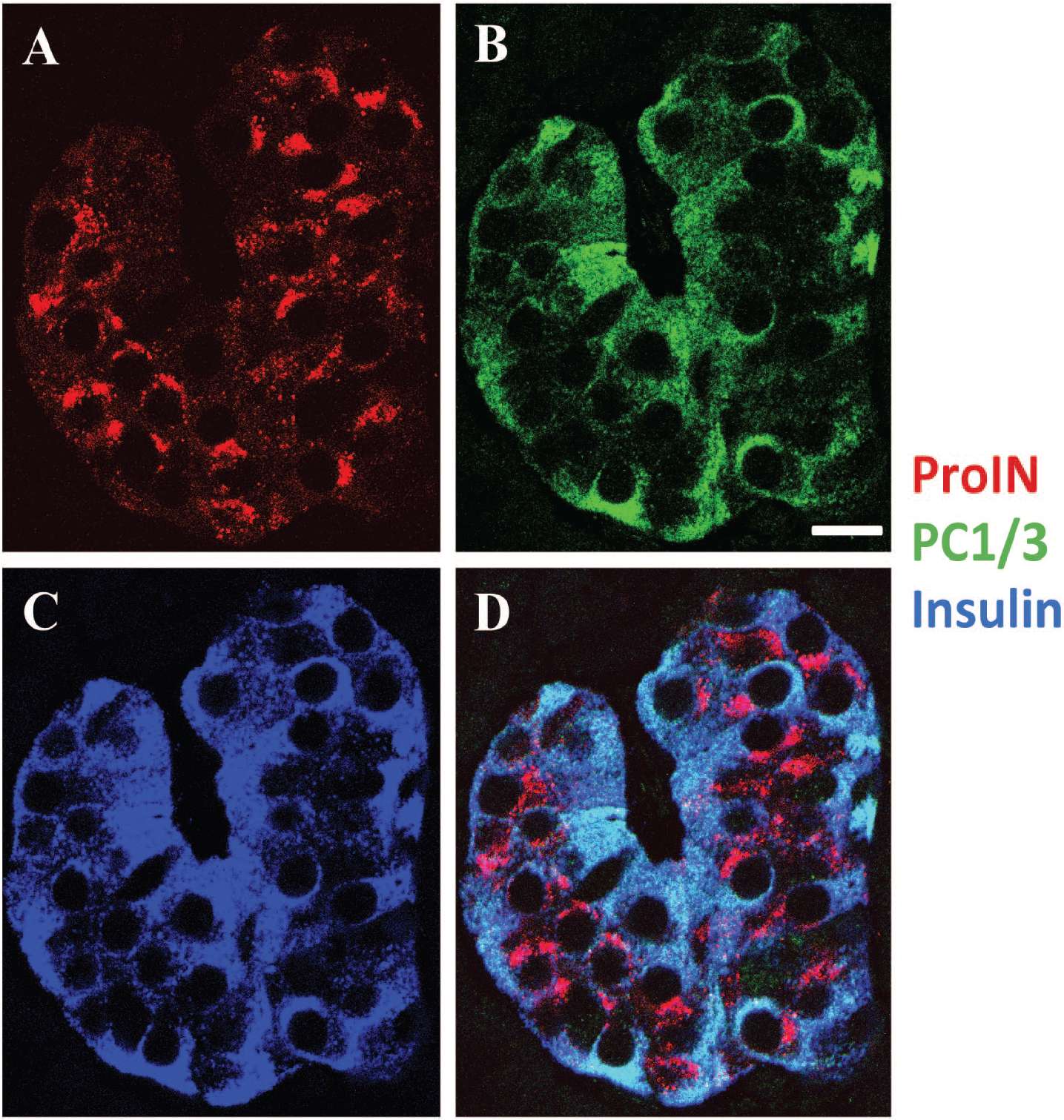
Variation in PC1/3 expression in beta cells of a homotypic human islet. Photomicrographs illustrate an islet comprised of ProIN+PC1/3+ beta cells. (A) ProIN (antibody ProIN-Ig4), (B) PC1/3 and (C) insulin. (D) shows the overlap of A + B + C. Note the difference in PC1/3 level of expression between beta cells. Bar: 15 μm. Abbreviations: PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
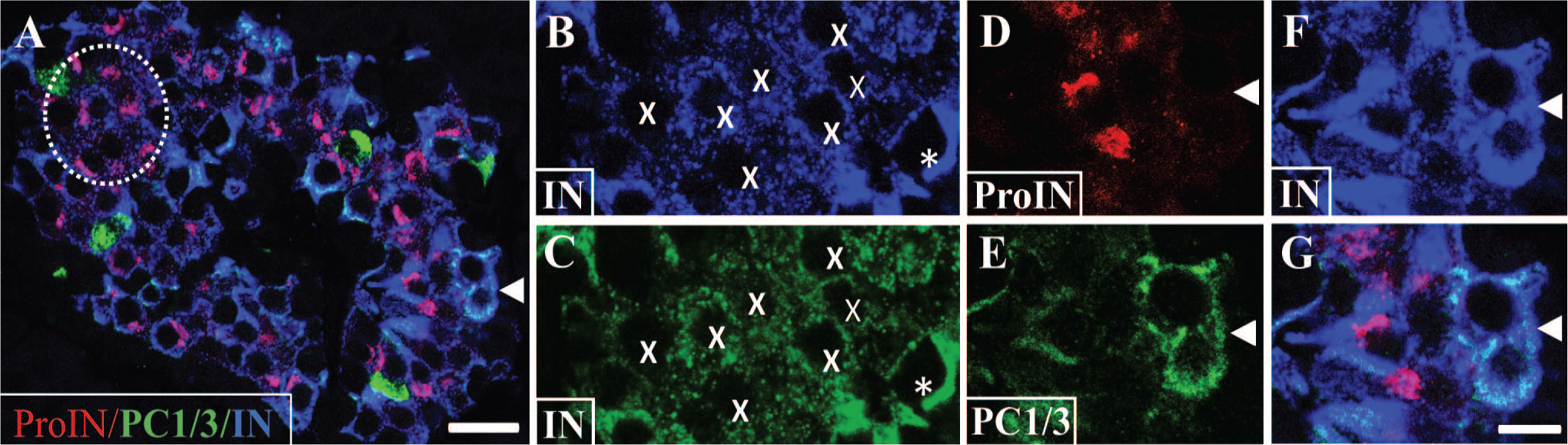
Depiction of beta cell types with or without proinsulin expression. Photomicrograph of an islet of control donor immunostained for proinsulin (antibody GS-9A8; red), PC1/3 (green), and insulin (blue). The dashed circle indicates a cluster of ProIN+PC1/3− cells with low insulin level. Bar: 25 μm. Insulin staining in this cluster is shown magnified in (B; blue). In (C), the blue color was converted to green using Photoshop to facilitate visualization of insulin expression. Note the decreased label in cells marked with an X than in a surrounding cell (asterisk). Cells with only PC1/3 (green fluorescence) probably are somatostatin cells. 16 Bar for (B) to (G): 10 μm. Cells indicated with an arrowhead in (A) are shown magnified in (D) to (G); these cells are ProIN−PC1/3+ because they lack proinsulin (D) but express PC1/3 (E) and mature insulin (F). (G) illustrates the overlap of the three images. Bar: 10 μm. Abbreviations: IN, insulin; PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
The percentage of the three cell types was determined in islets from five donors (Fig. 4). It was found that the percentage of the three beta cell types varied in different islets of the same donor (Fig. 4A) and between the five donors examined (Fig. 4B).
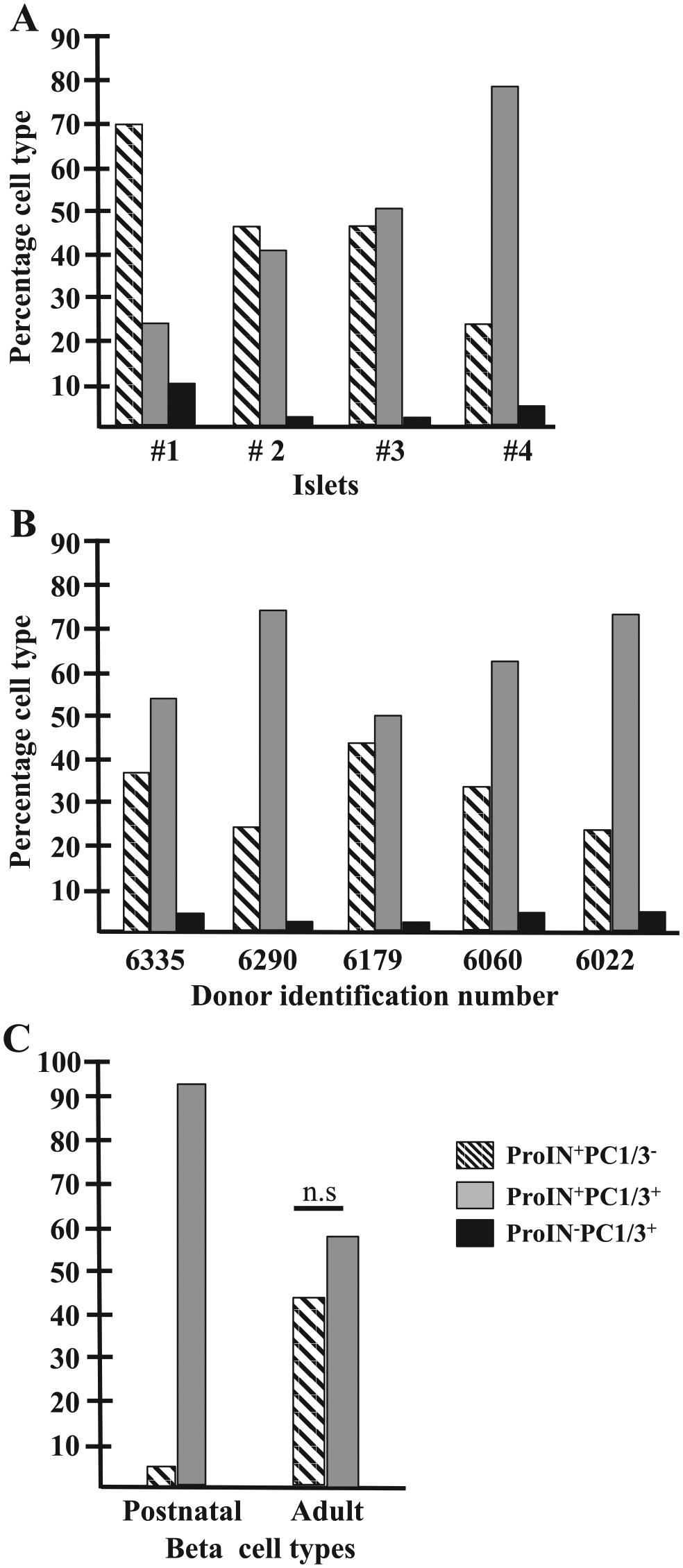
Percentage of the three beta cell types in heterotypic human islets: (A) Histogram illustrates the percentage of beta cell types in four representative islets of one donor control (#6179). This figure does not include the occasional homotypic islet. (B) Histogram indicates the percentage of ProIN+PC1/3−, ProIN+PC1/3+, and ProIN−PC1/3+ cells in islets from five adult donors (15 islets/section, two sections per pancreas; a total of 3312 cells were scored). (C) Histogram compares the percentage of ProIN+PC1/3− (1238 cells) and ProIN+PC1/3+ (1357 cells) in islets of adults and in three young postnatal donors (55 clusters <10 cells and 28 clusters >10 cells, 320 cells scored). Abbreviations: PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
Postnatal Appearance of Beta Cell Diversity
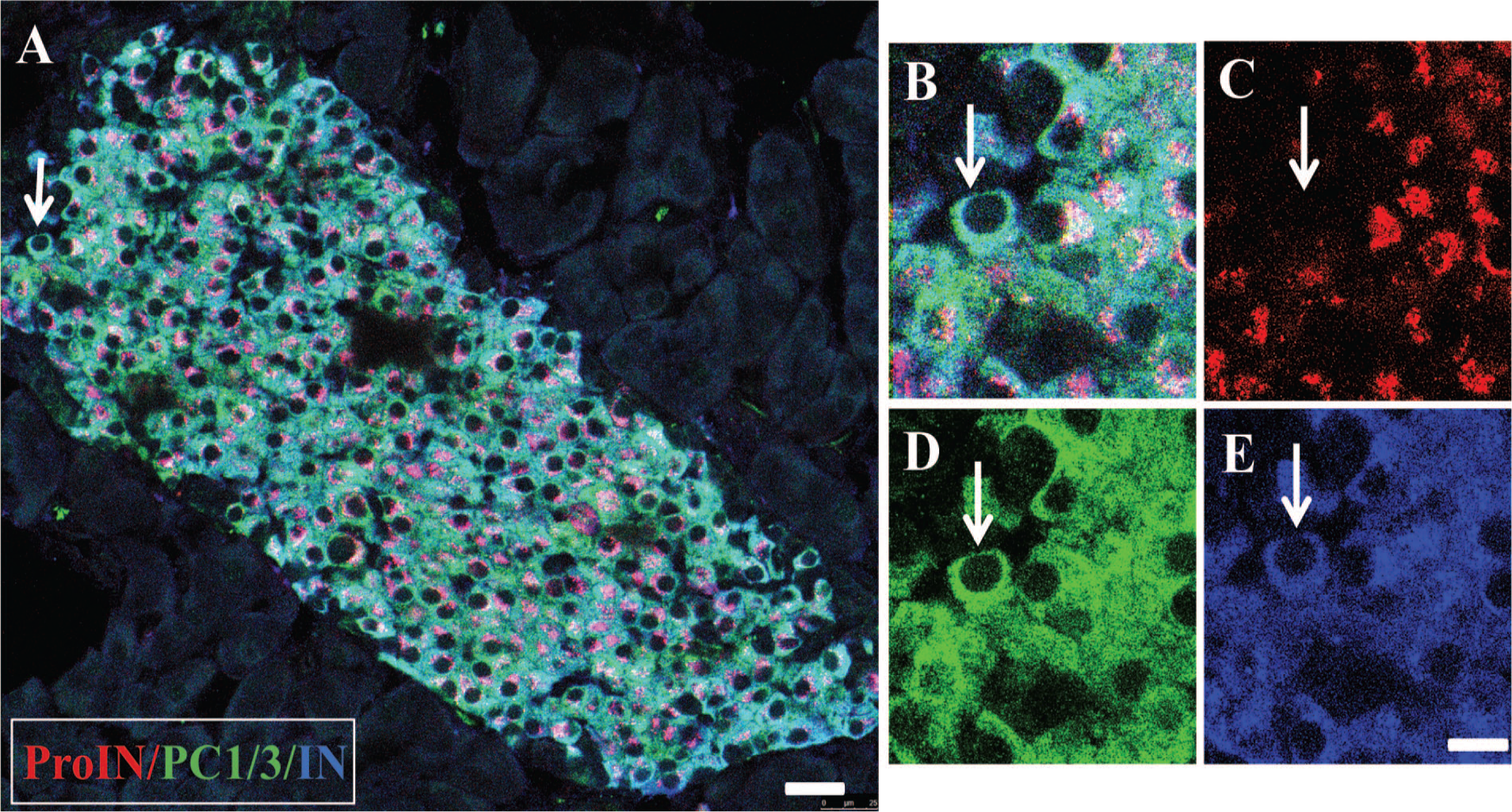
The presence of the three beta cell types in islets of adults raised the question of whether they are generated by a cell autonomous process such that their presence in islets would be independent of extrinsic signals. To assess this possibility, pancreas of newborn and early postnatal donors was examined. As it has been reported that islets are formed during development by clustering of isolated cells, 23 it was hypothesized that the phenotype of isolated cells present in pancreas of non-diabetic donors would be similar to that of beta cells in clusters if the cells are not affected by cell–cell contact. All isolated beta cells examined (256 cells from three donors, Table 1) were ProIN+PC1/3+ and had only a small rim of cytoplasm (Fig. 5A) which became enlarged following cell clustering (Fig. 5B and C). Larger islets were composed mostly of ProIN+PC1/3+ cells with scattered ProIN+PC1/3− or ProIN−PC1/3+ cells. This finding indicates that clustering affects cell size.
Beta cell clustering and maturation. (A) illustrates three isolated beta cells, whereas (B) shows two small clusters in pancreas of newborn stained for proinsulin (antibody ProIN-Ig4, red), PC1/3 (green), and insulin (blue). Note that these cells contain only a narrow rim of cytoplasm that increases in size in larger cell aggregates (C). Photomicrographs illustrate a pancreatic islet of postnatal donors (D) and adults (E) immunostained for PC1/3 (green) and insulin (blue). Note that the expression of the convertase was similar in all beta cells of the young donors but varied significantly in those of adults. Bar: 20 μm. Abbreviations: IN, insulin; PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
Comparison of the percentage of ProIN+PC1/3− cells in islets of young and adult donors revealed significant differences between the two groups. Thus, although in adults the percentage of ProIN+PC1/3− and ProIN+PC1/3+ cells differed between islets (Fig. 4A), these two cell types were amply represented in a large islet population (Fig. 4C). In contrast, islets from young donors were mostly composed of ProIN+PC1/3+ cells (Fig. 4C). Comparison of PC1/3 expression in beta cells of young postnatal and adult donors revealed an additional difference between the two groups. Thus, the level of expression of the convertase was similar for all beta cells of islets from early postnatal donors (Fig. 5D), whereas the enzyme displayed significant heterogeneity in beta cells of islets from adult donors 16 (Fig. 5E). These observations suggest that the abundance of the ProIN+PC1/3− cell type in islets and the appearance of variations in PC1/3 expression in ProIN+PC1/3+ cells are correlated with time of life.
Islets of Adult Mice Contain Two Beta Cells Types
In contrast to human islets of adults, most beta cells of islets of 6-month-old mice expressed similar PC1/3 levels and displayed a ProIN+PC1/3+ phenotype (Fig. 6A). Islets also contained rare ProIN−PC1/3+ beta cells (Fig. 6B–E) but lacked ProIN+PC1/3− cells. Of note, proinsulin expression was lower with both proinsulin antibodies in mouse than in human islets, perhaps reflecting decreased level of the precursor compared with humans or of its affinity to the antibody. The pattern of expression of the markers was confirmed by 1 µm Z-stack analysis, which also indicated that the antibodies penetrated throughout the 20-µm tissue slice (not shown).
Islets of adult mice contain only two beta cell types: (A) Photomicrograph illustrates an islet immunolabeled for proinsulin (antibody GS-9A8; red), PC1/3 (green), and insulin (blue). Note that all beta cells express similar levels of the convertase. Identical results were obtained using proinsulin antibody ProIN-Ig4 (not shown). Bar: 25 µm. (B)–(E) Cell indicated with an arrow in (A) is a ProIN−PC1/3+ cell (B) because it does not express proinsulin (C) but stains for PC1/3 (D) and insulin (E). Bar: 8 μm. Abbreviations: IN, insulin; PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
Expression of PC1/3 and ProIN in Islets of Donors With T2D
To determine the effect of T2D in the expression of three beta cell markers, 50 islets from five diabetic donors were examined. Each pancreatic slice contained islets comprising beta cells showing decreased PC1/3 and/or displaced proinsulin expression (Fig. 7A). Also present were islets containing ProIN+PC1/3+cells (Fig. 7C) and ProIN−PC1/3+ cells (Fig. 7D–G) exhibiting normal localization of the markers. In contrast to controls, only one cluster of ProIN+PC1/3− with perinuclear proinsulin label was found in all the islets/clusters from T2D donors examined (Fig. 7B). More common were ProIN−PC1/3+ cells with abundant proinsulin expression in the cytoplasm (Fig. 7G and H). All these cell types were still positive for insulin (Fig. 7A and F), presumably reflecting the presence of defects in insulin secretion associated with the disease.
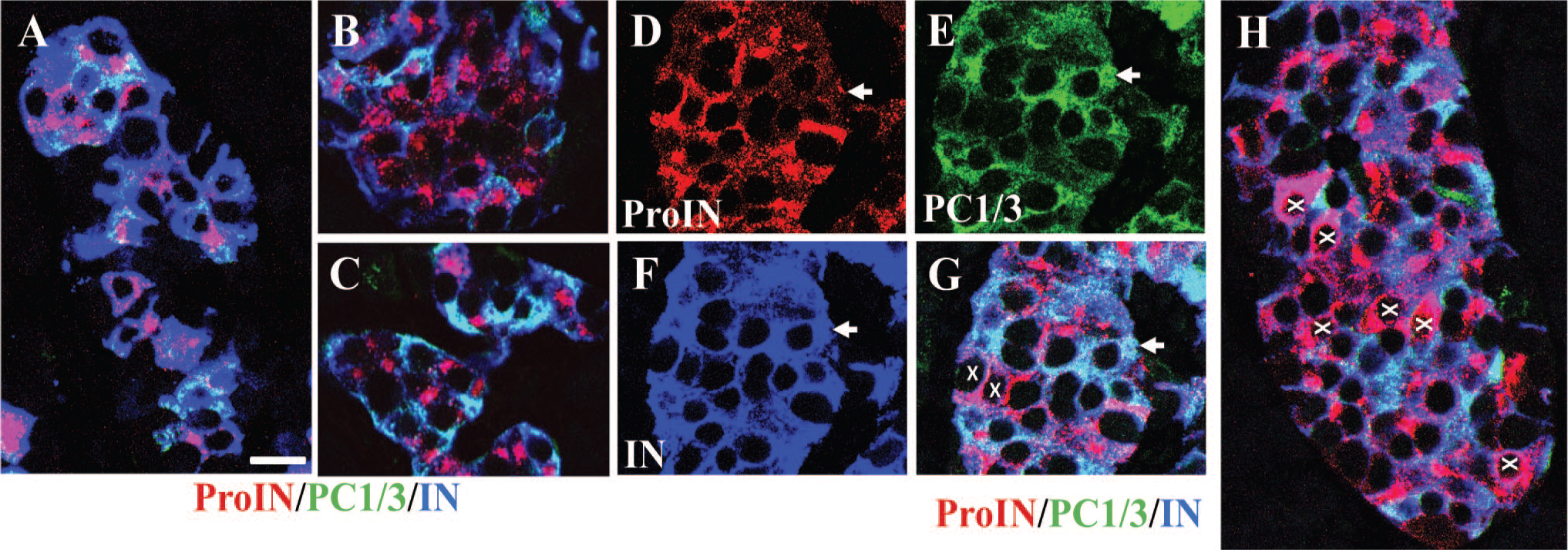
Some islets of diabetic donors contain beta cells with abnormal phenotypes. Figures show representative islets in pancreas of diabetic donors immunostained for ProIN (antibody ProIN-Ig4; red), PC1/3 (green), and insulin (blue). (A) illustrates an islet with decreased and/or anomalous expression of proinsulin and PC1/3, (B) shows a cluster of ProIN+PC1/3− cells, and (C) shows a cluster of ProIN+PC1/3+ cells. An islet containing one ProIN−PC1/3+ beta cell (arrow) is shown stained for ProIN (D), PC1/3 (E), and insulin (F); (G) shows the overlap of the three markers. Cells indicated with an x show cytoplasmic localization of insulin. (H) illustrates an islet containing several ProIN−PC1/3+ cells (indicated with X) with cytoplasmic localization of the prohormone. Bar: 15 μm. Abbreviations: IN, insulin; PC1/3, proprotein convertase 1/3; ProIN, proinsulin.
Discussion
The major stages involved in the processing of proinsulin into mature insulin and C-peptide were elucidated by the elegant work of Orci and collaborators. 24 The discovery that enzymes included in the secretory vesicles, the PCs, are key participants in the process of maturation provided further insight into the sequence of molecular events involved in the cleavage of the hormone precursor. 4 However, it remained to be determined whether beta cells differ in the level of expression of key molecules involved in the production of insulin. A specific goal of the present studies was to determine whether variations in the level of PC1/3 among human beta cells 16 were correlated with changes in the expression of proinsulin and insulin. This analysis led to the discovery of three human beta cell types that differed in the expression of these markers. One of these cells had decreased levels of mature insulin and undetectable levels of PC1/3 (ProIN+PC1/3−), which is the main enzyme involved in proinsulin cleavage.4,7,25
The transient nadir in PC1/3 expression is intriguing, particularly because the identity of the signals regulating its level in human beta cells is unknown. It is possible that the difference in PC1/3 expression reflects variations in the level of PC1/3 transcripts. Indeed, recent single cell transcriptome analysis of human beta cells revealed significant variation in PCSK1 levels. 26 It is also possible that the regulation occurs at the level of translation, with metabolites affecting its rate 27 and structure. Alternatively, the regulation of PC1/3 expression could occur at the level of the mature enzyme. PC1/3, like proinsulin, is produced as an inactive precursor that undergoes an initial autocatalytic pro-peptide cleavage in the ER and a second cleavage that is promoted by the acidic environment of the immature secretory granules, 28 transforming into the mature enzyme. PC1/3 activity is known to be regulated by oligomerization and binding to specific endogenous proteins.29 –31 It has also been reported that PC1/3 can be present in states with different kinetics properties and that binding of the substrate to its inactive form slowly draws the population into a catalytically active form. 32 This response, termed hysteretic, is characteristic of enzymes involved in metabolic pathways. 33 Taken together, these considerations indicate that the analysis of the regulation of PC1/3 expression in human beta cells is likely to provide important information of the mechanisms affecting the activity of this important metabolic enzyme.
The appearance of PC1/3 expression is correlated with the increase in mature insulin that is characteristic of ProIN+PC1/3+ cells. The level of PC1/3 in ProIN+PC1/3+ cells is variable, suggesting that the cells differ in the concentration of the molecular form of the enzyme that is recognized by the antibody.22,34 The observations also suggest that the ProIN+PC1/3+ cells become the ProIN−PC1/3+ cell type following the conversion of the cellular content of proinsulin into insulin. It can also be speculated that ProIN−PC1/3+ cells reinitiate expression of proinsulin and lose PC1/3 expression, a progression that leads to the re-emergence of the ProIN+PC1/3− cell type with the hormone precursor in a perinuclear location. This hypothetical model of phenotypic interconvention is illustrated in Fig. 8. This model, generated from the analysis of a small number of cases and a single analytical approach, will provide a blueprint for future studies geared to the development of techniques that will allow the unbiased examination of a larger sample number. In addition, it will be of particular interest to ascertain whether one of the three cell types described here shows immature traits or displays pacemaker properties similar to those found in mouse beta cells.35 –37
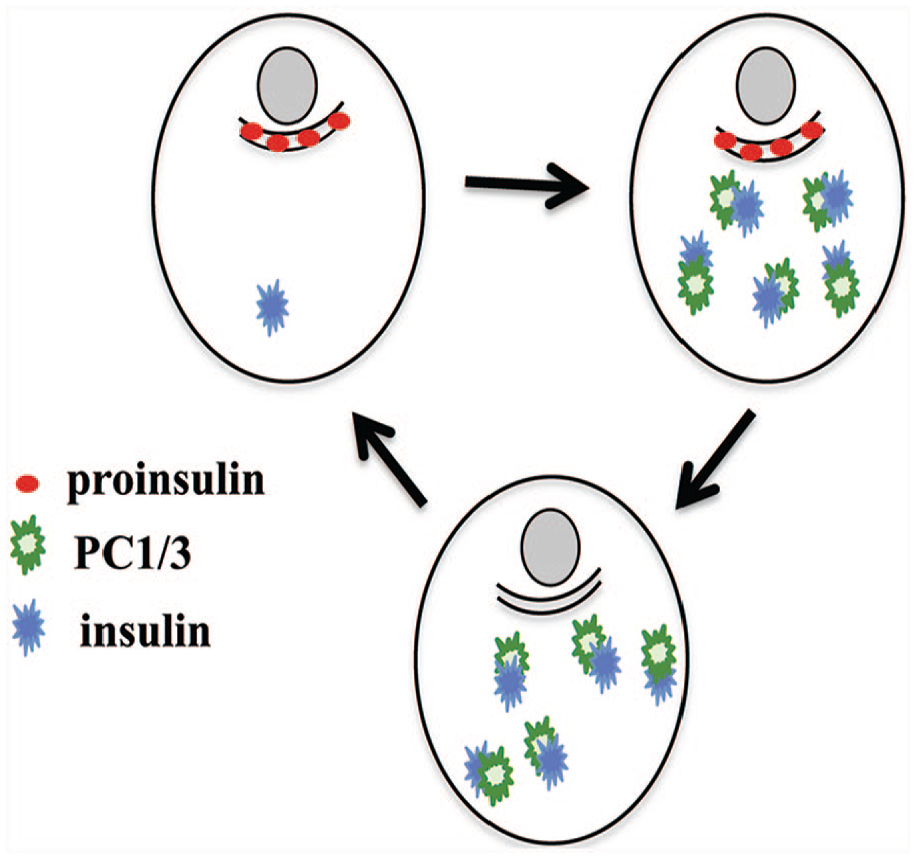
Hypothetical model of interconversion of beta cell phenotypes in human islets. It is postulated that human islets contain three beta cell types, namely, ProIN+PC1/3+, ProIN−PC1/3+, and ProIN+PC1/3− cells, and that these cells convert into the next cell type in the sequence. Abbreviations: PC, proprotein convertase; ProIN, proinsulin
This study also revealed that the percentage of ProIN+PC1/3− cells increased during postnatal life. Thus, evaluation of the percentage of three beta cell types in pancreatic islets from donors younger than 3 years old indicated that most beta cells expressed the ProIN+PC1/3+ phenotype. In contrast, adult islets contained similar percentage of ProIN+PC1/3− and ProIN+PC1/3+ cells. It is known that human islets undergo profound maturation changes during the postnatal period that include the ability to secrete insulin in response to glucose 38 and that changes in diet have a significant effect on the functional and structural maturation of the islets.39 –43 The increased number of ProIN+PC1/3− cells is likely to reflect the functional maturation of the islet population in humans, which includes the known cellular effort involved in the correct processing of the precursor in beta cells. 44
Analysis of pancreas from donors with T2D revealed the presence of islets comprising beta cells displaying normal localization of the three markers examined. However, clusters of ProIN+PC1/3− cells with perinuclear localization of the prohormone were extremely rare. More common were beta cells showing abundant proinsulin labeling in the cytoplasm, a characteristic of abnormal processing of the precursor. 18 These observations suggest that the ProIN+PC1/3− cells found in T2D islets are derived from the homonymous cells that populate normal islets and that this stage is followed by the decline in prohormone and enzyme expression during progression of the metabolic injury to the cells. This possibility will support the hypothesis, raised by others,44,45 that defects in proinsulin processing play a key role in the development of diabetes. However, these considerations underscore the fact that the discovery of different beta cell phenotypes in normal islets and of their alteration in islets from diabetic donors resulted from an approach that does not allow to follow the fate of individual cells in time. Further analysis will require use of a combination of technical tools to characterize the molecular properties of each cell type and establish their relationships.
A recent study discovered that human beta cells display heterogeneity in the level of expression of the insulin gene with cells containing either high or low level of expression of the gene. That study postulated that, following active insulin synthesis, beta cells activate a stress response geared to maximize proper folding of the molecule. 46 Conceivably, the ProIN−PC1/3+ cells are cells actively transcribing the insulin gene, whereas the cells involved in processing the precursor and characterized by high levels of stress in the ER would be the ProIN+PC1/3− cells. Analysis of the level of expression of the insulin gene in mouse islets indicated that most beta cells had low level of expression of the gene, whereas a small proportion expressed high levels of insulin mRNA, proinsulin, and PC2. 47 Perhaps the ProIN+PC1/3− cells are the human equivalent of those high expressers reported by Farack et al. 47 in mice. However, the present study indicates that mouse beta cells expressed similar levels of PC1/3, which is the preponderant enzyme involved in proinsulin processing.8,9 Further studies will seek to elucidate the cause of this discrepancy.
Although the distribution of the three beta cell types in human islets was varied, it was common to find clusters containing only one cell type. These clusters either were isolated or were part of a large islet that contained cells expressing a different phenotype. It has been reported that small human islets are formed by modules comprising a center core of insulin cells surrounded by a mantle of non-beta cells, 23 whereas many large islets contained several of these well-defined clusters. This study suggests that each initial cluster comprises either ProIN+PC1/3− or ProIN+PC1/3+ cells with an occasional ProIN−PC1/3− cell, and that these clusters aggregate forming larger islets and eventually mingle as a result of cellular rearrangements.
The presence of different beta cell types, the proposition that these cells interconvert, and the increased representation of one of these stages in human islets of adults suggest that these conversions play a critical role in the regulation of insulin production. It will be important to confirm the presence of the three cell types, identify the signals regulating the transitions between them, and elucidate whether the alteration of this sequence plays a role in the development of T2D.
Footnotes
Acknowledgements
A special thanks to the families of the donors who provided tissues for this study.
Competing Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
GT designed the study, performed the experiments, and wrote the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: These studies were partially supported by NIH grant 1S10RR026732-01 and intramural funds. Research was performed with the support of the Network for Pancreatic Organ donors with Diabetes (nPOD; RRID:SCR_014641), a collaborative type 1 diabetes research project sponsored by JDRF (nPOD: 5-SRA-2018-557-Q-R), and The Leona M. & Harry B. Helmsley Charitable Trust (Grant#2018PG-T1D053). The content and views expressed are the responsibility of the authors and do not necessarily reflect the official view of nPOD. Organ Procurement Organizations (OPO) partnering with nPOD to provide research resources are listed at ![]() .
.